
Ahsan A. Kadhimi
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, Malaysia
LiveDNA: 964.12048
Che Radziah Che Mohd Zain
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, Malaysia
Arshad Naji Alhasnawi
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, Malaysia
LiveDNA: 60.12099
Anizan Isahak
School of Environmental Science and Natural Resources, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, Malaysia
Mehdi Farshad Ashraf
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, Malaysia
Azhar Mohamad
Malaysian Nuclear Agency, Malaysia
Febri Doni
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, Malaysia
Wan Mohtar Wan Yusoff
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, Malaysia
Asian Journal of Crop Science
Year: 2016 | Volume: 8 | Issue: 2 | Page No.: 52-59







ABSTRACT
Background: A drought-tolerant genotype should be identified and improved for the continuity of food security because of the global importance of rice. Mutation and tissue culture techniques are effective and important for studies on the development of potential rice resources. The effects of irradiation and drought stress on rice genotype MR269 of Oryza sativa L., differing in drought tolerance capability were investigated. Methodology: The seeds of the genotypes were exposed to gamma rays at 350 Gy. Naked seeds were initiated onto half-strength standard MS medium enhanced with 0, 10 and 20% polyethylene glycol (PEG) 6000. The effects of PEG on root and shoot length, fresh and dry seedling weights, total chlorophyll and carbohydrate contents and proline accumulation were examined. Significant differences were observed in the studied traits. The non-irradiated seeds presented the highest mean shoot and seedling fresh weight, whereas the irradiated seeds presented the largest root length and highest proline concentrations. Results: The PEG produced a decrease in the means of all the characteristics investigated by enhancing PEG concentration, except for the means of proline content, which increased with the increase in PEG concentration. The irradiated genotypes produced the highest values of root length, dry weight, chlorophyll, proline and carbohydrates in concentrations of 20% PEG compared with the non-irradiated ones. Conclusion: Thus, a positive signal exists in the effect of radiation on these characteristics in drought tolerance. Research on the improvement of a drought-tolerant genotype under different drought conditions is necessary to enhance rice productivity and food security.
PDF Abstract XML References Citation
Received: January 30, 2016;
Accepted: February 16, 2016;
Published: March 15, 2016
How to cite this article
Ahsan A. Kadhimi, Che Radziah Che Mohd Zain, Arshad Naji Alhasnawi, Anizan Isahak, Mehdi Farshad Ashraf, Azhar Mohamad, Febri Doni and Wan Mohtar Wan Yusoff, 2016. Effect of Irradiation and Polyethylene Glycol on Drought
Tolerance of MR269 Genotype Rice (Oryza sativa L.). Asian Journal of Crop Science, 8: 52-59.
DOI: 10.3923/ajcs.2016.52.59
URL: https://scialert.net/abstract/?doi=ajcs.2016.52.59
DOI: 10.3923/ajcs.2016.52.59
URL: https://scialert.net/abstract/?doi=ajcs.2016.52.59
INTRODUCTION
Rice is the most significant food crop in the world and the main source of nutrition for approximately half of the world’s population1. The rice-growing parts of Asia, which comprise 130 million hectares are frequently endangered through severe abiotic pressures; most are generally deficient. These parts contain irrigated and rain-fed plains, which account for more than 85% of the entire rice supply in the world2. The improvement of drought-resistant cultivars and lines of rice over assortment and breeding is financially significant in enhancing rice manufacturers in regions with low rainfall or without any proper irrigation scheme3. Several effective drought-tolerant genotypes existed; however, these were amended because it was a significant goal of plant breeders for a long period4. Conventional breeding efforts are time, cost and worker-intensive. Biotechnology like tissue culture technology offer rapid alternative in crop improvement. Increased crop yield in a water-deficient environment may be achieved by breeding crops that are highly tolerant to drought, this is also an easy alternative in crop improvement. In recent decades, tissue culture assortment has emerged as a possible and cost-effective instrument to develop stress-tolerant plants. Plant tolerance for both biotic and abiotic stress may be enhanced by selecting particular agents, such as NaCl for salt tolerance and polyethylene glycol (PEG) for drought tolerance5. The PEG is a polymer compound with several functions. It has a large molecular weight, does not infiltrate plant tissues and is a perfect osmoticum for application in a hydroponics root mean6-8. Joshi et al.9 studied the reaction of several varieties of rice (Oryza sativa) callus to different amounts of PEG-induced water stress; callus growth, water substance and proline accretion were considered. The PEG has been extensively applied to induce water stress. Moreover, drought-tolerant cultivars have been achieved in several crops by expanding approaches derived from PEG application10. In most of these cases, PEG is utilized to induce drought.
