
G.M. Sindhu
Department of Studies in Botany, Applied Plant Pathology Laboratory, University of Mysore, Manasagangotri, 570 006, Mysuru,
Karnataka, India
M. Murali
Department of Studies in Botany, Applied Plant Pathology Laboratory, University of Mysore, Manasagangotri, 570 006, Mysuru,
Karnataka, India
M.C. Thriveni
Department of Studies in Botany, Applied Plant Pathology Laboratory, University of Mysore, Manasagangotri, 570 006, Mysuru,
Karnataka, India
N. Anupama
Department of Studies in Botany, Applied Plant Pathology Laboratory, University of Mysore, Manasagangotri, 570 006, Mysuru,
Karnataka, India
K.N. Amruthesh
Department of Studies in Botany, Applied Plant Pathology Laboratory, University of Mysore, Manasagangotri, 570 006, Mysuru,
Karnataka, India
LiveDNA: 91.24229
Asian Journal of Crop Science
Year: 2018 | Volume: 10 | Issue: 4 | Page No.: 160-167







ABSTRACT
Background and Objective: Melon cultivation is more depended on synthetic fungicides to control gummy stem blight caused by Stagonosporopsis cucurbitacearum (KJ782214). This study mainly aimed to minimize the use of harmful chemicals to control disease by inventing the biological alternatives. Materials and Methods: The crude proteins (CP) were extracted from rhizospheric fungi Penicillium verruculosum (Talaromyces verruculosus) (KU645999) to understand the growth promoting and disease protection ability of CP at four concentrations against gummy stem blight pathogen. Results: The CP treatment at 100 μg mL–1 for 6 h showed enhance vegetative growth parameters like germination (90%), seedling vigor (2329.11), plant height (42.35 cm), number of leaves (5) and root and shoot weights (0.05-2.3 g) and chlorophyll content (4.16 mg g–1) when compared to control. The CP treatment at 100 μg mL–1 for 6 h also exhibits considerable disease protection of 74.37%, which was found to be higher compared to control and almost equal to the effectiveness of fungicide (Mancozeb) treated (94.03%) plants. Conclusion: Present study concludes with the invention of protein composition of the rhizospheric fungi from muskmelon field as potential alternative to harmful chemicals in controlling the disease.
PDF Abstract XML References Citation
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
G.M. Sindhu, M. Murali, M.C. Thriveni, N. Anupama and K.N. Amruthesh, 2018. Growth Promotion and Disease Resistance in Muskmelon Induced by Crude Proteins of Penicillium verruculosum Against Gummy Stem Blight Disease. Asian Journal of Crop Science, 10: 160-167.
DOI: 10.3923/ajcs.2018.160.167
URL: https://scialert.net/abstract/?doi=ajcs.2018.160.167
DOI: 10.3923/ajcs.2018.160.167
URL: https://scialert.net/abstract/?doi=ajcs.2018.160.167
INTRODUCTION
Plants protect themselves against broad range of pathogens by developing complex immune system1,2. The immune system is triggered by the mediators, such chemicals are known as “elicitors”. The term elicitor was used for molecules capable of inducing the production of active compounds. They do not have any common chemical structure. These chemicals belong to a wide range of different classes of compounds viz., oligosaccharides, peptides, proteins, glycoproteins and sphingolipids3. Many elicitors have been isolated from various organisms, viz., bacteria, viruses and fungi to improve the pathogen resistance of plants. With the discovery of protein elicitors, there has been great interest in manipulating the inducible responses of plants for crop protection. Searching for novel protein elicitors has become a popular strategy in plant disease control4,1. A bio-preparation has been used successfully in many plants to elicit plant disease and insect resistance, to promote growth, yield and quality5. Many protein elicitors have been isolated and used to improve the pathogen resistance of plants. For example, protein elicitor harpins were first isolated from Erwinia amylovora6. They are glycine-rich proteins that lack cysteine, show heat-stable HR elicitor activity when infiltrated into the leaves of tobacco and several other plants7-8. It was shown to elicit the hypersensitive reaction (HR) and induce disease resistance in many plants6,9-12.
