
M. Syafii
Faculty of Agriculture, Universitas Singa Perbangsa, Karawang, Indonesia
I. Cartika
Faculty of Agriculture, Universitas Majalengka, Indonesia
D. Ruswandi
Padjadjaran University, Bandung, 40600, Indonesia
Asian Journal of Crop Science
Year: 2015 | Volume: 7 | Issue: 4 | Page No.: 244-255







ABSTRACT
Maize is one of the most important cereals in the world after wheat and rice. Now-a-days, the improvement of maize production is hindered by conversion of agricultural lands into industrial and residential areas. One of the suggested solutions is the production of maize cultivars that can be incorporated in agroforestry systems and can be grown with tree plants such as Albizia. The success of the improvement of new maize cultivars suited to this Maize-Albizia system depends on the availability of genetic variability. Research on the genetic diversity of maize inbred lines that is suited to the agroforestry system with Albizia were conducted in a real forests planted with Albizia tree plants that are around three years old in Cimalaka, Sumedang, West Java, Indonesia. The evaluation of inbred lines was laid on a split plot design with two replications. The main plot consisted of two cropping systems, namely; maize sole cropping system and Maize-Albizia agroforestry system. The subplots were having seventy five inbred lines of maize. The biometric characters included days to anthesis, days to silking, days to harvesting, plant height, length of nodes, plant diameter, chlorophyll content, leaf area index, ear length, ear diameter, number of rows, weight of 1000 seeds and ear weight per plant. Based on the analysis of the main components, in conditions maize sole cropping system, the results showed an Eigen value between 1.08-4.52 which contributes to 67.27% variability, whereas, in the condition Maize-Albizia cropping system, the Eigen value ranged from 1.16-5.37 which contributed to 71.85% variability. Cluster analysis of 75 maize genotypes showed a wide distribution obtaining nine clusters in the maize sole cropping system and five clusters in the Maize-Albizia cropping system. High genetic variability seen in both cropping system greatly supports the development of elite new cultivars of maize that can be utilized in agroforestry systems.
PDF Abstract XML References Citation
Received: July 26, 2015;
Accepted: September 02, 2015;
Published: October 08, 2015
How to cite this article
M. Syafii, I. Cartika and D. Ruswandi, 2015. Multivariate Analysis of Genetic Diversity among some Maize Genotypes under Maize-Albizia Cropping System in Indonesia. Asian Journal of Crop Science, 7: 244-255.
DOI: 10.3923/ajcs.2015.244.255
URL: https://scialert.net/abstract/?doi=ajcs.2015.244.255
DOI: 10.3923/ajcs.2015.244.255
URL: https://scialert.net/abstract/?doi=ajcs.2015.244.255
INTRODUCTION
Maize, together with wheat and rice are the important cereal crops in the world (Rezaeieh and Eivazi, 2011). Indonesia is among the important world maize producer in South East Asia (Ruswandi et al., 2014). Some of the problems encountered in maize development in Indonesia are the conversion of fertile land under maize production into industrial and residential areas, drought due to global climate change, displacement of maize planting area to marginal land including forest and others. One of the possible solutions in responding to the land shortage is incorporating maize in an agroforestry system. In this manner, several needs of farmers can be met and auxiliary sources of food, fodder, fuel-wood can be obtained.
Agroforestry is a land management system, which is a combination of crop production with forestry technologies (De Foresta and Michon, 1996). Albizia is a popular leguminous tree grown for agroforestry in Indonesia. This tree can be found in a very broad distribution of climate and is among the fastest growing timber species. It is also used in pulp, paper, veneer and furniture (Chairomani and Suhaendi, 1994) aside from being environmentally friendly (Nemoto, 2002). The leguminous supply nitrogen, organic materials and recycle leached nutrients (Yamoah et al., 1986). Thus, it has the potential to be used in an agroforestry system with maize, which is called as Maize-Albizia cropping system.
