
D. Ramla
Biotechnology and Plant Breeding Division, National Institute of Agronomic Research of Algeria, Algeria
LiveDNA: 213.11960
M.S. Yakhou
National Institute of Agronomic Research of Algeria, Algeria
N. Bilek
National Institute of Agronomic Research of Algeria, Algeria
M. Hamou
National Institute of Agronomic Research of Algeria, Algeria
A. Hannachi
National Institute of Agronomic Research of Algeria, Algeria
A. Aissat
Department of Agronomy, University Saad Dahlab Blida, Algeria
L. Mekliche-Hanifi
Department of Plant Science, National Agronomic Graduate School, Belfort, El-Harrach, Algiers, Algeria
Asian Journal of Crop Science
Year: 2016 | Volume: 8 | Issue: 2 | Page No.: 43-51







ABSTRACT
Aim: Production of new barley (Hordeum vulgare L.) genotypes with stable grain yield is an important challenge in variable and harsh climatic conditions such semi-arid zones. Methodology: For this purpose, twenty-nine 6-row barley genotypes, 26 barley anther culture-derived doubled haploid lines obtained from F2 plants of 2 biparental crosses, along with 3 parental cultivars were tested for grain yield and stability level at five semi-arid environments in Algerian highlands barley grown areas. Several stability parameters such, regression coefficient (bi), deviation from regression (S2di), Pinthus’ coefficient (R2), environmental variance (S2i), coefficient of variation (CV%), Wricke’s genotypic ecovalence (Wi), Shukla’s stability variance (σi2), heterogeneity variance (%HV), incomplete correlation (%IC) and Plaisted’s stability Parameter (P), were used to assess the stability of each genotype. Results: Twenty eight genotypes showed a wide adaptability (bi = 1) and only a single line (DH40) showed a specific adaptation (bi<1). About 10 genotypes showed yield stability over the environments studied (S2di = 0, low values of Wi, %HV, %CV, %IC, σi2 and high R2). Upon these 10 genotypes, eight gave high yields (Express, DH11, DH14, DH15, DH21, DH30, DH39 and DH10). The DH26, DH65 and DH2 lines have given the best yields but are unstable. The majority of doubled haploid lines derived from Tichedrett×Express hybrid, showed more stability than the local parent and a yield equal to the general mean yield. Conclusion: Significant correlations were obtained within the two groups of parameters of static stability concept and dynamic stability concept, indicating the possibility of using a single stability parameter per group.
PDF Abstract XML References Citation
How to cite this article
D. Ramla, M.S. Yakhou, N. Bilek, M. Hamou, A. Hannachi, A. Aissat and L. Mekliche-Hanifi, 2016. Grain Yield Stability Analysis of Barley Doubled Haploid Lines in Algerian Semi-arid Zones. Asian Journal of Crop Science, 8: 43-51.
DOI: 10.3923/ajcs.2016.43.51
URL: https://scialert.net/abstract/?doi=ajcs.2016.43.51
DOI: 10.3923/ajcs.2016.43.51
URL: https://scialert.net/abstract/?doi=ajcs.2016.43.51
INTRODUCTION
In Algeria, barley (Hordeum vulgare L.) is the second rainfed conditions cultivated cereal after durum wheat (Triticum durum Desf.) with 1 million ha harvested areas. Its area production is mainly located in highland semi-arid agro-climatic zone (300-400 mm rainfall) characterized by variability and severity of climate conditions (irregular quantity and distribution of rainfall, spring frost, low winter temperatures and high temperatures of end cycle). These harsh climatic conditions have a negative impact on the level and stability of grain yields witch varied between 11.0 q ha–1 in dry years and 27 q ha–1 in rainy years1. For a long time, the national improvement efforts focused on grain yield as criterion of selection but because this strategy was conditioned by favorable and stable conditions2, limited results have been obtained. So, varieties were released but because their low potential of adaptation and their instability, they have not been adopted by the farmers and only the two varieties, Tichedrett and Saida, selected within local population, remain widely used and cover the major surfaces occupied by this specie3. Since then, the objective of barley breeders is to develop more and more varieties characterized by both stability and good yield level. The comportment unstable of varieties, which show yield fluctuations across the different environments, is due to the presence of Genotype× Environment Interactions (GEI)4. The importance of GEI in the process of improvement has been reported for a long time because that compromises the progress of the selection by making difficult the classification and the identification of superior varieties5. To mitigate this problem and exploit positively the GEI, stratification of the environments and multi environments trials are necessary to identify stable and high yielding genotypes6. Muhleisen et al.7 considered that selection for yield stability is not usually feasible due to the required number of test environments, which exceed the common capacity of barley breeding programs. However, this objective remains a central concern and many studies are conducted to investigate stability of barley genotype under different environments8-14. Several statistical methods were developed to analyze GEI. These analyses provide the ability of characterizing genotypes towards their adaptation and their degrees of stability. Among these methods, the parametric univariate stability statistics15 which were frequently used16 can be cited. The majority of reported work on this subject concern the promising genotypes derived from the final stages of the conventional plant breeding, few have concerned doubled haploids lines which are however considered as suitable materiel for GEI studies17.