Gummy stem blight (GSB) disease is caused by Stagonosporopsis cucurbitacearum (Fr.:Fr.) Aveskamp, Gruyter and Verkley, which leads to severe yield losses in melons (Cucumis melo L.), watermelons (Citrullus vulgaris Schrad.), cucumbers (Cucumis sativus L.) and other Cucurbitaceae crops13. The GSB was observed and identified out in the early 1980s in Europe14,15, in the United States16-18, China, Japan and other tropical and subtropical countries19,20. The fungus causes seedling damping-off, foliar lesions as well as, stalk and stems cankers. It can be universally found in every continent and attacks at least 12 genera and 23 species of cucurbit plants21. Farmers depend mainly on synthetic fungicides to control cucurbit diseases. However, in addition to environmental pollution and contamination of consumable products, several cases of resistance to the main active fungicides on the market have been reported for GSB22,23. There is a need for alternative eco-friendly biological control measures. Hence, the aim of the present study was to investigate the effect of crude proteins from rhizospheric fungi on the plant growth and induction of disease protection in muskmelon against gummy stem blight disease. For this purpose, proteins were extracted from the rhizospheric fungi Penicillium verruculosum (the revised name: Talaromyces verruculosus) which, gave very good results of growth promotion and disease protection when primed to the seeds treated with conidial suspension in our previous studies.
MATERIALS AND METHODS
Collection of muskmelon infected plants and seed samples: Field survey was conducted in muskmelon growing agroclimatic zones of Karnataka for the collection of infected plant samples. Muskmelon plants were diagnosed on the basis of typical symptoms of GSB like, stem necrosis with gummy exudate, angular water-soaked lesions on the leaves. Seed samples were collected from the farmers, agricultural farms, Public and Private Seed Agencies, brought to the laboratory and stored at 4°C for further use.
Screening and isolation of S. cucurbitacearum: Seeds from infected fruits and symptomatic plant parts collected from fields of muskmelon were surface sterilized with 0.2% sodium hypochlorite (NaOCl) for 5 min and repeatedly washed with Sterile Distilled Water (SDW) to remove traces of sterilant. These samples were screened for pathogen following Standard Blotter Method (SBM) and incubated for 7 days at 25±2°C 24. After the incubation period, each sample was examined under Stereomicroscope for fungal colonies showing typical sporulating structure of S. cucurbitacearum. These colonies were aseptically picked with sterile needle and transferred on to Potato Dextrose Agar (PDA) medium. Pathogen was identified based on morphological, conidial, cultural characteristics25, sequence analysis of ITS region and comparison analysis with GenBank data. Sequence was deposited in GenBank and obtained the accession number (KJ782214).
Isolation and identification of rhizospheric fungi: One gram of soil sample was suspended in 9 mL of SDW. An aliquot of 0.1 mL of 10–3 to 10–5 dilutions from each of the soil samples were prepared. About 1 mL of each soil dilution was inoculated on PDA plates supplemented with antibiotic (100 mg L–1) under aseptic condition and incubated for 7 days at 25±2°C. The fungus was identified as Penicillium verruculosum (MRS-PGPF 24) on the basis of microscopic (mycelia and conidia) and macroscopic (culture morphology and appearance) characteristics25, sequence analysis of ITS region and comparison analysis with GenBank data. Sequence was deposited in GenBank and obtained the accession number (KU645999).
Preparation of pathogen (S. cucurbitacearum) inoculum: Spore suspension of S. cucurbitacearum was prepared using SDW by harvesting spores by lightly scraping the surface of the sporulating PDA culture plates with a sterilized spatula under aseptic conditions. The concentration of the inoculum was adjusted to 1×105 spores/mL using Haemocytometer26 and used for further studies.
