
Vaibhav Kumar Maurya
School of Bio Sciences and Technology, VIT University, Vellore, 632014, Tamil Nadu, India
R. Srinvasan
School of Bio Sciences and Technology, VIT University, Vellore, 632014, Tamil Nadu, India
N. Ramesh
School of Bio Sciences and Technology, VIT University, Vellore, 632014, Tamil Nadu, India
M. Anbalagan
School of Bio Sciences and Technology, VIT University, Vellore, 632014, Tamil Nadu, India
K.M. Gothandam
School of Bio Sciences and Technology, VIT University, Vellore, 632014, Tamil Nadu, India
Asian Journal of Crop Science
Year: 2015 | Volume: 7 | Issue: 4 | Page No.: 286-294







ABSTRACT
Carotenoids pathway is one of decisive anabolic pathway which is responsible for diverse, vital functions such as absorption, dissipation and transfer of solar energy for photosynthesis. Change in environmental factors causes modulation in molecular as well physiological responses to acclimate altered conditions. The current study is designed to assess the difference in the expression pattern of carotenoid pathway genes as well as accumulation of β-carotene among three capsicum varieties having different sensitivity level against salt. To achieve this objective salt tolerant (G4), moderate, tolerant (K2) and sensitive (CO1) varieties were treated under different NaCl concentration (25, 50, 100, 150 and 200 mM) for 30 days. Differential expressions of five major genes of carotenoid pathway (phytoene synthase, phytoene desaturase, zeta carotene desaturase, lycopene β-cyclase and capsanthin/capsorubin synthase) were examined. Semi-quantitative real time PCR expression data have demonstrated the declined expression of genes under study as the salt concentration gradually increased. We also analyzed the impact of salt on chlorophyll content and β-carotene content.
PDF Abstract XML References Citation
Received: August 18, 2015;
Accepted: September 28, 2015;
Published: October 08, 2015
How to cite this article
Vaibhav Kumar Maurya, R. Srinvasan, N. Ramesh, M. Anbalagan and K.M. Gothandam, 2015. Expression of Carotenoid Pathway Genes in Three Capsicum Varieties under Salt Stress. Asian Journal of Crop Science, 7: 286-294.
DOI: 10.3923/ajcs.2015.286.294
URL: https://scialert.net/abstract/?doi=ajcs.2015.286.294
DOI: 10.3923/ajcs.2015.286.294
URL: https://scialert.net/abstract/?doi=ajcs.2015.286.294
INTRODUCTION
Capsicum is one of the most demanded spices due to its nutritional and flavor attributes such as carotenoids and punginess (Maga and Todd, 1975). It is consumed in various forms such as dried, fresh and food additives (Govindarajan and Salzer, 1986). The genetic potential of plants always has been at stake due to biotic and abiotic stresses. These stresses not only hamper proper plant growth and productivity but also limit the availability of land (Cramer et al., 2008). Abiotic stresses such as salinity, drought, high temperature, cold and freezing can be potential threats to plants by reducing their potential yield and growth to that level which may lead to death (Rubio et al., 2009). High salinity hampers the cellular and molecular functions by imparting osmotic and ionic stress to individual cell as well as to whole plant (Zhu, 2002; Golldack et al., 2003). Ionic stress agitates the mineral distribution, membrane instability and turgor pressure while osmotic stress confines the water available to plant (Hsiao, 1973; Shahid et al., 2011). High Na+ accumulation mainly prevents the uptake of many fundamental nutrients due to competitive interactions and ion selection of membranes (Alam, 1999). This affects the photosynthetic electron transport extensively, thereby hinders the photosynthesis efficiency (Sudhir et al., 2005). The most of crop plants are salt sensitive and show a range of visual symptoms during vegetative growth (leaf chlorosis, necrosis and abscission, reduced growth rate, plant death) (Bethke and Drew, 1992).