The first step in the development of a Maize-Albizia cropping system is breeding of maize cultivars that are suitable for growth in such a system. In order to achieve this, maize plants should be screened for genetic diversity. The success in developing new high yield cultivars depends on the availability of genetically diverse germplasm (Buckler, 2009). The higher the genetic diversity possessed, the greater the chances of success for developing new superior cultivars. Germplasm improvement and genetic diversity is a key to reliable and sustainable production of the food crops through breeding.
Measuring the available genetic diversity is of utmost importance for effective evaluation and utilization of germplasm (Zubair et al., 2007). Some researchers emphasize the importance of the assessment of the levels and patterns of genetic diversity in crop breeding (Barrett and Kidwell, 1998; Cox et al., 1986; Franco et al., 2001; Habtamu and Milion 2013; Hallaur and Miranda, 1988; Mohammadi and Prasanna, 2003; Thompson et al., 1998). The diverse applications include: (1) Analysis of genetic variability in cultivars, (2) Identification of diverse parental combinations to create segregating progenies with maximum genetic variability for further selection, (3) Introgressing of desirable genes from diverse germplasm into the available genetic base, (4) Study of genetic relationships among inbred lines or pure lines for planning crosses, assigning lines to specific heterotic groups and for precise identification with respect to plant varietal protection, (5) Classification of accessions and identification of subsets of core accessions with possible utility for specific breeding purposes and (6) Identification groups with the same genetic background for preservation, evaluation and utilization of genetic resources.
Multivariate analysis is the most popular approach for genetic variability estimation to study the patterns of variation and their genetic relationships among germplasm collections (Ajmal et al., 2013; Malik et al., 2014). The PCA and cluster analysis are preferred tools for morphological characterization of genotypes and their grouping on similarity basis based on this approach (Mohammadi and Prasanna, 2003; Peeters and Martinelli, 1989). Multivariate analyses have been utilized in many countries for several crops such as wheat (Ajmal et al., 2013; Malik et al., 2014), sorghum (Ali et al., 2011) and maize (Lee et al., 2005; Babic et al., 2010; Azad et al., 2012).
We have previously developed a germplasm collection with more than 500 maize elite inbred lines representing the major tropical breeding programs of Indonesia. Some of the lines in the collection have been described in previous studies, viz. Ruswandi et al. (2014) reported some maize genotypes showing early maturity and also drought tolerance. Some maize hybrid formation showed high yield (Ruswandi et al., 2015a, b). Based on the information on maturity, drought tolerance and yield potential data obtained from the field experiments, 75 lines were chosen for the present study. The objective was to estimate genetic diversity of elite maize inbred collection that can potentially be used in a Maize-Albizia cropping system.
MATERIALS AND METHODS
Genetic materials: Genetic material evaluated in the research included 75 inbred lines of maize developed by Ruswandi, Laboratory of Plant Breeding, Faculty of Agriculture, University of Padjadjaran Indonesia and 2 inbred lines of ICERI (Indonesia Cereals Research Institute). These inbred lines were inbred lines series BR, DR, MBR and MDR. Evaluation of inbred lines in experimental trials was performed in rainy season of 2014 (April-July, 2014), in the community forests planted with 3 year old Albizia trees in 2×2 m. The forests are located in Cimalaka, West Java, Indonesia at 1100 m asl (above sea level), which represents climate type of C3 classified by Oldeman.
Field experimental details: The evaluation of inbred lines was laid on a split plot design with two replications. The main plot consisted of two cropping systems, namely; maize sole cropping system and Maize-Albizia agroforestry system. The subplots had 75 inbred lines of maize. The experimental plot consisted of single rows, 5 m long, with 0.75 m between rows and between plots and 0.20 m within rows. During the crop cultivation, the standard crop management practices were applied and the plots were manually harvested.