Mass production of Penicillium verruculosum (MRS-PGPF 24) for elicitor extraction: Fungi that stood best for growth promotion and disease protection was subjected for extraction of crude elicitors (Proteins). Fungal colonies were picked from the actively growing margin of the PDA plates (5 mm) with a sterile fine-tipped needle and transferred on to 500 mL Erlenmeyer flask containing 200 mL PDB. The inoculated PDB was incubated at 25±2°C under 12/12 h alternate cycles of light and darkness for 15 days. Fungal mat was separated from crude culture filtrate by pouring through two layers of cheese cloth. This was further filtered through two layers of Whatman No.2 filter paper27.
Extraction of cell wall crude proteins (CP) from Penicillium verruculosum (MRS-PGPF 24): Crude protein extract was isolated by the modified method of Djonovic et al.28. The fungal liquid culture was strained through two layers of cheese cloth. Each of the fungal supernatant was subjected to (NH)4SO4 precipitation for 5 h at 4°C with continuous stirring while monitoring the pH (pH 7). The pellet obtained at 80% saturation was collected by centrifugation at 12,000 g for 15 min at 4°C and dissolved in minimal volume of column buffer (50 mM sodium phosphate, 0.15 M NaCl, pH 7.0). Samples were desalted by applying them on to desalting column (Genei) to remove the ammonium sulphate. The amount of protein was estimated using a Bradford protein assay with bovine serum albumin (BSA) as standard. The CP powder was diluted with SDW to prepare various concentrations (50, 100, 150 and 200 μg mL–1) and used for seed treatment.
Muskmelon seed treatment with crude proteins (CP) from Penicillium verruculosum (MRS-PGPF 24): Seeds of muskmelon were surface sterilized with 0.2% NaOCl for 5 min, followed by repeated washing with SDW. After surface sterilization the seed samples were treated with CP at 50, 100, 150 and 200 μg mL–1 for 3 and 6 h, respectively. After the incubation period treated seeds were air dried aseptically and further used to study their efficacy of muskmelon plant growth promotion as well as protection against gummy stem blight disease. Distilled water treated seeds and fungicide (Mancozeb) treated seeds served as negative and positive control, respectively.
Effect of seed treatment with Crude Proteins (CP) from Penicillium verruculosum (MRS-PGPF 24) on seed germination and seedling vigor of muskmelon under in vitro conditions: The seed treatments with CP and SDW were used to study the growth of muskmelon.
The seeds were evaluated for germination (%)29 and seedling vigor30. The experiment consisted of four replicates of 100 seeds each and was repeated thrice. After 7 days of incubation at 25±2°C germination (%) and vigor index was calculated as mentioned below:
 |
Effect of seed treatment with Crude Proteins (CP) from Penicillium verruculosum (MRS-PGPF 24) on vegetative growth parameters of muskmelon under green house conditions: The CP (50, 100, 150 and 200 μg mL–1) and SDW treated susceptible seeds were sown in pots containing soil, sand and farmyard manure (FYM) in the ratio 2:1:1. Total 4 seeds were sowed to each pot and watered regularly up to 30 days. Growth promotion was evaluated under green house conditions by recording root length, shoot length, plant height, root and shoot fresh weight and dry weight, number of leaves per plant and chlorophyll content according to the method of Hiscox and Israelstam31. The experiment was repeated thrice with four replicates of each treatment. Total chlorophyll in the sample was calculated using the following formula:
Where:
| A | : | Absorbance at specific wave length (645 and 663 nm) |
| V | : | Final volume of the chlorophyll extract (mL) |
| W | : | Fresh weight of the sample (g) |
| a | : | Path length of light (cm) |
Effect of seed treatment with Crude Proteins (CP) from Penicillium verruculosum (MRS-PGPF 24) on muskmelon gummy stem blight disease protection under green house conditions: Plants raised from seeds primed with CP were challenge inoculated with pathogen after first true leaf emerges (15 days old). Disease (%) was recorded periodically (every 7 days) by observing the number of infected leaves26. The seedlings raised from SDW treated seeds and fungicide treated seeds served as negative and positive control respectively. The experiment was repeated thrice with four replicates of each treatment. Disease protection (DP) of S. cucurbitacearum was calculated using the formula:
where, C represents disease (%) incidence in control, T represents disease (%) incidence in treated plants.