Mutation induction is a means to create changes in a crop genotype. It offers the likelihood of incorporating anticipated qualities that cannot be articulated in nature or have disappeared during growth. Irradiation is more effective in inducing mutations in plants compared with other methods11. Many studies documented that the variation in rice genotypes through physiological criteria could be utilized as a tool in plant breeding. Meanwhile, data on the physiological mechanisms underlying plant responses to drought tolerance for MR269 remain insufficient. Thus, an attempt is made in this study to investigate the effect of irradiation and PEG on several biochemical and morphological characteristics in genotype MR269 and to identify a criterion for drought tolerance.
MATERIALS AND METHODS
Seeds of genotypes of local rice (MR269) were exposed to a dose (350 Gy) of gamma ray (Caesium-137) in the Malaysian Nuclear Agency12. The seeds were fully washed with distilled water three times. They were then washed with 70% ethanol for 2-3 min. In a laminar airflow cabinet, the seeds were treated with 0.1% mercury chloride (HgCl2) with the addition of a few drops of tween-20 for internal surface sterilization for 4-6 min. Finally, the seeds were washed with sterile distilled water several times to remove all sterilizing agents and were germinated in a flask inside an incubator at 25°C for 7 day. The seedlings were transferred to test tubes (15×2.5 cm) containing half-strength MS medium supplemented with 0, 10 and 20% PEG 6000. The test tubes were placed in a growth chamber at 16/8 h (light/dark) photoperiod at a light intensity of 1000 lux and ambient temperature of 25±2°C. The lengths of the shoots and roots as well as plant fresh weights were recorded after 4 weeks of seed germination. Plant dry weights were recorded after drying plant tissues for 48 h at 72°C13,14.
Determination of proline concentration in rice seedlings: Proline concentrations were determined according to the method of15. Plant seedlings with 5 mg fresh weight were homogenized with 3% sulfosalicylic acid. The filtrate was mixed with 2 mL of glacial acetic acid and ninhydrin reagent and incubated at 100°C for 30 min. The samples were rigorously mixed with 4 mL toluene. The light absorption of the toluene phase was estimated at 520 nm through the use of a spectrophotometer. Proline concentration was determined and expressed as micromolar per gram fresh weight.
Determination of carbohydrate concentrations in rice seedlings: Total sugar content (carbohydrate concentrations) was determined without the identification of specific sugar components based on the method of phenol sulfuric acid16. A total of 10 mg dried seedlings were homogenized with deionized water. The extract was filtered and then treated with 1 mL of 5% phenol and 1 mL of 98% sulfuric acid. The mixture was incubated at 30°C for 20 min. Then, its absorbance at 485 nm was determined with a spectrophotometer. The concentrations of soluble sugar were expressed as microgram per gram dry weight.
Experimental design and statistical analysis: The experiment was designed as a factorial experiment based on a CRD (completely randomized design). The factors included Gamma irradiation (2 level, with and without irradiation) and PEG (3 level: 0, 10 and 20%). In this study, three replication with 10 samples per each replication were applied. The data were subjected to the normality test prior to analysis of variance using SAS programme (Release 9.1 for Windows, SAS Institute Inc., Cary, NC, USA) software. Significant differences among the mean values of treatments were determined using the Duncan’s Multiple Range Test (DMRT) and the Least Significant Difference was calculated at the α = 0.05 level.
RESULTS
Length of shoots: Figure 1 shows that the mean value of shoot length increased significantly in the non-irradiated seedlings compared with the irradiated ones. The data in Fig. 1 shows that application of PEG during seedling growth decreased the shoot length irrespective of gamma ray application.
The interaction between irradiation and PEG% revealed that the largest shoot length of 17.4 cm was recorded for non-irradiation treatment with PEG 0.0% compared with irradiation treatment, which reached 15.5 cm. The non-irradiated treated sample produced the largest shoot length in concentrations of 20% PEG which reached 13.75 cm compared with the irradiated samples, which reached 9.5 cm.