The present study was carried out to assess the significance and magnitude of grain yield GEI, the stability of performance and the correlations among the stability parameters for 26 doubled haploid barley lines (DHs) and their parents under five semi-arid environments of Algeria.
MATERIALS AND METHODS
Crop material and field experiments: The crop material used in the experiment consisted in twenty-nine 6-row barley genotypes including 26 doubled haploid lines (DHs) and three parental varieties (local drought resistant and low yielding variety: The cv. Tichedrett and two commercial high yielding French introductions: The cvs. Express and Plaisant) (Table 1). This material was evaluated for grain yield at five semi-arid environments (locations×years) in 3 Algerian highlands barley growing areas during 3 years in 2011-2012 and 2013.
| Table 1: | Name and code of tested genotypes (parental varieties and doubled haploid lines) |
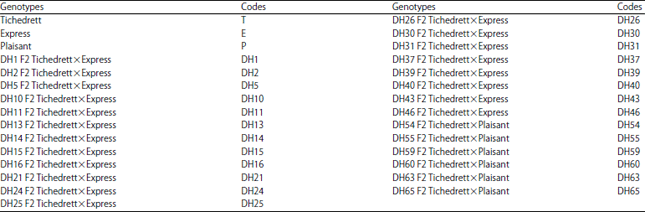 | |
The environments (locations×years) were principally differentiated by annual and seasonal rainfall variations (Table 2). The DHs lines were obtained in 2009 in the framework of INRAA’s barley breeding program by plant biotechnology tools. The DH lines were developed from F2 hybrids of 2 biparental single crosses between the local variety and the two introductions (F2 Tichedrett×Express, F2 Tichedrett×Plaisant) using anther culture procedure as described by Szarejko18,19, Cistue et al.20 and Jacquard et al.21. The field experiments were conducted at each location in a randomized complete block design with three replications. The experimental plots consisted of 5 m in length and 1.20 m in width (6 rows). Row to row distance was 20 cm. The seeding rate was 275 seeds m–2. The crop was harvested at maturity and the grain yield was obtained from 1 m section of 2 interior rows in the middle of each plot.
Data analysis: The variance analysis of data grain yield was realized by single environment to test the genotype effect and analyze the comportment and classify the genotypes using a Newman-Keuls’s test. A combined variance analysis of data of all environments was realized to test GE interaction effect and obtain variance components. The joint regression was used to analyze GE interaction for grain yield and approach the stability of genotypes. For this purpose, variance analysis of Finlay and Wilkinson22 was performed using "GEST" program, based on the model of Eberhart and Russell23 and developed by Ukai24. Each genotype was characterized by its regression coefficient (bi) and its variance of deviations from regression (S2di). A genotype with wide adaptation was defined as one with (bi = 1) and stable as one with (S2di = 0). The significance of regression slope (bi) from unity and deviation from regression (S2di) for each genotype were tested by t-test and F-test, respectively. The stability of genotypes was also approached through 2 static stability parameters such environmental variance25 (S2i) and coefficient of variation26 (%CV) in addition to six dynamic stability parameters consisted of Plaisted’s GE variance component27 (P), Wricke’s genotypic ecovalence28 (Wi), Shukla’s stability variance29 (σi2), Pinthu’s coefficient of determination30 (R2), heterogeneity variance (%HV) and incomplete correlation31 (%IC). Higher values for Pinthu’s coefficient (R2) and Plaisted’s stability parameter (P) indicate better genotypic stability. Lower values of remained parameters indicate higher stability. The relation magnitude between the used stability statistics was estimated from Pearson’s correlation coefficient.