Statistical analysis: Each experimental data was subjected to analysis of variance (one way ANOVA) using SPSS Inc., 16.0. Significant effects of treatments were determined by the magnitude of the F-value (p<0.05). Treatment means were separated by Tukey’s HSD test.
RESULTS
Effect of seed treatment with Crude Proteins (CP) from PGPF (MRS-PGPF 24) on seed germination and seedling vigor of muskmelon under in vitro conditions: Crude protein elicitors were tested for their effect on seed quality parameters in different concentrations and time intervals as explained in methodology. The seeds treated with elicitor recorded higher seed germination and seedling vigor when compared to control set. However, the enhancement of seed germination (%) and seedling vigor varied with different concentrations tested at two time intervals 3 h (Fig. 1a) and 6 h (Fig. 1b). Among them CP at 100 μg mL–1 concentration recorded highest seed germination of 90% and 2329.11 seedling vigor, followed by 150 μg mL–1 with 86.25% seed germination, 2242.23 seedling vigor at 6 h treatment (Fig. 1b).
Effect of seed treatment with Crude Proteins (CP) from PGPF (MRS-PGPF 24) on vegetative growth parameters of muskmelon under green house conditions: All the four CP elicitor concentrations were tested for enhanced growth parameters under green house conditions. The vegetative growth parameters such as plant height, number of leaves, shoot fresh and dry weight, root fresh and dry weight and total chlorophyll content were significantly enhanced on treatment with elicitors at the end of the experimental period.
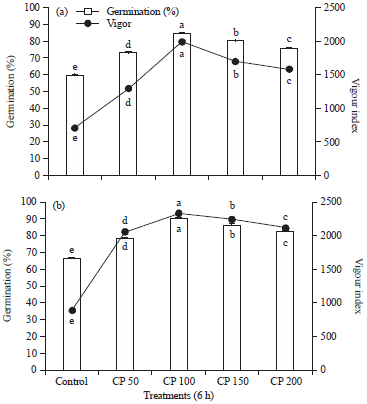 | |
| Fig. 1(a-b): | Effect of seed treatment with crude protein elicitors on gummy stem blight disease protection of muskmelon under green house conditions. (a) Crude protein treatment at 3 h, (b) Crude protein treatment at 6 h. |
| Values are means of four independent replicates. Vertical bars indicate±SE. Means followed by the same letter(s) within the same column are not significantly (p<0.05) different according to Tukey’s HSD | |
However, the level of growth promotion varied with the concentration of elicitors. Among the elicitor treatments, CP at 100 μg mL–1 concentration treated seeds recorded highest vegetative growth parameters with 42.35 cm plant height, 5 number of leaves, shoot fresh weight of 2.3 g, shoot dry weight of 1.30 g, root fresh weight of 0.43 g, root dry weight of 0.05 g and chlorophyll content of 4.16 mg g–1. The SDW treated seeds showed 21 cm height, 3 number of leaves, 0.5 g shoot fresh weight, 0.04 g shoot dry weight, 0.10 g root fresh weight, 0.01 g root dry weight and 0.05 mg g–1 chlorophyll content (Table 1).
Effect of seed treatment with Crude Proteins (CP) from PGPF (MRS-PGPF 24) on muskmelon gummy stem blight disease protection under green house conditions: The crude protein elicitor concentrations which offered best results than control for seed germination and seedling vigor was further tested for its efficacy to induce resistance in muskmelon against gummy stem blight disease under green house conditions.