During fruit development plastids differentiate into chromoplasts that are meticulous for the sequestration of lipophilic molecules such as carotenoids (Camara et al., 1995). Previous studies report the induction of carotenoid synthesis pathways such as phytoene synthase and put on during fruit development (Giuliano et al., 1993), the mRNAs encoding cyclization enzymes are downregulated (Ronen et al., 1999), which lead into an accumulation of carotenoids in fruit. Many pathways have been reported which undergo modulation under changed environment (Zhu, 2002). The carotenoid biosynthetic pathway is one of the vital pathways which are essential for plant productivity. The regulatory mechanisms that control carotenoid biosynthesis in plants are scantily spoken and the activities of carotenogenic enzymes in the plastics have neither been correlated with the amounts and types of carotenoids in the capsicum especially under saline conditions. In response to altered environmental condition these genes undergo transcriptional control which may play key role in the regulation of carotenogenesis in higher plants (Cunningham Jr and Gantt, 1998; Simkin et al., 2003; Campisi et al., 2006).
Scanty information is available about the molecular mechanism which is responsible of modulation in transcript expression of carotenoid pathways genes under salt stress condition. Therefore, differential study of carotenoid gene expression in capsicum varieties with various level of salt sensitivity may lead to better understanding about transcript regulation under salt stress. For this purpose we selected five major carotenoid genes phyotene synthetase (PSY), phytoene desaturase (PDS), zetacarotene desaturase (ZCD), lycopene β-cyclase (LCY-β) and Capsanthin/Capsorubin Synthase (CCS). This study aims to advance a better apprehension of the regulation of carotenoid biosynthesis in Capsicum annuum at the molecular levels under salt stress condition for this function, we have selected three capsicum salt tolerant (G4), moderate tolerant (K2) and salt sensitive (CO1) with various salt sensitivity levels. In this report, capsicum seedlings were used to address mainly 4 questions
| • | Are the carotenoid biosynthetic genes co-expressed in the leaves as they reported in fruit? |
| • | How salt treatments do affect the gene expression of carotenoid biosynthetic gene (PSY, PDS, ZDS, LCY-β and CCS)? |
| • | How does salt affect the chlorophyll content of capsicum cultivar? |
| • | Does salt affect the accumulation g β-carotene among selected varieties? |
MATERIALS AND METHODS
Plant material and salt treatment: Seeds of three capsicum cv. susceptible cultivar (CO1), tolerant (G4) and moderately sensitive (K2) (Lal et al., 1990) were collected from Coimbatore Agriculture University and allowed to grow in soil pots under controlled green house conditions at VIT University, Vellore. Ten plants were placed in six sets of plants (three replicates) for each cultivar and allowed to growth for 60 days. Then plants were treated with 500 mL solution of six different concentration of NaCl such as 25 mM (2.5 dS/m), 50 mM (4.95 dS/m), 100 mM (9.8d S/m), 150 mM (14.6 dS/m) and 200 mM (19.6 dS/m), respectively for 30 days. Control plants were treated with distilled water. This study is laboratory based and each experiment performed in this study was conducted in triplicates.
Chlorophyll estimation: Spectrophotometric determination of chlorophyll A and B from the plant tissue samples was carried out as described previously by Fray et al. (1995). The tissue samples were homogenized to fine powder in liquid nitrogen with a mortar and pestle. A water miscible organic solvent methanol was added to 50 mg of the sample. It was incubated for 20 min on ice to extract the carotenoid pigments. Any substantial amount of water present was removed by partitioning with 10% diethyl ether in petroleum ether (boiling point 40-60°C). The suspension was mixed by inversion for 1 h in dark. The aqueous phase was removed and distilled water was added to the organic phase to further facilitate the partition by removing any residual methanol. Following centrifugation for 3 min 9600×g, the epiphase had been discarded and the hypophase was reextracted with petroleum ether (boiling point 40-60°C). The organic extracts obtained were pooled, dried under vaccum and redissolved in 1 mL of diethyl ether.