Length of roots: The results displayed in Fig. 2 shows that the mean value of root length increased significantly in the irradiated samples compared with that in the non-irradiated ones. A significant decrease occurred in the length of the roots at 10 and 20% PEG for non-irradiated and irradiated genotypes compared with 0.0% PEG (Fig. 2). The interaction between irradiation and PEG% showed larger root length in 20% PEG with irradiation than in the non-irradiation treatment with the same concentration. According to the data, both genotypes exhibited a similar response under PEG stress for both irradiated and non-irradiated treatments. Plant height and root length decreased with increased PEG concentration.
Seedling fresh weight: According to Fig. 3, the mean fresh weight increased significantly in the non-irradiated sample compared with irradiated MR269. The data indicated that a similar response was generated under PEG stress by both irradiated and non-irradiated samples. Fresh weight decreased with increased PEG concentration.
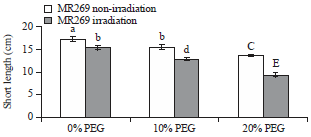 | |
| Fig. 1: | Mean values of shoot length of MR269 plants obtained from seeds subjected to gamma ray at 0 Gy (not irradiated) and 350 Gy (irradiated) and PEG treatments at concentrations of 0, 10 and 20% |
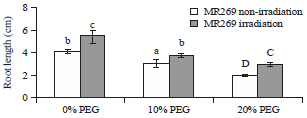 | |
| Fig. 2: | Mean values of root length of MR269 plants obtained from seeds subjected to gamma ray at 0 Gy (not irradiated) and 350 Gy (irradiated) and PEG treatments at concentrations of 0, 10 and 20% |
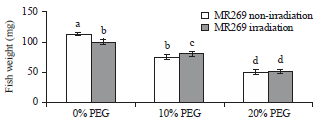 | |
| Fig. 3: | Mean values of seedling fresh weight of MR269 plants obtained from seeds subjected to gamma ray at 0 Gy (not irradiated) and 350 Gy (irradiated) and PEG treatments at concentrations of 0, 10 and 20% |
The interaction between irradiation and PEG% revealed that higher fresh weight (112.59 mg) was recorded for non-irradiation treatment with PEG 0.0% than for irradiation treatment (100.3 mg) at the same concentration. Meanwhile, no significant differences were observed between the non-irradiated and irradiated samples in concentrations of 10 and 20% PEG.
Seedling dry weight: Figure 4 shows that no significant differences in mean dry weight were observed in the non-irradiated and irradiated samples. Moreover, Fig. 4 reveals that the mean dry weight decreased significantly at 10 and 20% PEG compared with 0.0% PEG. The interaction between irradiation and PEG% irradiation treatments revealed a higher dry weight in MR269 non-radiation treatment at 0.0% PEG concentration than in irradiation treatment at the same PEG concentration.
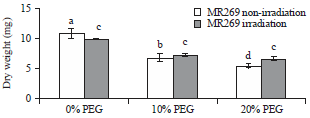 | |
| Fig. 4: | Mean values of seedling dry weight of MR269 plants obtained from seeds subjected to gamma ray at 0 Gy (not irradiated) and 350 Gy (irradiated) and PEG treatments at concentrations of 0, 10 and 20% |
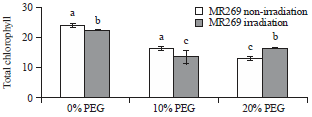 | |
| Fig. 5: | Total chlorophyll content of MR269 plants obtained from seeds subjected to gamma ray at 0 Gy (not irradiated) and 350 Gy (irradiated) and PEG treatments at concentrations of 0, 10 and 20% |
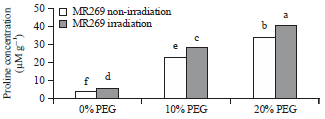 | |
| Fig. 6: | Mean values of seedling proline concentrations (μM g–1) of MR269 plants obtained from seeds subjected to gamma ray at 0 Gy (not irradiated) and 350 Gy (irradiated) and PEG treatments at concentrations of 0, 10 and 20% |
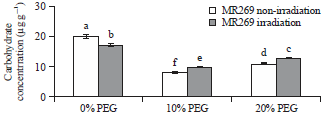 | |
| Fig. 7: | Mean values of carbohydrate concentrations (μg g–1) of MR269 plants obtained from seeds subjected to gamma ray at 0 Gy (not irradiated) and 350 Gy (irradiated) and PEG treatments at concentrations of 0, 10 and 20% |
The data showed a higher dry weight in MR269 radiation treatment at 20% PEG concentration than in non-irradiation treatment at the same concentration.