| Table 1: | Effect of crude protein elicitors on growth of muskmelon seedlings under green house conditions at 6 h |
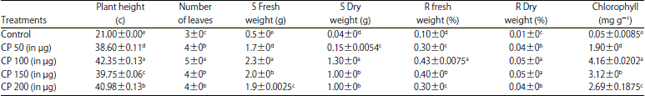 | |
| Values are means of four independent replicates. ±Indicate standard errors. Means followed by the same letter(s) within the same column are not significantly (p<0.05) different according to Tukey’s HSD | |
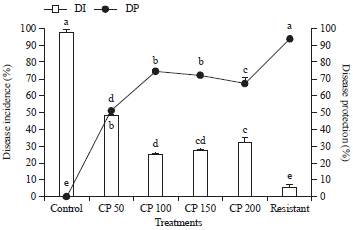 | |
| Fig. 2: | Effect of Seed treatment with crude protein elicitors on gummy stem blight disease protection of muskmelon under greenhouse conditions. |
| Values are means of four independent replicates. Vertical bars indicate±SE. Means followed by the same letter(s) within the same column are not significantly (p<0.05) different according to Tukey’s HSD | |
Here resistant treatment with Mancozeb fungicide was used as positive control. It was observed that fungicide treatment offered a maximum of 94.03% disease protection, whereas CP at 100 μg mL–1 concentration recorded 74.49% disease protection (Fig. 2). Among the four concentrations tested CP at 100 μg mL–1 recorded potential results in inducing disease resistance under green house conditions.
DISCUSSION
Rhizospheric fungi are studied for their growth promotion abilities and induction of resistance against wide array of pathogens. There is a continuous antagonistic war between the useful nonpathogenic microbes with the pathogens. This mechanism also holds good when tested directly as the seed inoculum of such beneficial organisms. In the preliminary studies various such beneficial fungi from healthy muskmelon rhizospheric soil were isolated to evaluate and identify their protective nature towards plants health. Among the isolates P. verruculosum was found to be very effective in promoting the muskmelon growth and also induction of resistance against gummy stem blight disease. These pathogens and rhizospheric fungal isolates were deposited in GenBank and obtained the accession numbers KJ782214 and KU645999, respectively. This was further selected to identify the compound that is responsible for this beneficial character of the fungus. Similar studies were conducted by various researchers against different host pathogen systems using soil borne fungi and bacteria32-37.
Elicitors are molecules that induces some biochemical defense responses during the infection by pathogens in resistant hosts. These molecules are studied extensively to understand the resistance mechanism in the host plants and also to induce resistance against array of disease causing organisms in plants3,38,37. In the present study crude protein elicitors from the culture filtrate of P. verruculosum seeds were primed with four concentrations (50, 100, 150 and 200 μg mL–1), tested for growth promotion activities and resistance induction in muskmelon against gummy stem blight disease. Similar studies conducted by Wu et al.12, who reported the systemic resistance induced by cyclodipeptides, which sheds light on the potential of cyclodipeptides for the control of plant diseases.
Protein elicitors acts as signaling molecules during the pathogen entry and also they are the necessary components for the plant growth. Crude protein extracts were tested for growth parameter studies in muskmelon under in vitro and green house conditions. Among all the concentrations tested CP at 100 μg mL–1 concentration recorded highest seed germination of 90% and 2329.11 seedling vigor, at 6 h treatment in vitro and 42.35 cm plant height, 5 number of leaves, shoot fresh weight of 2.3 g, shoot dry weight of 1.30 g, root fresh weight of 0.43 g, root dry weight of 0.05 g and chlorophyll content of 4.16 mg g–1 at 6 h under greenhouse conditions. These results were better than the seeds treated with sterile distilled water. Similar studies were conducted by the similar result was reported by Nandini et al.32 in pearl millet and Anupama et al.34 in tomato, Abhayashree et al.36 in chilli with crude oligosaccharide elicitors. From these results it will be clear that not all the fungal borne chemical compounds will interfere with the plant growth, but they also enhance growth parameters in plants.