| • | Chlorophyll a (Chla) (µg mL‾1) = 9.93 A660.0-0.75 A641.8 |
| • | Chlorophyll b (Chlb) (µg mL‾1) = 16.23 A641.8-2.42 A660.0 |
Quantitative analysis of β-carotene: The β-carotene was extracted from 0.25 g capsicum fresh fruit material using methanol and 10% diethyl ether in petroleum ether organic solvent and high performance liquid chromatography was employed to separate and quantify the β-carotenoid as detailed previously in study of Fraser et al. (1994). The chromatographic separation of extracted carotenoids was performed on a reverse phase C18 column maintained at 25°C in an isocratic elution mode. The mobile phase comprising acetonitrile: methanol: isopropanol (85:10:5) was maintained at a flow rate of 1 mL min‾1. The carotenoids were monitored using UV detector set at a wavelength of 470 nm. Carotenoids were identified by their characteristic spectral properties, relative mobilities and co-chromatography with authentic standards. Amount of individual carotenoids were determined by integration of the peak areas.
RNA extraction: All glassware plastic wares, mortar and pestle used in RNA isolation were treated with 0.1% (v/v) DEPC for 24 h followed two times autoclaving and oven drying at 80°C for 48 h. A 50 μg of leaves sample was homogenized with liquid nitrogen, resulting powder was mixed in 1 mL of trizol reagent (invitogen) and incubated at room temperature for 15 min at room temperature followed by addition of chloroform. Mixture was centrifuged at 12000×g for 15 min at 4°C and supernatant was precipitated at -20°C for 30 min after adding isopropanol. The air dried pellets were dissolved in RNAse free water and equal volume of saturated phenol was mixed. The mixture was centrifuged at 12000×g for 10 min. Resulting supernatant was taken in new tube and equal volume of chloroform was added followed by centrifugation for 10 min at 12000×g. Supernatant was taken out in new tube followed by addition of 2.5 volume of 70% ethanol 1/10 volume of 3 M sodium acetate and it was incubated at -20°C for overnight for precipitation. Then mixture was centrifuged at 13000×g for 25 min and pellet were dried and dissolved in RNAse free molecular grade water. The quality of total RNA was checked by spectrophotometric absorbance at 260 and 280 nm.
First strand c-DNA synthesis: The c-DNA was synthesized using RNA three replicate isolated from three set of salt treated as well as control plant using oligo (dT) primer (900 ng). To the RT-PCR reaction, 2 μL dNTPS (10 mM), reverse transcriptase 1 μL, 5X assay buffer and 2 μL DTT, 1 μL RNAse inhibitor and 6 μL RNA (200 ng) and make up to 20 μL with DEPC treated molecular grade water. The reaction was carried out at 37°C for 60 min and 72°C for 15 min in eppendorf thermo cycler. The cDNA was tested for amplification using β-actin specific primer for 30 cycles.
Primer designing: Gene specific primers (Table 1) were designed manually based on the sequence information available at NCBI GenBank database (http://www.ncbi.nlm.nih.gov) and thermodynamic properties were analyzed with the help of Vector NTI software.
| Table 1: | Primers used in analysis of carotenoid biosynthetic pathway genes |
 | |
Primer optimization and PCR normalization: The gene specific primers were used to carry out transcript expression analysis using PCR. The reaction was made up using 2X buffer, 200 μM dNTPS, reverse primer 1 μL, forward primer 1 μL DNA polymerase 2 μg, make up the volume up to 20 μL with RNAse free molecular water. The thermal cycling included initial denaturation (95°C, 5 min), followed by 30 cycles of denaturation (95°C, 30 sec), primer annealing (65°C, 30 sec) and primer extension (72°C, 1 min) followed by final extension 72°C for 15 min. For ascertaining equal RNA loading in PCR reaction, actin was used as an internal control. For each gene transcript analysis three replicates of c-DNA from three set of treated plant were used.
Statistical analysis: All the experiment was performed in triplicate. The analysis of variance (Two-way ANOVA) between the control and sodium bicarbonate supplemented groups was carried out using Tukey’s multiple comparisons test at p<0.05 in Graph pad Prism (Version 6.0). Figures represent in mean and their standard deviation.