The characters included days to anthesis, days to silking, days to harvesting, plant height, length of nodes, plant diameter, chlorophyll content, leaf area index, ear length, ear diameter, number of rows, weight of 1000 seeds and ear weight per plant. The biometric characters (mentioned above) were recorded from 15 randomly chosen competitive plants in each replicate.
Data analysis: The average data were subjected to Analysis of Variance (ANOVA) using Microsoft Office Excel 2010. Principal Component Analysis (PCA) was used to classify maize genotypes and see how big contribution to the character of maize genotype appearance. Genetic diversity and distance based on the similarity between objects under study were analyzed using cluster analysis based on the "t" Euclidian coefficient. The XLSTAT 2012 computer software was used for cluster analysis and PCA.
RESULTS AND DISCUSSION
Analysis of variance: The results of Table 1 show that days to tasseling, days to silking, days to harvesting, node length, leaf area index, chlorophyll content and grain yield is influenced by cropping system, genotype and the interaction between cropping system and genotype.
| Table 1: | ANOVA and coefficient of variance from 14 biometric characters observed |
 | |
| **Significant at 0.01 probability level, UBB: Days to silking; UBJ: Days to tasseling, UP: Days to harvesting, TT: Plant height, PR: Node length, DB: Stem diameter, LD : Leaf area, ILD: Leaf area index, KLO: Chlorophyll content, DT: Ear diameter, PT : Ear length, JB: Rows number, B1000 =1000 seeds weight, Yield: Grain yield per ha | |
| Table 2: | Origin of 75 maize genotypes in the study |
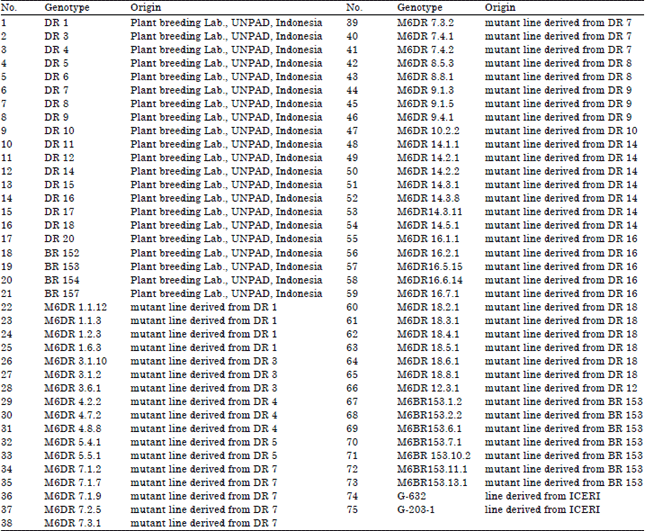 | |
However, characters such as plant height, stem diameter, diameter of ear, length of ear, weight of 1000 seeds were found influenced by genotype and interaction between genotype and cropping system, whereas, the number of rows per ear was found influenced only by the genotype.
Fehr (1987) reported that the level of performance of different genotypes might not be the same if they are planted in different environments and will grow according to their genetic capability. The performance of one plant in one population varies with their genetic make up and the environment affecting it (Poehlman and Sleper, 1995).
The efficiency of one trial can be determined in terms of its Coefficient of Variation (CV). The greater the CV, the efficiency is lower. A higher CV shows a decrease in accuracy of the trial. The ideal CV should be less than 20%, which indicates a higher efficiency of the results. Table 2 shows that in all characters, the CV is less than 20%, which indicates the accuracy of the results.