Plants have developed various weapons to fight against the un welcomed guests that affect their natural metabolic activities and growth. These weapons are in the form of mechanical barriers or chemical compounds that trigger the defense responses immediately. Proteins are one such elicitors act as defense reaction inducers in plants against entry of phytopathogens. Studies conducted by Wu et al.12 revealed that cyclo (L-Pro-L-Pro) and cyclo (D-Pro-D-Pro) (where Pro is proline) could induce defense responses and systemic resistance in Nicotiana benthamiana. Treatment with the two cyclodipeptides led to a reduction in disease severity by Phytophthora nicotianae and Tobacco mosaic virus (TMV) infections compared with controls. Similar research by Wang et al.39, explored the effects of protein and polysaccharide in Meyerozyma guilliermondii on active compounds in Glycyrrhiza uralensis Fisch adventitious roots. In their study, a responsive protein LSP1 was purified from the Meyerozyma guilliermondii since the excellent induction. The responsive protein LSP1 significantly activated the defense signaling, mitogen-activated protein kinases and extremely up-regulated the expression of defense-related genes and functional genes involved in glycyrrhizic acid biosynthesis. In the present study protein elicitors treated to the susceptible seeds of muskmelon to induce resistance against gummy stem blight disease. Along with the sterile distilled water treated negative control, fungicide (Mancozeb) treated seeds are also used as positive control to compare results of present study with crude protein elicitors.
CONCLUSION
From the results of this study, it can be concluded that crude proteins from P. verruculosum can be effective elicitor in inducing resistance against gummy stem blight disease. The results of the study showed that fungicide treatment offered a maximum of 94.03% disease protection, whereas CP at 100 μg mL–1 concentration recorded 74.49% disease protection.
SIGNIFICANCE STATEMENT
The study was aimed to find an ecofriendly alternative control strategy to the present chemical control method for GSB disease. The study exploits the disease controlling potential of the crude protein extracts from muskmelon rhizospheric fungus P. verruculosum. This elicitor can be further purified to identify the pure compound with antagonistic activity to enhance its disease protection over the systemic fungicides. This study not only control the major disease of muskmelon but also provides a better eco-friendly alternative to effectively control crop loss in muskmelon caused by GSB disease thereby reduced the environmental pollution.
ACKNOWLEDGMENT
The first author is thankful for the funding agency “Department of Science and Technology (DST), New Delhi, India” for awarding the DST- INSPIRE fellowship for our research work (Award No: IF120571). The authors are thankful to CSIR (RA-371016/2K13/1) and UGC for their financial assistance and also to Institution of Excellence (IOE) Project Authorities, University of Mysore, DST-FIST for the central instrumentation facilities.
REFERENCES
- Zhang, H., S. Dong, M. Wang, W. Wang and W. Song et al., 2010. The role of vacuolar processing enzyme (VPE) from Nicotiana benthamiana in the elicitor-triggered hypersensitive response and stomatal closure. J. Exp. Bot., 61: 3799-3812.
CrossRefDirect Link - Zuppini, A., B. Baldan, R. Millioni, F. Favaron, L. Navazio and P. Mariani, 2003. Chitosan induces Ca2+-mediated programmed cell death in soybean cells. New Pthythol., 161: 557-568.
Direct Link - Montesano, M., G. Brader and E.T. Palva, 2003. Pathogen derived elicitors: Searching for receptors in plants. Mol. Plant Pathol., 4: 73-79.
CrossRefDirect Link - Mishra, A.K., K. Sharma and R.S. Misra, 2009. Purification and characterization of elicitor protein from Phytophthora colocasiae and basic resistance in Colocasia esculenta. Microbiol. Res., 164: 688-693.
CrossRefDirect Link - Shao, M., J. Wang, R.A. Dean, Y. Lin, X. Gao and S. Hu, 2008. Expression of a harpin‐encoding gene in rice confers durable nonspecific resistance to Magnaporthe grisea. Plant Biotechnol. J., 6: 73-81.
CrossRefDirect Link - Wei, Z.M., R.J. Laby, C.H. Zumoff, D.W. Bauer, S.Y. He, A. Collmer and S.V. Beer, 1992. Harpin, elicitor of the hypersensitive response produced by the plant pathogen Erwinia amylovora. Science, 257: 85-88.
CrossRefDirect Link - Alfano, J. and A. Collmer, 1997. The type III (hrp) secretion pathway of plant pathogenic bacteria: Trafficking harpins, Avr proteins and death. Bacteriology, 179: 5655-5662.