RESULTS AND DISCUSSION
Analysis of grain yield performance of genotypes by environment: The variance analysis of the grain yield by environment showed a significant genotype effect for the environments E1-Khroub 2010-11, E2-Khroub 2011-12, E3-SBB 2011-12, E5-Setif 2012-13 (p<0.001) and E4-Setif 2011-12 (p<0.05) (Table 3). This effect indicated the existence of a usable genetic diversity for the selection. The average grain yield by environment varied from 20-39 q ha–1 (Table 4).
| Table 2: | Testing environments description |
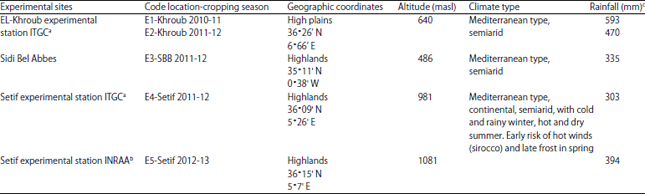 | |
| aTechnical Institute of Field Crops, bNational Institute of Agronomic Research of Algeria and cFrom September to June | |
| Table 3: | One way analysis of variance for grain yield in each of 5 environments tested |
 | |
| *,***Significant at p<0.05 and p<0.001, respectively | |
| Table 4: | Mean yield performancea (q ha–1) of 29 barley genotypes (26 DHs and 3 parents) field tested across 5 environments |
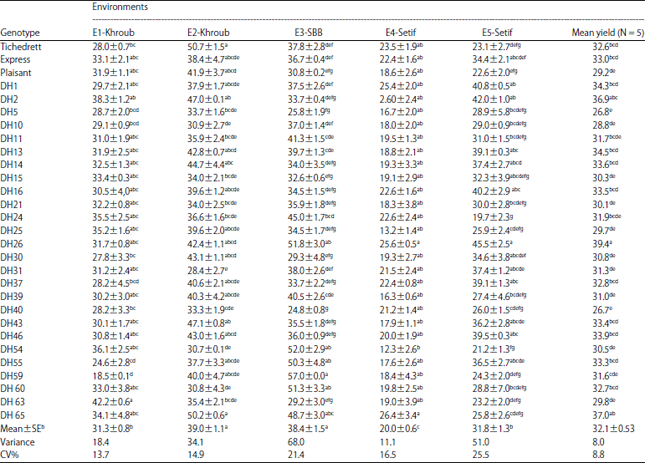 | |
| aData are given Mean±SE, bMean grain yield per trial (q ha–1), Values within the same column without the same letters indicate a significant difference according to Newman Keuls’s test at p<0.05 | |
The highest grain yield was obtained at E2-Khroub 2011-12 (39.4 q ha–1), followed by that obtained at E3-SBB 2011-12 (38.4 q ha–1), these two sites were the most favorable and occupied the first place (according to Newman-Keul’s test). The E5-Setif 2012-13 and E1-Khroub 2010-11 occupied the second place with respective yields of 31.8 and 31.3 q ha–1. The E4-Setif constituted the least favourable environment, registering the lowest average yield, 20.4 q ha–1 (Table 4). The average yields for genotypes (Table 4) varied from 12.3 q ha–1 (DH54 at E4-Setif site) to 57 q ha–1 (DH59 at E3-SBB 2011-12 site). Thus, at the E1-Khroub 2010-11 site, grain yield varied from 18.5 q ha–1, recorded by line DH59, to 42.2 q ha–1 for DH63 line. At E2-Khroub 2011-12, DH31recorded the lowest yield (28.4 q ha–1), while the parental cv. Tichedrett and DH65 line recorded the highest values respectively, 50.7 and 50.2 q ha–1. At E3-SBB 2011-12 site, DH40 line registered the lowest value performance (24.8 q ha–1) and DH59 line, the highest value (57q ha–1). At E4-Setif 2011-12, the DH65 line distinguished itself again and registered the most important value (26.4 q ha–1), the first place was also occupied by the DH26 line (25.6 q ha–1), which distinguished itself by second one better score (45.5 q ha–1) at the experimental site E5-Setif 2012-13. The lowest yields at the two latter sites were respectively registered by DH54 line (12.3 q ha–1) and DH24 line (19.7 q ha–1). The differential ranking of lines through the environments indicated the presence of a GE interaction32, which was confirmed by combined variance analysis (Table 5). The most important proportion of the variation (57.2%) had for source the environment, indicating contrasting environments. The part of the variation, which was due to genotype, represented only 9.4%, this weak proportion is certainly explained by the origin of the estimated lines. Indeed, all these lines have in common the same local parent Tichedrett, furthermore twenty of these lines were derived from a single cross (Tichedrett×Express) and the remaining lines from Tichedrett×Plaisant cross. The proportion of the variation explained by the GE interaction was high; it represented 33.4% of the total sum of square deviations (G+E+GEI) (Table 5). These proportions, it means, the largest part represented by the environment, followed by that of GE interaction and the lowest represented by the genotype, are in accordance with those reported by Bantayehu33 and Kadi et al.8. Besides that, the confirmation of the presence of the GE interaction required to conduct the analysis of grain yield stability to identify the stable and performing lines.