Total chlorophyll: Regardless of the applied PEG concentration, no significant differences in total chlorophyll content were observed in the non-irradiated and radiated samples. Figure 5 indicates that a significant decrease occurred in total chlorophyll at 10 and 20% PEG for non-irradiated and irradiated genotypes compared with 0.0% PEG. The interaction between irradiation and PEG% irradiation treatments showed higher total chlorophyll for irradiation treatment than for non-irradiation treatment at 20% PEG concentration.
Proline concentrations in seedlings: According to Fig. 6, the mean value of proline concentrations increased significantly in the irradiated genotypes compared with the non-irradiated ones. The proline concentrations exhibited a significant increase at 10 and 20% PEG compared with 0.0% PEG. The interaction between irradiation and PEG% showed higher proline concentrations in the radiation treatment at 0.0% PEG than in the non-irradiation treatment at 0.0% PEG. The data showed higher proline concentrations in the radiation treatment at 20% PEG concentration than in the non-irradiation treatment at the same concentration.
Carbohydrate concentrations: According to Fig. 7, the mean carbohydrate concentrations increased significantly in the non-irradiated samples compared with the irradiated ones. Figure 7 also indicates that a significant decrease occurred in carbohydrate concentrations at 10 and 20% PEG concentrations for both non-irradiated and irradiated samples compared with 0.0% PEG. The interaction between irradiation and PEG% showed higher carbohydrate concentrations in the non-radiation treatment at 0.0% PEG compared with the irradiation treatment at 0.0% PEG. The data showed higher carbohydrate concentrations in the radiation treatment at 10 and 20% PEG concentrations compared with the non-irradiation treatment at the same concentrations.
DISCUSSION
Rice is highly susceptible to water stress17,18 and the magnitude of yield losses depends on the duration of water stress and stage of crop growth. The large reduction in all characteristics with the increase in PEG percentage, except for the mean values of proline concentration was in line with the results of Soni et al.19 who observed a decrease in root and shoot fresh weight under stressed circumstances in all investigated genotypes. Additionally, most of the tolerant genotypes gathered more dry matter than the vulnerable ones. By adding PEG to the culture medium, the water of the medium decreased20-21. Physiological procedures might rely on cell capacity mainly in plants sensitive to water shortfall. Moreover, the decrease in seedling fresh and dry weight with cumulative PEG levels might have occurred because of the osmotic pressure influencing the accumulation of soluble elements in cells; consequently, it affected the other physiological functions in the cells22. Growth is accomplished through cell enlargement and cell division and includes genetic variation, physiological, ecological and morphological actions and related complicated relations. The extent and success of plant development are based on actions under the influence of drought. Cell development is considered one of the most drought-sensitive physiological procedures because of the decrease in turgor pressure23. Under water shortage, cell extension in developed plants may be hampered by the disruption of the water flux from the xylem to the nearby extension cells24. Simultaneously, a significant reduction occurred in seedling shoot length with the increase in PEG percentage compared with the control (Fig. 1).
Figure 2 shows that MR269 irradiation can maintain the root length even at high PEG concentrations. Such genotype might be useful for future activities to improve drought tolerance. Pattanagul and Thitisaksakul25, stated that root development is a general indicator of the response to stress conditions. Even when leaves cease growing, photosynthesis continues26. Likewise, Jaleel et al.27 stated that a ramified root system is associated with drought tolerance and high biomass mainly because of its capability to extract more water from soil and its transference to aboveground parts for photosynthesis. Hence, drought inhibits plant cell division and improvement because of its influence on physiological procedures. The root length at initial development phases is considered a good criterion for selection under stress conditions.