PubMedDirect Link - Oh, J., J.G. Kim, E. Jeon, C.H. Yoo, J.S. Moon, S. Rhee and I. Hwang, 2007. Amyloidogenesis of type III-dependent harpins from plant pathogenic bacteria. J. Biol. Chem., 282: 13601-13609.
CrossRefDirect Link - Adam, A.L., S. Pike, M.E. Hoyos, J.M. Stone, J.C. Walker and A. Novacky, 1997. Rapid and transient activation of a myelin basic protein kinase in tobacco leaves treated with harpin from Erwinia amylovora. Plant Physiol., 115: 853-861.
CrossRefDirect Link - Kvitko, B.H., A.R. Ramos, J.E. Morello, H.S. Oh and A. Collmer, 2007. Identification of harpins in Pseudomonas syringae pv. tomato DC3000, which are functionally similar to HrpK1 in promoting translocation of type III secretion system effectors. J. Bacteriol., 189: 8059-8072.
CrossRefDirect Link - Peng, D.H., D.W. Qiu, L.F. Ruan, C.F. Zhou and M. Sun, 2011. Protein elicitor PemG1 from Magnaporthe grisea induces Systemic Acquired Resistance (SAR) in plants. Mol. Plant-Microbe Interactions, 24: 1239-1246.
CrossRefDirect Link - Wu, L., H. Wu, L. Chen, H. Zhang and X. Gao, 2017. Induction of systemic disease resistance in Nicotiana benthamiana by the cyclodipeptides cyclo (l‐Pro‐l‐Pro) and cyclo (d‐Pro‐d‐Pro). Mol. Plant Pathol., 18: 67-74.
CrossRefDirect Link - Keinath, A.P., M.W. Farnham and T.A. Zitter, 1995. Morphological, pathological and genetic differentiation of Didymella bryoniae and Phoma spp. isolated from cucurbits. Phytopathology, 85: 364-369.
Direct Link - Frantz, J.D. and M.M. Jahn, 2004. Five independent loci each control monogenic resistance to gummy stem blight in melon (Cucumis melo L.). Theor. Applied Genet., 108: 1033-1038.
CrossRefDirect Link - Amand, P.C.S. and T.C. Wehner, 1991. Crop loss to 14 diseases in cucumber in North Carolina for 1983 to 1988. Cucurbit Genet. Cooperative Rep., 14: 15-17.
Direct Link - Gusmini, G., T.L. Ellington and T.C. Wehner, 2003. Mass production of gummy stem blight spores for resistance screening. Cucurbit Genet. Cooperative Rep., 26: 26-30.
Direct Link - Gusmini, G., R. Song and T.C. Wehner, 2005. New sources of resistance to gummy stem blight in watermelon. Crop Sci., 45: 582-588.
CrossRefDirect Link - Wako, T., Y. Sakata, M. Sugiyama, T. Ohara, D. Ishiuchi and A. Kojima, 2001. Identification of melon accessions resistant to gummy stem blight and genetic analysis of the resistance using an efficient technique for seedling test. Acta Hortic., 588: 161-164.
CrossRefDirect Link - Tsutsumi, C.Y. and N. da Silva, 2004. Screening of melon populations for resistance to Didymella bryoniae in greenhouse and plastic tunnel conditions. Braz. Arch. Biol. Technol., 47: 171-177.
CrossRefDirect Link - Keinath, A.P., 2011. From native plants in central Europe to cultivated crops worldwide: The emergence of Didymella bryoniae as a cucurbit pathogen. HortScience, 46: 532-535.
Direct Link - Thomas, A., D.B. Langston Jr., H.F. Sanders and K.L. Stevenson, 2012. Relationship between fungicide sensitivity and control of gummy stem blight of watermelon under field conditions. Plant Dis., 96: 1780-1784.
CrossRefDirect Link - Keinath, A.P., 2013. Susceptibility of cucurbit rootstocks to Didymella bryoniae and control of gummy stem blight on grafted watermelon seedlings with fungicides. Plant Dis., 97: 1018-1024.