Analysis of grain yield stability and average performance of parental varieties and DHs lines: Variance analysis of Finlay and Wilkinson22 (Table 6) revealed that the joint regression was unsuitable to explain the GE interaction. In fact, heterogeneity of regression represented only 14.3% of the sum of squares of the GE interaction, while the deviation of the regression was explained most of the interaction (85.7%). More, heterogeneity of regression and pooled deviation from regression were respectively not significant and significant (p<0.001). Becker and Leon32 consider that only a small part of the GE interaction could be generally explained by the heterogeneity of regressions. It is because the majority of bi slopes have a value close to 1. This was confirmed in the present study, indeed 28 of the tested genotypes had a slopes bi =1 (Table 7), these genotypes were classified at wide adaptation. A single line, the DH 40, had a slope bi significantly lower than unity; it was characterized by a specific adaptation. According to the adaptability definition of Finlay and Wilkinson22, DH26, DH65 and DH2 lines, which had a slopes equal to the unity and a grain yield significantly superior to the general mean yield (μ) (Table 7), respectively equal to 39.4***, 37* and 36.9**, showed to be well adapted to the five tested environments. The DH5 line also had a slope equal to 1 but a grain yield significantly lower than (μ) 26.8**, this line was classified poorly adapted to all environments tested. Regarding the DH40 line whose slope was significantly less than 1 and a yield significantly inferior to (μ) 26.7***, was characterized by a specific adaptation to unfavorable environments. The parents varieties and the rest of DHs lines, had slopes equal to the unity and yield equal to (μ), they were classified as genotypes at wide adaptation and average stability. According to Eberhart and Russell23, who defined the stability of genotypes on the basis of two parameters (bi) and (S2di), the DH14, DH11, DH39, DH30, DH15, DH21, DH10 and DH5 lines and parental cv. Express, which had regression slopes equal to 1 and deviations from the regression S2di = 0 were defined as stable lines. The remaining lines had S2di significantly different from 0, were therefore based on this parameter and characterized by instability. The Pinthus’s (R2), varied from 25.6-84.8%. The most unstable 10 genotypes, with the lowest coefficients (25.6-51.1%) were, HD63, DH31, DH40, DH60, HD5, HD24, HD1, HD30, HD37 and Tichedrett, respectively. The first ten most stable lines, those with the highest coefficients (84.8-64.4%) were, DH13, DH11, DH25, DH43, DH39, DH10, DH46, DH21, Express and DH26, respectively.
| Table 5: | Combined analysis of variance for grain yield of 29 genotypes grown in five environments |
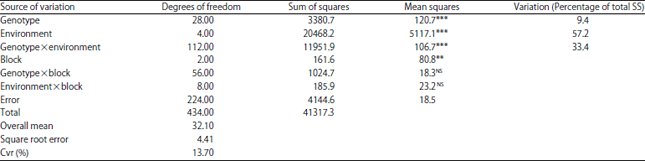 | |
| **,***Significant at p<0.01 and p<0.001, respectively, NS: Not significant at 5% level | |
| Table 6: | Finlay-Wilkinson variance analysis for grain yield stability over five environments |
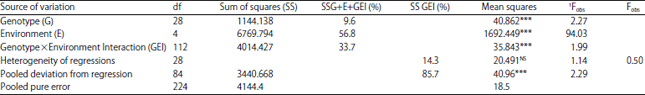 | |
1Calculated by comparing all mean squares to pooled pure error, 2Calculated by comparing pooled deviation from regression mean square to Pooled deviation from regression, ***Significant at p<0.001, NS: No significant | |
| Table 7: | Mean yield performance and stability parameters values of 29 barley genotypes (26 DHs and 3 parents) for grain yield |
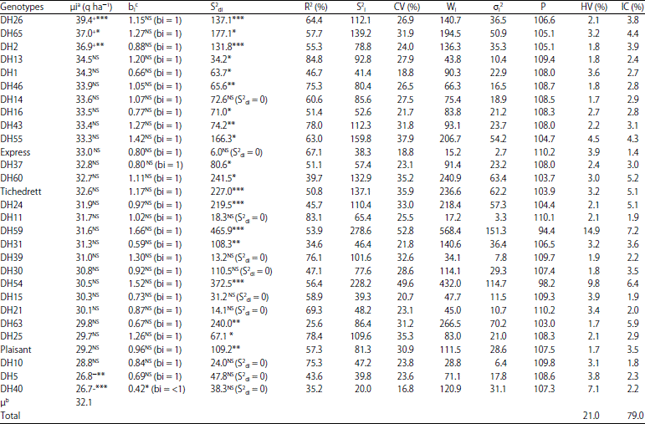 | |
aGrain yield of each genotype across all environments, bμ = General means yield of the 29 barley genotypes across 5 environments, +*, +**, +***Significantly superior to μ at p<0.05, p<0.01 and p<0.001, respectively, according to student test, NS: No significantly different to μ at p<0.05, according to student test, -*, -**, -***Significantly inferior to μ at p<0.05, p<0.01, p<0.001, respectively, according student test, bi: Regression coefficient, cAll bi are significantly different from zero based on linear regression analysis, bi was tested against "1" according to student test, at (0.05), S2di: Deviation of regression, S2di was tested against "0" according F-test (regression analysis), R2 (%): Coefficient of determination, S2i: Phenotypic variance, CV (%): Coefficient of variation (%), Wi: Wricke’s ecovalance, σi2: Shukla’s stability, P: Plaisted’s stability parameter, HV (%): Variance heterogeneity (%), IC (%): Incomplete correlation | |