Principal component analysis: Multivariate analysis is used to analyze a group of data with more than one variable. One of the multivariate analyses is Principal Component Analysis (PCA), which aims to reduce the dimension from a group of data with a higher number of variables that are interrelated (Jolliffe, 2002).
| Table 3: | Principle component analysis of various morpho-physiological traits in maize under conditions maize sole cropping system and Maize-Albizia cropping system |
 | |
| Table 4: | Factor loadings morpho-physiological trait markers contributed to variability in maize sole cropping system |
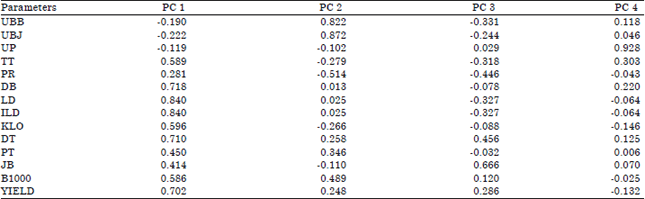 | |
| UBB: Days to silking, UBJ: Days to tasseling, UP: Days to harvesting, TT: Plant height, PR: Node length, DB : Stem diameter, LD: Leaf area, ILD: Leaf area index, KLO: Chlorophyll content, DT: Ear diameter, PT: Ear length, JB: Rows number, B1000 =1000 seed weight, Yield: Grain yield per ha | |
| Table 5: | Factor loadings morphological trait markers contributed to variability in Maize-Albizia cropping system |
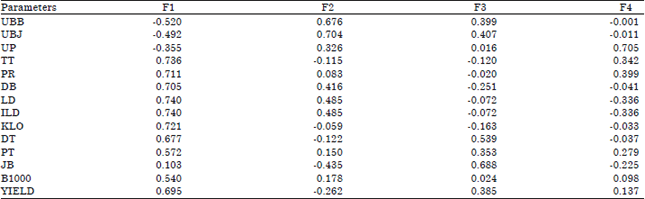 | |
| UBB: Days to silking, UBJ: Days to tasseling, UP: Days to harvesting, TT: Plant height, PR: Node length, DB: Stem diameter, LD: Leaf area, ILD: Leaf area index, KLO: Chlorophyll content, DT: Ear diameter, PT: Ear length, JB: Rows number, B1000 =1000 seeds weight, Yield: Grain yield per ha | |
Main component analysis is a technique used to determine the contribution of one character on variability to easily determine the character that can represent a genotype (Afuape et al., 2011). The PCA is also used to describe a genotypic data to look for characters that have high contribution towards variability shown within the biplot.
Results of the PCA analysis showed four main axial components that have Eigen values more than 1.00 (Table 3). Table 4 and 5 shows the Principal Component (PC) that forms variability from 14 characters observed. The determination of the PC is based on an eigenvalue more than 1.00 (Jeffer, 1967) and a cumulative percentage more than 80% (Jolliffe, 2002). For each PC, characters are with PC values more than 0.70 (Jeffer, 1967).
In maize sole cropping system, the Eigen value ranged from 1.08-4.52 which contributed to 67.27% variability. Primary component 1 (PC 1) covers 32.31% of variability from a variation of 75 genotypes given the biometric characters namely; stem width, leaf width, leaf area, leaf area index, ear width and yield per hectare. The PC 2 covers 16.86% variability from variations in 75 genotypes given the characters days to tasseling and days to silking. PC 3 covers 10.32% variability given the character rows number. For PC 4, it covers 7.77% variability given the character days to harvesting.
Under Maize-Albizia cropping system, the Eigen value was between 1.16-5.37 which gives a variability of as high as 71.85%. The PC 1 covered 38.31% of variability from a variation of 75 genotypes given the characters plant height, node length, stem length, leaf area and chlorophyll content. The PC 2 showed 14.66% variability given the characters days to tasseling and for PC 4, variability was at 8.28% given the character days to harvesting. For both sets of cropping, maize sole cropping system and Maize-Albizia cropping system, the one that gave a variability contribution were stem diameter, leaf area, leaf area index, days to tasseling and days to harvesting.