CrossRefDirect Link - Zhao, J., Q.H. Xue, G.H. Shen, L. Xue, J.L. Duan and D.S. Wang, 2012. Evaluation of Streptomyces spp. for biocontrol of gummy stem blight (Didymella bryoniae) and growth promotion of Cucumis melo L. Biocontrol Sci. Technol., 22: 23-37.
CrossRefDirect Link - Koike, N., M. Hyakumachi, K. Kageyama, S. Tsuyumu and N. Doke, 2001. Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi: Lignification and superoxide generation. Eur. J. Plant Pathol., 107: 523-533.
CrossRefDirect Link - Djonovic, S., W.A. Vargas, M.V. Kolomiets, M. Horndeski, A. Wiest and C.M. Kenerley, 2007. A proteinaceous elicitor Sm1 from the beneficial fungus Trichoderma virens is required for induced systemic resistance in maize. Plant Physiol., 145: 875-889.
CrossRefDirect Link - Singh, S.D. and R. Gopinath, 1985. A seedling inoculation technique for detecting downy mildew resistance in pearl millet. Plant Dis., 69: 582-584.
Direct Link - Abdul-Baki, A.A. and J.D. Anderson, 1973. Vigor determination in soybean seed by multiple criteria. Crop Sci., 13: 630-633.
CrossRefDirect Link - Hiscox, J.D. and G.F. Israelstam, 1979. A method for the extraction of chlorophyll from leaf tissue without maceration. Can. J. Bot., 57: 1332-1334.
CrossRefDirect Link - Nandini, B., P. Hariprasad, S.R. Niranjana, H.S. Shetty and N.P. Geetha, 2013. Elicitation of resistance in pearl millet by oligosaccharides of Trichoderma spp. against downy mildew disease. J. Plant Interactions, 8: 45-55.
CrossRefDirect Link - Jogaiah, S., M. Abdelrahman, L.S.P. Tran and I. Shin-Ichi, 2013. Characterization of rhizosphere fungi that mediate resistance in tomato against bacterial wilt disease. J. Exp. Bot., 64: 3829-3842.
CrossRefDirect Link - Anupama, N., M. Murali, J. Sudisha and K.N. Amruthesh, 2014. Crude oligosaccharides from Alternaria solani with Bacillus subtilis enhance defense activity and induce resistance against early blight disease of tomato. Asian J. Sci. Technol., 5: 412-416.
Direct Link - Murali, M. and K.N. Amruthesh, 2015. Plant growth-promoting fungus Penicillium oxalicum enhances plant growth and induces resistance in pearl millet against downy mildew disease. J. Phytopathol., 163: 743-754.
CrossRefDirect Link - Abhayashree, M.S., M. Murali, M.C. Thriveni, G.M. Sindhu and K.N. Amruthesh, 2017. Crude oligosaccharides mediated resistance and histo-chemical changes in Capsicum annuum against anthracnose disease caused by Colletotrichum capsici. Plant Biosyst., 151: 221-233.
CrossRefDirect Link - Hossain, M.M., F. Sultana and M. Hyakumachi, 2017. Role of ethylene signalling in growth and systemic resistance induction by the plant growth‐promoting fungus Penicillium viridicatum in Arabidopsis. J. Phytopathol., 165: 432-441.
CrossRefDirect Link - Adrian, M., M. Lucio, C. Roullier-Gall, M.C. Heloir and S. Trouvelot et al., 2017. Metabolic fingerprint of PS3-induced resistance of grapevine leaves against Plasmopara viticola revealed differences in elicitor-triggered defenses. Frontiers Plant Sci., Vol. 8.
CrossRef - Wang, J., J. Li, J. Li, J. Li, S. Liu and W. Gao, 2017. LSP1, a responsive protein from Meyerozyma guilliermondii, elicits defence response and improves glycyrrhizic acid biosynthesis in Glycyrrhiza uralensis Fisch adventitious roots. J. Cell. Physiol., 232: 3510-3519.
CrossRefDirect Link