RESULTS
Level of Chlorophyll in treated and untreated plants of three capsicum cultivars: Major changes were observed in pigments as the plants were treated with increase salt concentration. Chlorophyll A and B were maximal in untreated controls. Figure 1a clearly demonstrates rapid decline chlorophyll A e.g., 0.546141 (CO1), 0.382886 (K2) and 0.593712 (G4) as salt treatment varied from 0-200 mM. Increase in salt concentration was accompanied by rapid decrease in Chlorophyll B content e.g., 0.332282 (CO1), 0.454354 (K2) and 0.372658 (G4) for Fig. 1b.
Level of β-carotene content in treated and untreated plants of three capsicum cultivars: The amount β-carotene in three capsicum cultivars were determined by HPLC analysis as illustrated in Fig. 2, where characteristic pattern of decrease in β-carotene can be observed as the salt concentration increased from 0-200 mM. A significant decrease in β-carotene levels by 0.488234 (CO1), 0.496517 (K2) and 0.583724 (G4) folds were seen in fruits of selected cultivars when compared to control plant fruits (Fig. 2).
Expression level of carotenoid genes in the treated and untreated plants of three capsicum cultivars: The PSY expression decline in all plants exposed to salt differed significantly among the genotypes and significantly decreased in plants exposed to salt. However, the highest expression level was observed in salt tolerant cultivar (G4) followed by moderate sensitive cultivar (K2) while it was least expressed in salt susceptible cultivar (CO1). The PDS expression level reduced in all plants exposed to salt differed among the genotypes and significantly decreased in plants exposed to salt as compare to control plants. Though higher expression level was detected tolerant cultivar (G4) than moderate sensitive (K2) susceptible cultivar (CO1) but between G4 and K2 cultivar transcript expression was not found significantly varied.
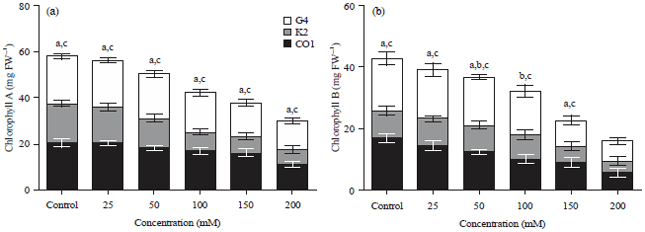 | |
| Fig. 1(a-b): | (a) Chlorophyll A and (b) Chlorophyll B content of Capsicum cultivars CO1, K2 and G4 were estimated in the plants treated with sodium chloride and control, (1) 0 mM, (2) 50 mM, (3) 100 mM, (4) 150 mM and (5) 200 mM of sodium carbonate supplemented cultures values are expressed in Mean±SD (n = 3), a, b, c significant value between capsicum cultivars CO1×K2, CO1×G4 and K2×G4 (p<0.05) |
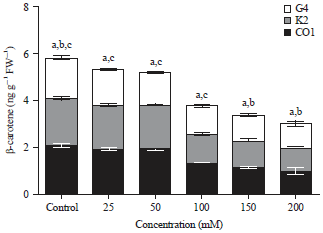 | |
| Fig. 2: | β-carotene content of Capsicum cultivars CO1, K2 and G4 were estimated in the plants treated with sodium chloride and control, (1) 0 mM, (2) 50 mM, (3) 100 mM, (4) 150 mM and (5) 200 mM of sodium carbonate supplemented cultures Values are expressed in Mean±SD (n = 3). a, b, c significant value between capsicum cultivars CO1×K2, CO1×G4 and K2×G4 (p<0.05) |
The gene transcripts expression levels of ZDS decreased significantly in tolerant cultivar (G4) and moderate sensitive cultivar (K2) as the plants were exposed to salt. While among the selected cultivars more reduction in transcript expression was observed in salt susceptible cultivar than moderate sensitive cultivar and tolerant cultivar as the salt concentration increased (Fig. 3). The LYC-β expression declined in all plants exposed to salt differed significantly among the genotypes and significantly decreased in plants exposed to salt. The extent of decrease was significantly more in salt susceptible cultivar (CO1) as compared to moderate sensitive (K2) cultivar while LYC-β was least expressed in leaves of salt tolerant cultivar (G4). The CCS expression level reduced in all plants exposed to salt differed significantly among the genotypes and significantly decreased in plants exposed to salt. However, the decrease in expression level was least in G4 cultivar followed by moderately sensitive cultivar (K2). But in case of salt susceptible cultivar (CO1) it was least in 200 mM treated plant among applied salt concentrations (Fig. 3).