The environmental variance specific at each genotype (S2i) varied from 20.0-278.6. According to this parameter, the 10 most stable genotypes, S2i from 20-57.4 were DH40, Express, DH15, DH5, DH31, DH10, DH21, DH16 and DH37, respectively and the ten more unstable, S2i from 109.6-278.6 were, DH25, DH24, HD26, DH43, HD60, Tichedrett, HD65, DH55, DH54 and DH59, respectively. The coefficient of variation (CV%) varied from 16.8-52.8, depending on this parameter, the more stable genotypes were DH40, DH1, Express, DH15, DH16 and the more unstable were DH59, DH54, DH55, Tichedrett, DH25 and DH60. Genotypic ecovalence (Wi) and stability of the variance of Shukla (σi2) parameters, varied from 15.2-568.4 and 2.7-151.3, respectively. These two parameters showed, that the most stable genotypes were respectively the introduced variety Express (Wi = 15.2 and σi2 = 2.7), the line DH11 (Wi = 17.2 and σi2 = 3.3), the line DH10 (Wi = 28.8 and σi2 = 6.4), the line DH39 (Wi = 34.1 and σi2 = 7.8), the line DH13 (Wi = 43.8 and σi2 = 10.4), while the most unstable genotypes were the local variety Tichedrett (Wi = 236.6 and σi2 = 62.2), the line DH60 (Wi = 240.9 and σi2 = 63.4), the line DH63 (Wi = 266.5 and σi2 = 70.2), the line DH54 (Wi = 432 and σi2 = 114.7) and the line DH59 (Wi = 568.4 and σi2 = 151.3). Plaisted parameter stability (P) varied from 110.2-94.4, thus identifying different levels of stability. The most stable level was characterized by Express (P = 110.2), DH21 (P = 110.2), DH11 (P = 110.1), followed by DH10 lines (P = 109.8), DH39 (P = 109.7), DH13 (P = 109.4) and DH15 (P = 109.3). The most unstable level was characterized by Tichedrett (P = 103.9), followed by DH60 lines (P = 103.7), DH63 (P = 103.0), DH54 (P = 98.2) and DH59 (P = 94.4), the most unstable line. Heterogeneity of variance (% HV) represented only 21.01% of the GE interaction (Table 7), its use in the assessment of the stability was not very effective, as reported by Grada and Ciulca34, contrary to the incomplete interaction (%IC) which explained 79.99% (Table 7) of the GE interaction. According to this last parameter, the cv. Express (%IC = 1.4%), the lines DH10 (%IC = 1.8%) and DH15 (% IC = 1.9%) were the most stable across the five environments, whereas the lines DH63 (5.9%), DH54 (6.4%) and DH59 (7.2%) were the most unstable (Table 7). The cv. Express, which was classified the most stable according to %IC, was among the most unstable according to the heterogeneity of variance (% HV = 3.9). The highest values of %HV (9.8 and 14.9%) were obtained respectively by DH54 and DH59, they were the most unstable. These lines were also the most unstable based on the genotypic ecovalence (Wi), Shukla stability variance (σi2), the incomplete correlation (%IC), Plaisted parameter (P), the %CV and the variance (S2i). If the majority of the parameters cited placed unequivocally these two lines in the ranks of the most unstable (Table 7), it was not the case for the ranking of the remaining lines, which showed some differences depending on the parameters. These results are in agreement with those of other authors who reported different rankings of lines according to the parameters33,13,16. It is important to indicate, for these tested environments that the introduced parent Express, although of the same level performance as the local parent Tichdrett, showed generally a good level of stability in contrast to the local parent and cv. Plaisant (Table 7). Moreover, DH5 line that derived from the cross between the local parent and the introduced cv. Express, showed a high stability and a grain yield performance below the general mean (μ) and both parents. This is in agreement with the conclusions of Bouzerzour et