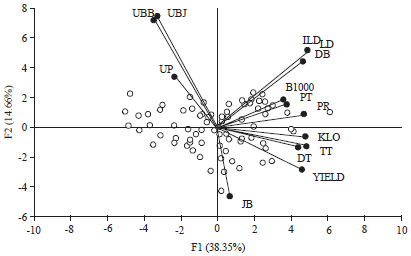
The scatter plots are shown in a graphic biplot as determined by the PC values that give the highest contribution to variability. The PC 1 and PC 2 are the component values that give the highest contribution towards variability of a character. Under maize sole cropping system, the PC 1 and PC 2 values are 32.31 and 16.86%, respectively, whereas, under Maize-Albizia cropping system, the PC 1 and PC 2 values were 38.31 and 14.66%, respectively.
The variability values from the two components for each maize genotype are scattered within the biplot forming 4 quadrants (Fig. 1 and 2). The figures show that the different morpho-physiological and maize genotypes are found in four different quadrants. In the study by Worede et al. (2014), PC 1 and PC 2 that showed cumulative variabilities as high as 61.16% were used. This shows that there are characters in the PC that have relatively high variability and are important in separating genotypes. Under maize sole cropping system (Fig. 1), quadrant I has a positive value showing the biometric characters viz. leaf area index, leaf area, yield per hectare and weight per 1000-seeds and ear length are tightly related or linked.
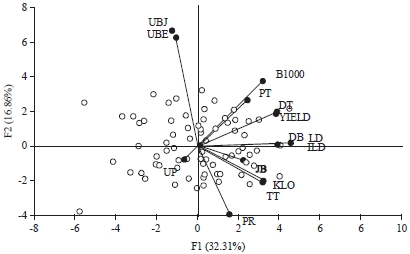 | |
| Fig. 1: | Distribution pattern of 75 maize inbred in maize sole cropping system based on 14 morpho-physiological characters |
 | |
| Fig. 2: | Distribution pattern of 75 maize inbred in Maize-Albizia cropping system based on 14 morpho-physiological characters |
Genotypes in quadrant I (DR 4, DR 6, DR 9, DR 10, DR 11, DR 12, DR 14, DR 16, DR 17, DR 18, DR 20, BR 154, M6DR 16.1.1, M6DR 18.5.1, M6DR 16.6.14, M6DR 3.1.10, M6BR 153.2.2, M6DR 1.1.3 and M6DR 1.1.12) are closely related and they have high level of similarities. For Maize-Albizia cropping system, (Fig. 2), characters with positive values are leaf area index, leaf area, stem diameter, weight per 1000 seeds, ear length and node length. Genotypes that have high similarities are M6DR 1.6.3, M6DR 8.5.3, M6DR 16.6.14, M6DR 3.1.10, M6DR 1.1.3, M6DR 7.3.1, M6DR 16.2.1, M6DR 14.5.1, M6DR 1.2.3, M6DR 14.2.2, M6DR 7.4.2, M6DR 16.1.1, DR 4, DR 6, DR 7, DR 9, DR 10, DR 11, DR 12, DR 20, BR 153. Different characters in the same quadrant mean that they are closely and positively related. The opposite is true if they are in different quadrants.
Agglomerative Hierarchical Clustering (AHC) analysis: Cluster analysis is a method used in grouping a set of characters into clusters. Genotypic clustering makes use of a procedure called Agglomerative Hierarchical Clustering (AHC) using Unweighted Pair Group Method with Arithmetic mean (UPGMA) mentioned by Mohammadi and Prasanna (2003). Cluster analysis is one of the statistical techniques aimed in grouping objects in clusters so that the objects in one cluster have high similarities than those in other clusters (Sharma, 1996). Euclidian distance values in the range of 0-1 indicates a small dissimilarity, whereas, its value more than 1 indicates a large dissimilarity coefficient. A small dissimilarity coefficient indicates that for each genotype one or the other characters have a narrow variability (Tairo et al., 2008). Cluster analysis for 75 genotypes using 14 morpho-physiological characters is presented in Fig. 3 (maize sole cropping system) and Fig. 4 (Maize-Albizia cropping system). Morphological traits have been successfully used for estimation of genetic diversity and cultivar development since they provide a simple way of quantifying genetic variation (Fufa et al., 2005).