The non-irradiated genotypes presented the maximum shoot length and seedling fresh and dry weights at 0 and 10% PEG (Fig. 1, 3 and 4). The irradiated genotypes presented the maximum values of root length and dry weight at 20% PEG (Fig. 2 and 4). The physiological significance of gamma radiation is due to the formation of free radicals over the hydrolysis of water that can haste to the modulation of an antioxidative scheme, accumulation of phenolic combinations and chlorophyll pigments affecting shoot and root distance and fresh and dry heaviness28,29.
Drought enhances senescence by alleviating chlorophyll degradation, nitrogen damage and lipid peroxidation10. Drought decreases photosynthesis by decreasing stomatal and mesophyll conductance30. Figure 5 shows that the total chlorophyll decreased with increased PEG concentration. This finding is in line with that of researchers who found that osmotic stress generated by (PEG-6000) usually reduces the photosynthetic rate. Exposure to drought stress affects chlorophyll content31. From the physiological viewpoint, the leaf chlorophyll content is associated with the photosynthetic degree. The decrease in chlorophyll standards because of drought stress could be related to the typical symptom of oxidative stress and could be the result of pigment photo-oxidation and chlorophyll degradation24. The opposing consequence of water shortfall stress on chlorophyll content and comparative water content overlaps with the general trend of the consequence of drought on physiological procedures in plants. Almeselmani et al.32 showed that the water content of wheat leaves decreases with increased water stress, such a decrease adversely affects the development and harvest of the plant. Likewise, drought has been found to reduce the chlorophyll content in drought-stressed cotton33,34.
The non-irradiated cultivars showed the highest content of chlorophyll, carbohydrates and proline in 0 and 10% PEG. Meanwhile, the irradiated cultivars presented the highest content of chlorophyll, carbohydrates and proline in 20% PEG (Fig. 5-7). Gamma rays are imperative in improving the development of genetic variability35. They usually affect plant growth by inducing cytological, genetically, biochemical, physiological and morphogenetic variations in cells and tissues36. However, high doses of gamma ray (2-20 kGy) applied to the seed earlier than sowing change the enzyme action37-41. Photosynthetic pigments might be destroyed by high doses of gamma irradiation and with the concomitant damage of photosynthetic volume42. Gamma radiation can develop stress resistance by affecting the antioxidant enzyme movement and photosynthetic volume43. This result is in line with that of Baek et al.44, who found that gamma radiation can affect the osmotic potential because of the superior proline content. Drought stress induces several biochemical responses in crops. These responses may sometimes help plants adapt to limiting environmental conditions45. Plants exposed to water stress accumulate molecules, such as proline, in their tissues. Proline is a non-protein amino acid that accumulates in the majority of tissues related to water stress; it is simply metabolized upon retrieval from water insufficiency46. This accumulation can be considered a means of osmotic modification that protects plant tissues from destructive consequences related to water stress. However, Kanawapee et al.47 reported that increased proline concentration during stress conditions works as an osmoticum, a desiccation-protectant and a sink for nitrogen and reduces power during stress. The physiological characteristic of proline in preserving tolerance to drought might be starvation evasion. Consequently, a dominance of anabolic actions is perhaps the key issue regarding drought tolerance. These results (Fig. 6) show that proline accumulated in an upper level below PEG stress in irradiation, which indicates the plant’s answer to maintaining osmotic homeostasis. Datta et al.48 reported that the enhanced level of proline in leaves under salt stress might be due to synthesis or the breakdown of proteins during stress. Solute accumulation is also one of the mechanisms for drought tolerance in rice49.
Unlike in 0.0% PEG, a significant decrease in carbohydrate concentration was observed in 10% PEG (Fig. 7). Plant cells may utilize a large amount of energy, including carbohydrates, to tolerate stress50. A significant increase in carbohydrate concentration at 10 and 20% PEG was observed in the irradiated cultivars compared with 10 and 20% PEG in the non-irradiated ones. This increase can be explained by the decomposition of multiple sugars into unilateral and bilateral sugars in the case of water shortage. In this condition, it works with proline to organize the cells in osmotic stress. Pattanagul51, also observed an increase in carbohydrate concentrations under drought conditions. Additionally, the results are consistent with those of52,53.
CONCLUSION
A significant reduction in the studied traits was observed, except for proline content, which increased with increased PEG percentage. The non-irradiated genotypes had the highest traits in 0% PEG. The irradiated genotypes had the largest value of root length, dry weight, chlorophyll, proline and carbohydrates in concentrations of 20% PEG compared with the non-irradiated ones. Hence, a positive signal exists regarding the effect of radiation on these characteristics in drought tolerance.