al.35 who reported that in unfavorable environments the most stable genotypes were less performing. The remaining lines derived from this crossing showed most of the time more stability than the parent Tichedrett (Table 7). These lines had a grain yield performance equal to the general mean (μ), except for DH26 and DH2 lines, which exhibited a grain yield superior to the general mean (μ) and to both parents. Even if, they were unstable, these two lines distinguish themselves from the rest and can be considered promising (Table 4 and 7). The lines that showed most instability were those derived from the cross between the local variety and cv. Plaisant (Table 7). This may be the result of the level of instability of both parents. The high instability of the DHs lines derived from this cross would be due to negative transgressions resulting from the association of alleles with negative effects contained in each parent36,37. The two lines from this crossing, which showed most instability, were DH59 and DH54. These lines were characterized by grain yields lower than the local parent (cv. Tichedrett) and of the same level as the introduced parent Plaisant, whose yield was among the lowest. Regarding the remaining DHs lines derived from this crossing, except for the DH65 line, which was characterized by a grain yield higher than the general mean yield (μ), both lines, DH63 and DH55 had grain yields equal to the general mean μ (Table 4).
Assessment of the level of correlations of the parameters of stability used: The majority of the correlations were significant (p<0.05-p<0.001) except for correlations with the general mean yield and between bi-%HV, R2-S2i, R2-CV%, R2-%HV. According to Sabaghnia et al.16 and El-Hashash and El-Absy13 no correlation between the average performance and the rest of the parameters is observed. Correlations between R2-S2di, R2-Wi, R2-σi2, R2-IC and the majority of the correlations with the parameter P were generally negative with the exception of correlations between P-R2 and R2-bi. These negative correlations, such as positive between P and R2 indicated a similar assessment of the level of stability of genotypes. While the positive correlation between R2-bi suggested an opposite assessment. The regression coefficient (bi) was significantly and positively correlated with S2di (r = 0.54***), R2 (r = 0.47**), S2i (r = 0.88***), CV% (r = 0.86***), Wi (r = 0.52***), σi2 (r = 0.52***) and %IC (r = 0.50***), a single negative correlation was obtained between this parameter and P (r = -0.51***). The deviation from the regression (S2di), was strongly and positively correlated with S2i (r = 0.87***), CV% (r = 0.83***), Wi (r = 0.98***), σi2 (r = 0.98***), %IC (r = 0.98***), %HV (r = 0.65***) and negatively correlated with R2 (r = -0.40**) and P (r = -0.98***). The coefficient of determination of Pinthu’s (R2) revealed significant and negative correlations with ecovalence Wi (r = -0.41**), Shukla’s stability of the variance σi2 (r = -0.41**), %IC (r = -0.45**) and positive with P (r = 0.41**). Phenotypic variance S2i was positively and significantly correlated to the coefficient of variation CV% (r = 0.96***), to the ecovalence Wi (r = 0.86***), to the stability of the variance of Shukla σi2 (r = 0.86***), to %HV (r = 0.63***), %IC (r = 0.82***) and negatively correlated with P (r = -0.85***). The coefficient of variation (CV%) was significantly and positively correlated with ecovalence Wi (r = 0.82***), stability of the variance of Shukla σi2 (r = 0.82***), %HV (r = 0.55**), %IC (r = 0.80***) and negatively correlated with P (r = -0.81***). The ecovalence Wi showed a complete correlation with the stability of variance of Shukla (1.00***), a positive correlations with %HV (r = 0.75***), %IC (r = 0.94***) and a strong negative correlation with P (r = -0.99***). The stability of the variance of Shukla (σi2) revealed identical correlations with %HV, %IC and Plaisted’s variance P like those revealed with genotypic ecovalence, r = 0.75***, r = 0.94*** and r = -0.99***, respectively. Finally Plaisted’s variance