DISCUSSION
Owing to the health beneficial potential and economic importance of carotenoids, carotenoid biosynthetic pathway remains as a topic of scrutinized research since many years. Earlier reports on the expression pattern of carotenoid pathway genes have been targeted on flower and fruit part but very scanty information is available in leaves (Plaut et al., 1987; Babu et al., 2011). Our data clearly show that a transcript of selected genes (PSY, PDS, ZDS, LYC-β and CCS) were detectable in leaf samples of all capsicum varieties (Romer et al., 1993). As real time data clearly establish that a tendency of decline in gene expression of selected genes (PSY, PDS, ZDS, LCY-β and CCS) were celebrated as the plants were exposed to different salt concentration.
Earlier studies on carotenoid genes have already reported how abiotic stress, such as temperature, salt, drought and hormones influences the gene expression. A study in maize under thermal stress caused a decline in expression level of phytoene synthase gene. Previous reports reveal that gene expression of carotenoid pathways in regulated by various factors such as transcription factor (RAP2) (Welsch et al., 2007), phytochrome interacting factors (Toledo-Ortiz et al., 2010) and phytohormones (ethylene and ABA) (Sandmann et al., 2006; Welsch et al., 2008; Lee et al., 2012). The PSY and PDS have motif (ATCTA) region which is binding region for RAP2 transcription factors. Generally RAP2 represses the expression of these carotenoid genes by binding to cis-acting motif (ATCTA). Some of previous study also reported that transcription factor exhibit enhancement under abiotic stress such as drought and salinity (Castillon et al., 2007). In addition of RAP2 transcriptional regulation phytochrome interacting factors also play crucial role in regulating the carotenoid pathways genes expression. High level of PIPs generally Phytochrome Interacting Factor (PIF) family generally down-regulate the accumulation of carotenoids by specifically suppressing the gene encoding phytoene synthase (Yaduvanshi, 2009), the rate-determining enzyme of the pathway. Microarray data of Arabidopsis reveals that PIPs generally induced under stress condition such drought and salinity.
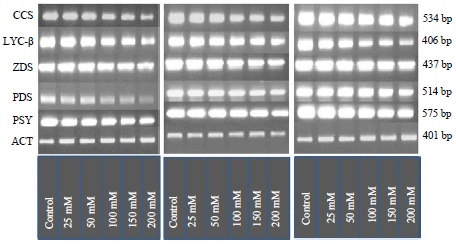 | |
| Fig. 3: | Gene expression of carotenoid pathway genes of three capsicum variety under salt stress |
When plants were exposed different salt concentration we did notice decline in expression of carotenoid pathway genes but maximum decline was observed in case of salt susceptible variety followed by moderate sensitive while salt tolerance variety was able to compensate the deleterious effects of salt. This could be explained by our previous study on difference in tolerance level among the selected varieties. Present study also advocates the previous reports which demonstrate the decline of carotenoid and chlorophyll pigments under abiotic stress (Babu et al., 2011). Decline in accumulation of β-carotene and individual chlorophylls in salt treated plant of capsicum cultivars can be justified by decrease in expression levels of carotenoid pathways genes under salt stress.