In maize sole cropping system, clustering resulted in nine main clusters with a point Euclidian coefficient of 4.8, level of dissimilarity between genotypes is high in the dendogram with a genetic distance of 0-0.8. In Maize-Albizia cropping system, five main clusters were formed with a point Euclidian coefficient of 5.1.
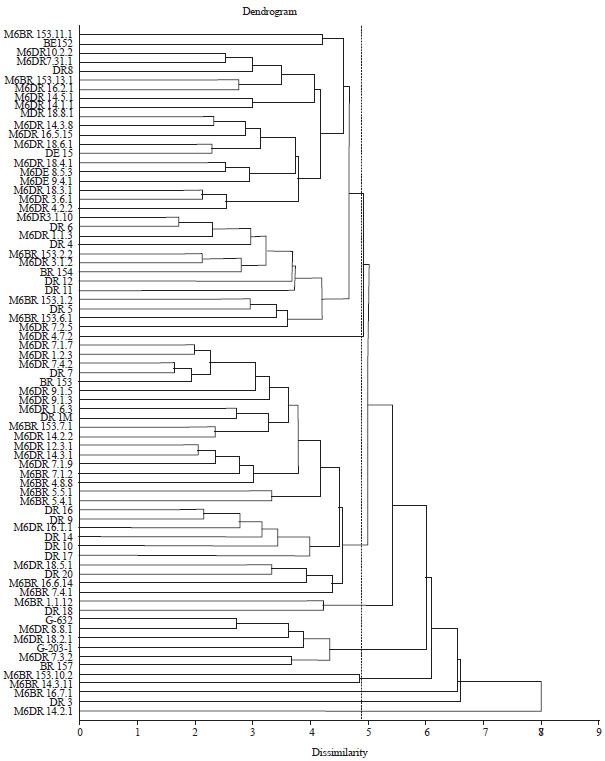 | |
| Fig. 3: | Dendogram for 75 maize inbred based on morpho-physiological characters in maize sole cropping system |
The level of dissimilarity among genotypes in the dendogram has a distance of 0-6.4. From the dendogram results, the clustering of genotypes based on the morpho-physiological characters is as shown in Table 6. The dendogram shows that the distribution of the 75 genotypes is wide. Nine and five clusters were obtained in the dendogram forming a wide distance from its cluster.
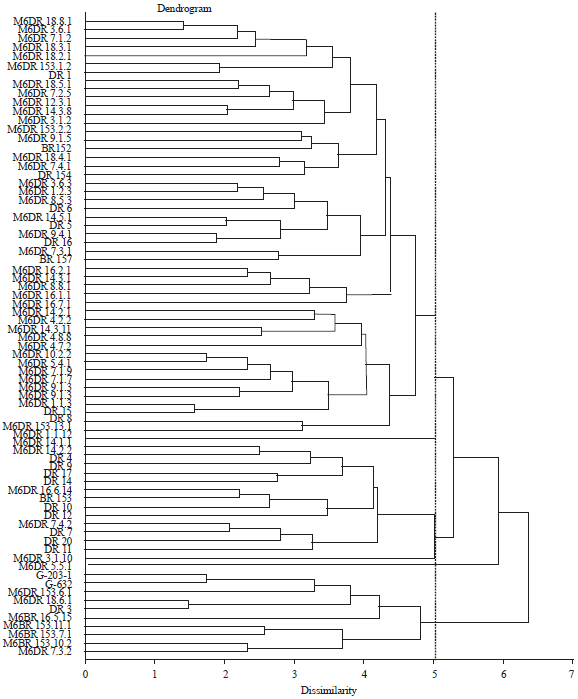 | |
| Fig. 4: | Dendogram for 75 maize inbred based on morpho-physiological characters in Maize-Albizia cropping system |
Morpho-physiological diversity in the materials studied is structured by genotypes and this diversity could be utilized for cultivar breeding and germplasm conservation programs (Ali et al., 2011; De Jesus et al., 2013), particularly in breeding for Maize-Albizia cropping system.