P showed a negative correlation with %HV (r = -0.73***), a strong and negative correlation with %IC (r = -0.95***) and %HV showed a positive correlation with %IC (r = 0.49***). Regarding the relations between the ecovalence (Wi) and Pinthu’s R2 (S2di) and the variance of Shukla’s (σi2), as well as between (S2i) and CV%, the present results are in accordance with those of Bantayehu33. The results of the present study, are also in agreement with those obtained by El-Hashash and El-Absy13 about the existence of correlations between stability variance of Shukla’s parameter σi2 and %CV, σi2 and bi and between %CV and S2di. The results are also in accordance with those of Sabaghnia et al.16 about the relationship between %CV and S2i, while they finds a perfect positive correlation between Wi and Plaisted’s P (r = 1.00). The parameters which were used in this study refer to two concepts of stability, static stability (bi, S2i and %CV) and dynamic stability (bi, R2, S2di, Wi, σi2, %HV and %IC). Relations between parameters within each group, as revealed by Becker38 and Becker and Leon32 were expected. This implies a similarity in the detection of stable genotypes by the parameters of each group (except for HV%) and therefore the possibility to simplify the analysis by the use of only one parameter per group39. The existence of correlations between parameters of the two different groups, also mean some similarities in the level of stability of genotypes determined by these parameters and may indicate the existence of DHs lines to static and dynamic stability as DH15, DH21, DH11 and DH10 lines. This may be due to the origin of these lines and the fact that they have identical genetic background.
CONCLUSION
The results indicated that barley doubled haploid population was suitable material for G×E interaction analysis. Grain yield performance was influenced by G×E interaction effects. This was resulted instability within the DHs population. Moreover, the results indicated that the DHs population contained desirable genotypes in terms of stability and performance. The DH14, DH11, DH39, DH30, DH15, DH21 and DH10, were identified as adapted and stable lines with a good average yield, DH26 and DH2 as the two best performing lines, adapted and more stable than the local parent. Therefore, all these doubled haploids lines can be considered as promising genotypes and can be recommended for the semi-arid environments tested. Significant correlations revealed that it could be sufficient to use only one parameter of each group to select genotypes of interest in a barley breeding program.
ACKNOWLEDGMENTS
This study was funded by the national fond of Algerian research of Ministry of Higher Education and Research. Thanks to M. Teffahi and A. Chikr for the installation of the trials (El-Khroub site) and the performance of measurements.
REFERENCES
- Powell, W., P.D.S. Caligari, M.S. Phillips and J.L. Jinks, 1986. The measurement and interpretation of genotype by environment interaction in spring barley (Hordeum vulgare). Heredity, 56: 255-262.
CrossRefDirect Link - Basford, K.E. and M. Cooper, 1998. Genotype x environment interactions and some considerations of their implications for wheat breeding in Australia. Aust. J. Agric. Res., 49: 153-174.
CrossRefDirect Link - Muhleisen, J., H.P. Piepho, H.P. Maurer, Y. Zhao and J.C. Reif, 2014. Exploitation of yield stability in barley. Theor. Applied Genet., 127: 1949-1962.
CrossRefDirect Link - Jalata, Z., A. Ayana and F. Fufa, 2011. Assessment of yield stability and disease responses in Ethiopian barley (Hordeium vulgare L.) landraces and crosses. Int. J. Agric. Res., 6: 754-768.
CrossRefDirect Link - Hanifi-Mekliche, L., A. Mekliche and P. Monneveux, 2011. Genotype x environment interaction and stability analysis on barley (Hordeum vulgare L.) lines in Algeria. Acta Agronomica Hungarica, 59: 325-336.
CrossRefDirect Link - Jalata, Z., 2012. Evaluation of ICARDA Barley genotypes for yield stability and lodging resistance in southeastern Ethiopia highlands. Electron. J. Plant Breed., 3: 722-732.
