
W. Pilahome
Department of Biology, Faculty of Science, Khon Kaen University, Khon Kaen, 40002, Thailand
S. Bunnag
Center of Excellence of Agricultural Biotechnology (Ag-BIO/PERDO-CHE) Bangkok, 10900, Thailand
A. Suwanagul
Department of Agricultural Technology, Thailand Institute of Scientific and Technological Research (TISTR), Pathumthani, 12120, Thailand
Asian Journal of Crop Science
Year: 2014 | Volume: 6 | Issue: 4 | Page No.: 305-319







ABSTRACT
This study was carried out to establish a simple, efficient and reproducible protocol for plant regeneration in four rice genotypes which are Pokkali, RD6, Sakon Nakhon and Niaw Ubon 2. Two protocols were proposed and compared in this study, including (1) Multiple shoots induction and rooting and (2) Callus induction and regeneration. Single seeds of each cultivar were sown and germinated seed and shoot segments were used as explant source for the experiments. The results showed that a great number of multiple shoots induced from germinated seed could be achieved on MS solid medium supplemented with 50 μM 6-BAP alone, whereas rooting of the obtained shoots was easily induced on hormone-free MS solid medium. Compact callus formation could be induced from shoot segments on MS solid medium amended with 2 mg L-1 2,4-D alone, whilst a combination of 0.5 mg L-1 NAA and 2.0 mg L-1 6-BAP was most effective in promoting shoot regeneration from compact calli. Our study suggests that the regeneration frequency obtained via callus induction was inferior to that yielded through multiple shoots induction. The procedure reported here is expected to be useful in micropropagation and in vitro breeding and selection for drought and/or salt tolerance in rice for increasing the yield potential of this vital crop.
PDF Abstract XML References Citation
Received: May 22, 2014;
Accepted: July 16, 2014;
Published: August 27, 2014
How to cite this article
W. Pilahome, S. Bunnag and A. Suwanagul, 2014. Development of a Plant Regeneration System from Seed-derived Shoot Segments of Rice (Oryza sativa L.). Asian Journal of Crop Science, 6: 305-319.
DOI: 10.3923/ajcs.2014.305.319
URL: https://scialert.net/abstract/?doi=ajcs.2014.305.319
DOI: 10.3923/ajcs.2014.305.319
URL: https://scialert.net/abstract/?doi=ajcs.2014.305.319
INTRODUCTION
Rice (Oryza sativa L.) is an important versatile cereal crop that feeds over half of the world’s population. It is estimated that more than 400 million people in rice-producing areas of Asia, Africa and South America still endure chronic hunger with the demand for food expected to increase by another 38% within 30 years (Surridge, 2004). However, the productivity of rice in many areas is affected by several abiotic and biotic stresses due to narrow genetic diversity present in the existing rice cultivars as consequence of human selection for early maturing and high yielding rice genotypes (Mazid et al., 2013). Water deficit and salinity are among the most important growth-limiting factors for plants species in arid and semi-arid regions of the world (Levitt, 1980). In Thailand, 62% (6.08x106 tons ha-1) of rice-growing areas are located in the northeastern part of the country but due to water shortage and soil salinity problems arising from the presence of underground salt domes, rice productivity from this area has been relatively low in comparison to that from the central plain which hardly experiences water shortage and has no saline soil (Nounjan et al., 2012). Improvement of this vital crop with desirable characteristics is, therefore, urgently needed. As has been well-documented by several studies that field trials to establish plants with preferable traits require large rice-producing areas at certain seasons, where the environment is uncontrollable, time-consuming and laborious (Anzoua et al., 2010; Boling et al., 2011; Tsubo et al., 2009; Wang et al., 2013; Wu et al., 2013), plant biotechnology is considered as an alternative procedure to produce rice plants with increased tolerance to biotic and abiotic stresses.
For improvement of rice via biotechnology, plant tissue culture is globally regarded as an attractive tool since it offers the possibility to produce plants with desirable characteristics, as well as potentially generating fertile plants under aseptic conditions (Ramesh et al., 2009). The advantages of this viable technique are that it allows for large-scale production of plants in limited space under controlled environments where light intensity, temperature and nutrient compositions in culture medium can be adjusted. In addition, this technique is neither time-consuming nor laborious (Reinert and Bajaj, 1995). Over the past three decades, numerous tissue culture techniques have been developed for rice improvement all over the world (Raina, 1989) using various explant materials such as endosperms (Breitenbach et al., 2014), seeds (Xiao et al., 2007), embryos (Zhao et al., 2013) and leaf bases (Karthikeyan et al., 2011; Ramesh et al., 2009).