CONCLUSION
Based on the results of the experiment, it can be concluded that there is a high variability in the morpho-physiology in the maize genotypes from UNPAD. This is found in almost all the characters observed in maize sole cropping system and Maize-Albizia cropping system. Multivariate analysis showed high genetic variability in all genotypes in both cropping system.
| Table 6: | Genotypes accessions related to various clusters based on various morpho-physiological traits |
 | |
ACKNOWLEDGMENTS
The research fund was granted by the program KKP3N Agricultural Research and Development Agency Ministry of Agriculture, Republic of Indonesia 2014-2015, whereas, the publication was funded by PUPTN 2015 research grant to Dedi Ruswandi.
REFERENCES
- Afuape, S.O., P.I. Okocha and D. Njoku, 2011. Multivariate assessment of the agromorphological variability and yield components among sweet potato (Ipomoea batatas (L.) Lam) landraces. Afr. J. Plant Sci., 5: 123-132.
Direct Link - Ajmal, S.U., N.M. Minhas, A. Hamdani, A. Shakir, M. Zubair and Z. Ahmad, 2013. Multivariate analysis of genetic divergence in wheat (Triticum aestivum) germplasm. Pak. J. Bot., 45: 1643-1648.
Direct Link - Ali, M.A., K. Jabran, S.I. Awan, A. Abbas and M. Zulkiffal et al., 2011. Morpho-physiological diversity and its implications for improving drought tolerance in grain sorghum at different growth stages. Aust. J. Crop Sci., 5: 311-320.
Direct Link - Azad, M.A.K., B.K. Biswas, N. Alam and Sk.S. Alam, 2012. Genetic diversity in maize (Zea mays L.) inbred lines. Agriculturists, 10: 64-70.
CrossRefDirect Link - Babic, V., Z. Pajic, S. Prodanovic, M. Babic and M. Filipovic, 2010. Visual assessment of sweet maize lines phenotype, according to UPOV descriptor, as indicator of heterosis. Genetika, 42: 313-322.
CrossRefDirect Link - Barrett, B.A. and K.K. Kidwell, 1998. AFLP-based genetic diversity assessment among wheat cultivars from the Pacific Northwest. Crop Sci., 38: 1261-1271.
CrossRefDirect Link - Cox, T.S., J.P. Murphy and D.M. Rodgers, 1986. Changes in genetic diversity in the red winter wheat regions of the United States. Proc. Natl. Acad. Sci., 83: 5583-5586.
PubMedDirect Link - De Foresta, H. and G. Michon, 1996. The agroforest alternative to Imperata grasslands: When smallholder agriculture and forestry reach sustainability. Agrofor. Syst., 36: 105-120.
CrossRefDirect Link - De Jesus, O.N., J.P.X. de Freitas, J.L.L. Dantas and E.J. de Oliveira, 2013. Use of morpho-agronomic traits and DNA profiling for classification of genetic diversity in papaya. Genet. Mol. Res., 12: 6646-6663.
Direct Link - Fehr, W.R., 1987. Principles of Cultivar Development. Vol. 1, Macmillan Publishing Co., New York.
Direct Link - Franco, J., J. Crossa, J.M. Ribaut, J. Bertran, M.L. Warburton and M. Khairallah, 2001. A method for combining molecular markers and phenotypic attributes for classifying plant genotypes. Theoret. Applied Genet., 103: 944-952.
CrossRefDirect Link - Fufa, H., P.S. Baenziger, B.S. Beecher, I. Dweikat, R.A. Graybosch and K.M. Eskridge, 2005. Comparison of phenotypic and molecular marker-based classifications of hard red winter wheat cultivars. Euphytica, 145: 133-146.