Direct Link - Mohammadi, R., B. Vaezi, A. Mehraban, H. Ghojigh, R. Mohammadi and N. Heidarpour, 2012. Analysis of multi-environment trials of rainfed barley in warm regions of Iran. J. Crop Improvement, 26: 503-519.
CrossRefDirect Link - El-Hashash, E.F. and K.M. El-Absy, 2013. Genotype X environment interaction, environmental indices and stability analyses for some selected genotypes of Barley. J. Applied Sci. Res., 9: 4371-4377.
Direct Link - Flores, F., M.T. Moreno and J.I. Cubero, 1998. A comparison of univariate and multivariate methods to analyze GxE interaction. Field Crops Res., 56: 271-286.
CrossRefDirect Link - Sabaghnia, N., M. Mohammadi and R. Karimizadeh, 2013. Yield stability of performance in multi-environment trials of barley (Hordeum vulgare L.) genotypes. Acta Univ. Agric. Silvic. Mendelianae Brun., 61: 787-793.
CrossRefDirect Link - Szarejko, I., 2001. In vitro and Rogenesis for Doubled Haploid Production in Barley (Hordeum vulgare L.) and Wheat (Triticumae stivum L). In: Manual on General Genetics and Basic Methods in Plant Biotechnology, Szarejko, I. and R.N. Jones (Eds.). Wydawnictwo Uniwersytetu Slaskeigo, Katowice, Poland, ISBN: 9788322610855, pp: 159-173.
- Cistue, L., M.P. Valles, B. Echavarri, J.M. Sanz and A. Castillo, 2003. Barley Anther Culture. In: Doubled Haploid Production in Crop Plants: A Manual, Maluszynski, M., K.J. Kasha, B.P. Forster and I. Szarejko (Eds.). Springer Science and Business Media, New York, USA., ISBN-13: 9781402015441, pp: 29-34.
- Jacquard, C., R. Asakaviciute, A.M. Hamalian, R.S. Sangwan, P. Devaux and C. Clement, 2006. Barley anther culture: Effects of annual cycle and spike position on microspore embryogenesis and albinism. Plant Cell Rep., 25: 375-381.
CrossRefPubMedDirect Link - Finlay, K.W. and G.N. Wilkinson, 1963. The analysis of adaptation in a plant-breeding programme. Aust. J. Agric. Res., 14: 742-754.
CrossRefDirect Link - Eberhart, S.A. and W.A. Russell, 1966. Stability parameters for comparing varieties. Crop Sci., 6: 36-40.
CrossRefDirect Link - Francis, T.R. and L.W. Kannenberg, 1978. Yield stability studies in short-season maize. I. A descriptive method for grouping genotypes. Can. J. Plant Sci., 58: 1029-1034.
CrossRefDirect Link - Plaisted, R.L., 1960. A shorter method for evaluating the ability of selections to yield consistently over locations. Am J. Potato Res., 37: 166-172.
CrossRefDirect Link - Shukla, G.K., 1972. Some statistical aspects of partitioning genotype-environmental components of variability. Heredity, 29: 237-245.
PubMedDirect Link - Muir, W., W.E. Nyquist and S. Xu, 1992. Alternative partitioning of the genotype-by-environment interaction. Theor. Applied Genet., 84: 193-200.
CrossRefDirect Link - Becker, H.C. and J. Leon, 1988. Stability analysis in plant breeding. Plant Breed, 101: 1-23.
CrossRef - Bantayehu, M., 2009. Analysis and correlation of stability parameters in malting barley. Afr. Crop Sci. J., 17: 145-153.
Direct Link - Grada, F. and S. Ciulca, 2013. Analysis of genotype x environment interaction for yield in some maize hybrids. J. Hortic. For. Biotechnol., 17: 192-196.
Direct Link - Vega, U. and K.J. Frey, 1980. Transgressive segregation in inter and intraspecific crosses of barley. Euphytica, 29: 585-594.
CrossRefDirect Link - Rieseberg, L.H., A. Widmer, A.M. Arntz and J.M. Burke, 2003. The genetic architecture necessary for transgressive segregation is common in both natural and domesticated populations. Phil. Trans. R. Soc. Lond. B, 358: 1141-1147.
CrossRefDirect Link - Mekliche, A., F. Dahlia and L. Hanifi-Mekliche, 2013. Agro-morphological diversity and stability of durum wheat lines (Triticum durum Desf.) in Algeria. Acta Agronomica Hungarica, 61: 149-159.
CrossRefDirect Link