ACKNOWLEDGMENTS
Special thanks to Ministry of Higher Education and Scientific Research Iraq, for financial supports and moral support and special thanks to Ministry of Higher Education Malaysia for financial funding. To complete this search, under Grant FRGS/2/2014/STWNO3/UKM/01/1 are acknowledged. Also to Nuclear Malaysia Agency for providing facility for acute gamma radiation.
REFERENCES
- Hadiarto, T. and L.S.P. Tran, 2011. Progress studies of drought-responsive genes in rice. Plant Cell Rep., 30: 297-310.
CrossRefDirect Link - Kadhimi, A.A., A.N. Alhasnawi, I. Anizan, M.F. Ashraf and A. Mohamad et al., 2014. Use of biotechnology to improve the tolerance in rice (Oryza sativa) to drought stress. J. Pure Applied Microbiol., 8: 4001-4010.
Direct Link - Zheng, X., L. Chen, M. Li, Q. Lou and H. Xia et al., 2013. Transgenerational variations in DNA methylation induced by drought stress in two rice varieties with distinguished difference to drought resistance. PLoS ONE, Vol. 8.
CrossRefDirect Link - Rao, S. and F.T.Z. Jabeen, 2013. In vitro selection and characterization of polyethylene glycol (PEG) tolerant callus lines and regeneration of plantlets from the selected callus lines in sugarcane (Saccharum officinarum L.). Physiol. Mol. Biol. Plants, 19: 261-268.
CrossRefDirect Link - Michel, B.E. and M.R. Kaufmann, 1973. The osmotic potential of polyethylene glycol 6000. Plant Physiol., 51: 914-916.
CrossRefPubMedDirect Link - Money, N.P., 1989. Osmotic pressure of aqueous polyethylene glycols: Relationship between molecular weight and vapor pressure deficit. Plant Physiol., 91: 766-769.
CrossRefDirect Link - Patade, V.Y., S. Bhargava and P. Suprasanna, 2011. Salt and drought tolerance of sugarcane under iso-osmotic salt and water stress: Growth, osmolytes accumulation and antioxidant defense. J. Plant Interact., 6: 275-282.
CrossRefDirect Link - Joshi, R., A. Shukla and R. Sairam, 2011. In vitro screening of rice genotypes for drought tolerance using polyethylene glycol. Acta Physiol. Plant., 33: 2209-2217.
CrossRefDirect Link - Swain, P., M. Anumalla, S. Prusty, B.C. Marndi and G.J.N. Rao, 2014. Characterization of some Indian native land race rice accessions for drought tolerance at seedling stage. Aust. J. Crop Sci., 8: 324-331.
Direct Link - El-Degwy, I.S., 2013. Mutation induced genetic variability in rice (Oryza sativa L.). Int. J. Agric. Crop Sci., 5: 2789-2794.
Direct Link - Kadhimi, A.A., A.N. Alhasnawi, A. Isahak, M.F. Ashraf, A. Mohamad, W.M.W. Yusoff and C.R.C.M. Zain, 2016. Gamma radiosensitivity study on MRQ74 and MR269, two elite varieties of rice (Oryza sativa L.). Life Sci. J., 13: 85-91.
Direct Link - Wani, S.H., P.A. Sofi, S.S. Gosal and N.B. Singh, 2010. In vitro screening of rice (Oryza sativa L.) callus for drought tolerance. Commun. Biometry Crop Sci., 5: 108-115.
Direct Link - Prajuabmon, A., P. Theerakulpisut, B. Kijwijan and N. Muangsan, 2009. In vitro investigation on salt tolerant characteristics of rice seedlings (Oryza sativa L.). Res. J. Agric. Biol. Sci., 5: 423-427.
Direct Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Zain, N.A.M., M.R. Ismail, M. Mahmood, A. Puteh and M.H. Ibrahim, 2014. Alleviation of water stress effects on MR220 rice by application of periodical water stress and potassium fertilization. Molecules, 19: 1795-1819.
CrossRefDirect Link - Pandey, A., A. Kumar, D.S. Pandey and P.D. Thongbam, 2014. Rice quality under water stress. Indian J. Adv. Plant Res., 1: 23-26.