CrossRefDirect Link - Habtamu, S. and F. Million, 2013. Multivariate analysis of some Ethiopian field pea (Pisum sativum L.) genotype. Int. J. Genet. Mol. Biol., 5: 78-87.
Direct Link - Jeffer, J.N.R., 1967. Two case studies in the application of principal component analysis. J. R. Statist. Soc. Ser. C (Applied Statist.), 16: 225-236.
CrossRefDirect Link - Lee, K.M., T.J. Herrman, J. Lingenfelser and D.S. Jackson, 2005. Classification and prediction of maize hardness-associated properties using multivariate statistical analyses. J. Cereal Sci., 41: 85-93.
CrossRefDirect Link - Mohammadi, S.A. and B.M. Prasanna, 2003. Analysis of genetic diversity in crop plants-salient statistical tools and considerations. Crop Sci., 43: 1235-1248.
CrossRefDirect Link - Malik, R., H. Sharma, I. Sharma, S. Kundu and A. Verma et al., 2014. Genetic diversity of agro-morphological characters in Indian wheat varieties using GT biplot. Aust. J. Crop Sci., 8: 1266-1271.
Direct Link - Peeters, J.P. and J.A. Martinelli, 1989. Hierarchical cluster analysis as a tool to manage variation in germplasm collections. Theor. Applied Genet., 78: 42-48.
CrossRefDirect Link - Rezaeieh, K.A. and A. Eivazi, 2011. Evaluation of morphological characteristics in five Persian maize (Zea mays L.) under drought stress. Afr. J. Agric. Res., 6: 4409-4411.
Direct Link - Ruswandi, D., Agustian, E.P. Anggia, A.O. Canama, H. Marta, S. Ruswandi and E. Suryadi, 2014. Mutation breeding of maize for anticipating global climate change in Indonesia. Asian J. Agric. Res., 8: 234-247.
CrossRefDirect Link - Ruswandi, D., J. Supriatna, A.T. Makkulawu, B. Waluyo, H. Marta, E. Suryadi and S. Ruswandi, 2015. Determination of combining ability and heterosis of grain yield components for maize mutants based on line x tester analysis. Asian J. Crop Sci., 7: 19-33.
CrossRefDirect Link - Ruswandi, D., J. Supriatna, B. Waluyo, A.T. Makkulawu, E. Suryadi, Z.U. Chindy and S. Ruswandi, 2015. GGE biplot analysis for combining ability of grain yield and early maturity in maize mutant in Indonesia. Asian J. Crop Sci., 7: 160-173.
CrossRefDirect Link - Tairo, F., E. Mneney and A. Kullaya, 2008. Morphological and agronomical characterization of sweet potato [Ipomoea batatas (L.) Lam.] germplasm collection from Tanzania. Afr. J. Plant Sci., 2: 77-85.
Direct Link - Thompson, J.A., R.L. Nelson and L.O. Vodkin, 1998. Identification of diverse soybean germplasm using RAPD markers. Crop Sci., 38: 1348-1355.
CrossRefDirect Link - Worede, F., T. Sreewongchai, C. Phumichai and P. Sripichitt, 2014. Multivariate analysis of genetic diversity among some rice genotypes using morpho-agronomic traits. J. Plant Sci., 9: 14-24.
CrossRefDirect Link - Yamoah, C.F., A.A. Agboola, G.F. Wilson and K. Mulongoy, 1986. Soil properties as affected by the use of leguminous shrubs for alley cropping with maize. Agric. Ecosyst. Environ., 18: 167-177.
CrossRefDirect Link - Zubair, M., S.U. Ajmal, M. Anwar and M. Haqqani, 2007. Multivariate analysis for quantitative traits in mungbean [Vigna radiata (L.) Wilczek]. Pak. J. Bot., 39: 103-113.
Direct Link