When plants were subjected to salt, a trend of decline in β-carotene level was noticed. This may be either due to degradation of carotenoid under salt stress condition or due to decreased gene expression of carotenoid pathway genes. To develop salt tolerant the plants, a proper knowledge about molecular mechanisms and factor involved in proper development and maturation of the plant is indispensable. Since carotenoids play critical role in harvesting solar energy as well as in protecting the photo system. Thus form essential components of photosynthetic antenna and reaction center complex, these vital functions make carotenoid of the essence in plant development. Our major interest was to analyze the expression pattern in leaves of three capsicum plants as well as to observe the difference in expression level between the selected varieties which are having different sensitivity level against salt. This work clearly emphasizes that capsicum plants under salt stress shows a pattern of decline in expression of carotenoid biosynthetic genes in leaves and also the gene extent of decrease varied from single form to another depending upon the tolerance point.
REFERENCES
- Babu, M.A., D. Singh and K.M. Gothandam, 2011. Effect of salt stress on expression of carotenoid pathway genes in Tomato. J. Stress Physiol. Biochem., 7: 87-94.
Direct Link - Bethke, P.C. and M.C. Drew, 1992. Stomatal and nonstomatal components to inhibition of photosynthesis in leaves of Capsicum annuum during progressive exposure to NaCl salinity. Plant Physiol., 99: 219-226.
Direct Link - Camara, B., P. Hugueney, F. Bouvier, M. Kuntz and R. Moneger, 1995. Biochemistry and molecular biology of chromoplast development. Int. Rev. Cytol., 163: 175-247.
CrossRefDirect Link - Campisi, L., M. Fambrini, V. Michelotti, M. Salvini, D. Giuntini and C. Pugliesi, 2006. Phytoene accumulation in sunflower decreases the transcript levels of the phytoene synthase gene. Plant Growth Regul., 48: 79-87.
CrossRefDirect Link - Castillon, A., H. Shen and E. Huq, 2007. Phytochrome interacting factors: Central players in phytochrome-mediated light signaling networks. Trends Plant Sci., 12: 514-521.
CrossRefDirect Link - Cramer, V.A., R.J. Hobbs and R.J. Standish, 2008. What's new about old fields? Land abandonment and ecosystem assembly. Trends Ecol. Evol., 23: 104-112.
CrossRefDirect Link - Cunningham, Jr. F.X. and E. Gantt, 1998. Genes and enzymes of carotenoid biosynthesis in plants. Annu. Rev. Plant Biol., 49: 557-583.
CrossRefDirect Link - Fraser, P.D., M.R. Truesdale, C.R. Bird, W. Schuch and P.M. Bramley, 1994. Carotenoid biosynthesis during tomato fruit development (evidence for tissue-specific gene expression). Plant Physiol., 105: 405-413.
Direct Link - Fray, R.G., A. Wallace, P.D. Fraser, D. Valero, P. Hedden, P.M. Bramley and D. Grierson, 1995. Constitutive expression of a fruit phytoene synthase gene in transgenic tomatoes causes dwarfism by redirecting metabolites from the gibberellin pathway. Plant J., 8: 693-701.
CrossRefDirect Link - Giuliano, G., G.E. Bartley and P.A. Scolnik, 1993. Regulation of carotenoid biosynthesis during tomato development. Plant Cell, 5: 379-387.
Direct Link - Golldack, D., F. Quigley, C.B. Michalowski, U.R. Kamasani and H.J. Bohnert, 2003. Salinity stress-tolerant and -sensitive rice (Oryza sativa L.) regulate AKT1-type potassium channel transcripts differently. Plant Mol. Biol., 51: 71-81.
CrossRefDirect Link - Govindarajan, V.S. and U.J. Salzer, 1986. Capsicum-production, technology, chemistry and quality-part II. Processed products, standards, world production and trade. Crit. Rev. Food Sci. Nutr., 23: 207-288.
CrossRefPubMedDirect Link - Hsiao, T.C., 1973. Plant responses to water stress. Annu. Rev. Plant Physiol., 24: 519-570.