Direct Link - Soni, P., M. Rizwan, K.V. Bhatt, T. Mohapatra and G. Singh, 2011. In-vitro response of Vigna aconitifolia to drought stress induced by peg 6000. J. Stress Physiol. Biochem., 7: 108-121.
Direct Link - Anjum, S.A., X.Y. Xie, L.C. Wang, M.F. Saleem, C. Man and W. Lei, 2011. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res., 6: 2026-2032.
Direct Link - Farooq, M., A. Wahid, N. Kobayashi, D. Fujita and S.M.A. Basra, 2009. Plant drought stress: Effects, mechanisms and management. Agron. Sustainable Dev., 29: 185-212.
CrossRefDirect Link - Pattanagul, W. and M. Thitisaksakul, 2008. Effect of salinity stress on growth and carbohydrate metabolism in three rice (Oryza sativa L.) cultivars differing in salinity tolerance. Indian J. Exp. Biol., 46: 736-742.
PubMedDirect Link - Davatgar, N., M.R. Neishabouri, A.R. Sepaskhah and A. Soltani, 2009. Physiological and morphological responses of rice (Oryza sativa L.) to varying water stress management strategies. Int. J. Plant Prod., 3: 19-32.
Direct Link - Jaleel, C.A., P. Manivannan, A. Wahid, M. Farooq, H.J. Al-Juburi, R. Somasundaram and R. Panneerselvam, 2009. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol., 11: 100-105.
Direct Link - Ashraf, M., 2009. Biotechnological approach of improving plant salt tolerance using antioxidants as markers. Biotechnol. Adv., 27: 84-93.
CrossRefPubMedDirect Link - Borzouei, A., M. Kafi, H. Khazaei, B. Naseriyan and A. Majdabadi, 2010. Effects of γ radiation on germination and physiological aspects of wheat (Triticum aestivum L.) seedlings. Pak. J. Bot., 42: 2281-2290.
Direct Link - Chutia, J. and P. Borah, 2012. Water stress effects on leaf growth and chlorophyll content but not the grain yield in traditional rice (Oryza sativa Linn.) genotypes of Assam, India II. Protein and proline status in seedlings under PEG induced water stress. Am. J. Plant Sci., Am. J. Plant Sci.,: 971-980.
CrossRefDirect Link - Almeselmani, M., F. Abdullah, F. Hareri, M. Naaesan, M.A. Ammar, O. ZuherKanbar and A.A. Saud, 2011. Effect of drought on different physiological characters and yield component in different varieties of syrian durum wheat. J. Agric. Sci., 3: 127-133.
CrossRefDirect Link - Massacci, A., S.M. Naviev, L. Pietrosanti, S.K. Nematov, T.N. Chernikova, K. Thor and J. Leipner, 2008. Response of the photosynthetic apparatus of cotton (Gossypium hirsutum) to the onset of drought stress under field conditions studied by gas-exchange analysis and chlorophyll fluorescence imaging. Plant Physiol. Biochem., 46: 189-195.
CrossRefPubMedDirect Link - Kiani, S.P., P. Moury, A. Sarrafi and P. Grieu, 2008. QTL analysis of chlorophyll fluorescence parameters in sunflower (Helianthus annuus L.) under well-watered and water-stressed conditions. Plant Sci., 175: 565-573.
CrossRefDirect Link - Jan, S., T. Parween, R. Hameed, T.O. Siddiqi and Mahmooduzzafar, 2013. Effects of presowing gamma irradiation on the photosynthetic pigments, sugar content and carbon gain of Cullen corylifolium (L.) Medik. Chilean J. Agric. Res., 73: 345-350.
CrossRefDirect Link - Jan, S., T. Parween, T.O. Siddiqi and Mahmooduzzafar, 2011. Gamma radiation effects on growth and yield attributes of Psoralea corylifolia L. with reference to enhanced production of psoralen. Plant Growth Regul., 64: 163-171.
CrossRefDirect Link - Al-Rumaih, M.M. and M.M. Al-Rumaih, 2008. Influence of ionizing radiation on antioxidant enzymes in three species of Trigonella. Am. J. Environ. Sci., 4: 151-156.
Direct Link - Vandenhove, H., N. Vanhoudt, J. Wannijn, M. Van Hees and A. Cuypers, 2009. Effect of low-dose chronic gamma exposure on growth and oxidative stress related responses in Arabidopsis thaliana. Radioprotection, 44: 487-491.
CrossRefDirect Link - Stajner, D., B. Popovic and K. Taski-Ajdukovic, 2009. Effects of γ-irradiation on antioxidant activity in soybean seeds. Cent. Eur. J. Biol., 4: 381-386.
CrossRefDirect Link - Jan, S., T. Parween, T.O. Siddiqi and Mahmooduzzafar, 2012. Effect of γ radiation on morphological, biochemical and physiological aspects of plants and plant products. Environ. Rev., 20: 17-39.
CrossRefDirect Link - Jan, S., T. Parween, T.O. Siddiqi and Mahmooduzzafar, 2012. Anti-oxidant modulation in response to gamma radiation induced oxidative stress in developing seedlings of Psoralea corylifolia L. J. Environ. Radioact., 113: 142-149.
CrossRefDirect Link - Ling, A.P.K., Y.C. Ung, S. Hussein, A.R. Harun, A. Tanaka and H. Yoshihiro, 2013. Morphological and biochemical responses of Oryza sativa L. (cultivar MR219) to ion beam irradiation. J. Zhejiang Univ. Sci. B, 14: 1132-1143.
CrossRefDirect Link - Kiong, A.L.P., A.G. Lai, S. Hussein and A.R. Harun, 2008. Physiological responses of Orthosiphon stamineus plantles to gamma irradiation. Am. Euras. J. Sustain. Agric., 2: 135-149.
Direct Link - Baek, M.H., B.Y. Chung, J.H. Kim, S.G. Wi, J.S. Kim and I.J. Lee, 2006. Gamma radiation and hormone treatment as tools to reduce salt stress in rice (Oryza sativa L.). J. Plant Biol., 49: 257-260.
CrossRefDirect Link - Shehab, G.G., O.K. Ahmed and H.S. El-Beltagi, 2010. Effects of various chemical agents for alleviation of drought stress in rice plants (Oryza sativa L.). Not. Bot. Hort. Agrobot. Cluj., 38: 139-148.
Direct Link - Singh, D.K., P.W.G. Sale, C.K. Pallaghy and V. Singh, 2000. Role of proline and leaf expansion rate in the recovery of stressed white clover leaves with increased phosphorus concentration. New Phytol., 146: 261-269.
CrossRefDirect Link - Kanawapee, N., J. Sanitchon, W. Lontom and P. Threerakulpisut, 2012. Evaluation of salt tolerance at the seedling stage in rice genotypes by growth performance, ion accumulation, proline and chlorophyll content. Plant Soil, 358: 235-249.
CrossRefDirect Link - Datta, J.K., S. Nag, A. Banerjee and N.K. Mondal, 2009. Impact of salt stress on five varieties of wheat (Triticum aestivum L.) cultivars under laboratory condition. J. Applied Sci. Environ. Manag., 13: 93-97.
Direct Link - Mostajeran, A. and V. Rahimi-Eichi, 2009. Effects of drought stress on growth and yield of rice (Oryza sativa L.) cultivars and accumulation of proline and soluble sugars in sheath and blades of their different ages leaves. Am.-Eurasian J. Agric. Environ. Sci., 5: 264-272.
Direct Link - Thakur, P.S. and V.K. Rai, 1980. Water stress effects on maize: Carbohydrate metabolism of resistant and susceptible cultivars of Zea mays L. Biol. Plant., 22: 50-56.
CrossRefDirect Link - Pattanagul, W., 2011. Exogenous abscisic acid enhances sugar accumulation in rice (Oryza sativa L.) under drought stress. Asian J. Plant Sci., 10: 212-219.
CrossRefDirect Link - Zhou, Y., H.M. Lam and J. Zhang, 2007. Inhibition of photosynthesis and energy dissipation induced by water and high light stresses in rice. J. Exp. Bot., 58: 1207-1217.
CrossRefDirect Link - Guo, C., X. Ge and H. Ma, 2013. The rice OsDIL gene plays a role in drought tolerance at vegetative and reproductive stages. Plant Mol. Biol., 82: 239-253.
CrossRefDirect Link