CrossRefDirect Link - Lal, S., J. Mangal and P. Hooda, 1990. Salt tolerance of six varieties of chilli. Haryana Agric. Univ. J. Res., 20: 49-53.
Direct Link - Lee, J.M., J.G. Joung, R. McQuinn, M.Y. Chung and Z. Fei et al., 2012. Combined transcriptome, genetic diversity and metabolite profiling in tomato fruit reveals that the ethylene response factor SlERF6 plays an important role in ripening and carotenoid accumulation. Plant J., 70: 191-204.
CrossRefDirect Link - Maga, J.A. and P.H. Todd, 1975. Capsicum. Crit. Rev. Food Sci. Nutr., 6: 177-199.
CrossRefDirect Link - Plaut, Z., M.L. Mayoral and L. Reinhold, 1987. Effect of altered sink: Source ratio on photosynthetic metabolism of source leaves. Plant Physiol., 85: 786-791.
Direct Link - Romer, S., P. Hugueney, F. Bouvier, B. Camara and M. Kuntz, 1993. Expression of the genes encoding the early carotenoid biosynthetic-enzymes in Capsicum annuum. Biochem. Biophys. Res. Commun., 196: 1414-1421.
CrossRefDirect Link - Ronen, G., M. Cohen, D. Zamir and J. Hirschberg, 1999. Regulation of carotenoid biosynthesis during tomato fruit development: Expression of the gene for lycopene epsilon-cyclase is down-regulated during ripening and is elevated in the mutant Delta. Plant J., 17: 341-351.
CrossRefPubMedDirect Link - Rubio, J.S., F. Garcia-Sanchez, F. Rubio and V. Martinez, 2009. Yield, blossom-end rot incidence and fruit quality in pepper plants under moderate salinity are affected by K+ and Ca2+ fertilization. Scientia Horticulturae, 119: 79-87.
CrossRefDirect Link - Sandmann, G., S. Romer and P.D. Fraser, 2006. Understanding carotenoid metabolism as a necessity for genetic engineering of crop plants. Metab. Eng., 8: 291-302.
CrossRefDirect Link - Shahid, M.A., M.A. Pervez, R.M. Balal, R. Ahmad, C.M. Ayyub, T. Abbas and N. Akhtar, 2011. Salt stress effects on some morphological and physiological characteristics of okra (Abelmoschus esculentus L.). Soil Environ., 30: 66-73.
Direct Link - Simkin, A.J., C. Zhu, M. Kuntz and G. Sandmann, 2003. Light-dark regulation of carotenoid biosynthesis in pepper (Capsicum annuum) leaves. J. Plant Physiol., 160: 439-443.
CrossRefDirect Link - Sudhir, P.R., D. Pogoryelov, L. Kovacs, G. Garab and S.D.S. Murthy, 2005. The effects of salt stress on photosynthetic electron transport and thylakoid membrane proteins in the cyanobacterium Spirulina platensis. J. Biochem. Mol Biol., 38: 481-485.
CrossRefDirect Link - Toledo-Ortiz, G., E. Huq and M. Rodriguez-Concepcion, 2010. Direct regulation of phytoene synthase gene expression and carotenoid biosynthesis by phytochrome-interacting factors. Proc. Natl. Acad. Sci. USA., 107: 11626-11631.
CrossRefDirect Link - Welsch, R., D. Maass, T. Voegel, D. DellaPenna and P. Beyer, 2007. Transcription factor RAP2.2 and its interacting partner SINAT2: Stable elements in the carotenogenesis of Arabidopsis leaves. Plant Physiol., 145: 1073-1085.
Direct Link - Welsch, R., F. Wust, C. Bar, S. Al-Babili and P. Beyer, 2008. A third phytoene synthase is devoted to abiotic stress-induced abscisic acid formation in rice and defines functional diversification of phytoene synthase genes. Plant Physiol., 147: 367-380.
Direct Link - Zhu, J.K., 2002. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol., 53: 247-273.
CrossRefPubMedDirect Link