Establishment of a simple and reproducible protocol for plant regeneration in rice is an essential prerequisite in any in vitro rice improvement programs. The literature has revealed that numerous agronomically valuable rice genotypes are recalcitrant to in vitro manipulation due to their poor callus production and regeneration capability. Hartke and Lorz (1989) evaluated callus formation and plant regeneration in 15 indica rice lines and observed that, only seven of them produced embryogenic calli whilst, only four regenerated into whole plants. Development of a reliable protocol for plant regeneration from induced calli following a stress acclimatization process is of particular interest because the literature has suggested that majority of calli subjected to the acclimatization process fail to regenerate into whole plants (Basu et al., 2002; Winicov, 1996; Zinnah et al., 2013). Therefore, the present study was carried out to establish a simple, efficient and reproducible protocol for callus induction and plant regeneration in rice. In this context, four rice genotypes which are the salt-tolerant Pokkali, the salt-sensitive cultivars RD6 and Sakon Nakhon and the moderately salt-tolerant Niaw Ubon 2 were evaluated for their callus induction and regeneration ability. Two different plant production procedures which are (1) Multiple shoots induction and rooting and (2) Callus induction and plant regeneration, were included in this study. The protocols proposed in this study are expected to be used to increase the frequency of plant regeneration in an in vitro rice improvement program in cultivars of commercial and ecological importance.
MATERIALS AND METHODS
Plant materials: Seeds of four rice (Oryza sativa L.) genotypes were provided by Pathum Thani Rice Research Center, Khon Kaen Rice Research Center, Sakon Nakhon Rice Research Center and Ubon Ratchathani Rice Research Center. The salt-sensitive cultivars RD6 and Sakon Nakhon and the moderately salt-tolerant Niaw Ubon 2 were improved cultivars commonly grown in Thailand. Pokkali, the salt-tolerant landrace from India, was included for comparison. Single seeds of each rice cultivars were used for producing multiple shoot buds in vitro. In brief, dehusked seeds were surface-sterilized in 70% v/v ethanol for 5 min followed by subsequent immersion in 50% commercial bleach (Haiter®, containing 6% sodium hypochlorite) for 5 min, rinsed thrice using sterile distilled water and transferred to hormone-free Murashige and Skoog (MS) solid medium (Murashige and Skoog, 1962) for seed germination and germinated seeds were used as explants for in vitro culture.
Multiple shoots induction and rooting: For shoot bud induction, germinated seeds were cultivated on MS solid medium supplemented with 3% w/v sucrose, 0.2% w/v gellan gum (Kelcogel®) and various concentrations of 6-BAP (0, 25, 50 and 75 μM). The cultures were maintained for 8 weeks at 25°C under 16 h photoperiod regimes provided by cool-white fluorescent lights with 40 μmol m-2 sec-1 illumination. Following 8 weeks of culture, the obtained multiple shoots were separated into single shoots, each of which was dissected into 1 cm segments before being cultured on the same medium for multiplication of multiple shoots. Subculturing of newly regenerated shoots to freshly prepared medium was performed at four-week intervals. For rooting, multiple shoots of each rice cultivars were transferred to hormone-free MS solid medium for four weeks.
Callus induction and plant regeneration: Multiple shoots obtained from previous experiments were dissected into small segments (about 0.5 cm long). The shoot segments were then cultivated on MS solid medium amended with various concentrations of 2,4-dichlorophenoxyacetic acid (2,4-D) (0, 1, 2 and 4 mg L-1) for 4 weeks for callus induction. For plant regeneration, the obtained calli were transferred to MS solid medium containing 0.5 mg L-1 NAA, 300 mg L-1 casein hydrolase and different combinations of 6-benzylaminopurine (6-BAP) (0, 1, 2 and 3 mg L-1) and kinetin (Kn) (0, 1, 2 and 3 mg L-1) for 8 weeks. The cultures were subcultured to freshly prepared medium at four-week intervals.
Growth parameters and water content analysis: Callus fresh weight was determined and then the calli were dried in a hot-air oven at 60°C for a few days until the dry weight was stabilized. Water content was determined using the following equation:
where, DW is callus dry weight and FW is callus fresh weight. Callus formation rate was determined using the following equation:
where, RCF is callus formation rate, NCPS is number of callus-producing shoot segments, NTS is total number of shoot segments. Plant regeneration rate was calculated using the following equation:
where, RPG is plant regeneration rate, NPPC is number of plant-producing calli and NTC is total number of calli.
Statistical analysis: The experiment was arranged in a randomized complete block. Statistical analysis of data was processed using analysis of variance (one-way ANOVA) and all means were separated at the p<0.05 level using the DMRT method. All calculations and data analyses were performed using the SPSS 11.5 for Windows software package.
RESULTS
Multiple shoots formation: Germination of rice occurred 1-2 days after sowing in vitro. The percentage of germination was relatively high, ranging from 98-100%. Green shoot buds started to form 15-20 days after culture of germinated seeds on MS solid medium supplemented with various concentrations of 6-BAP. Initially, numerous aggregates of green shoot buds were produced around germinated seeds. The obtained shoot buds then developed into multiple shoots within 3-4 weeks of culture. Germinated seeds of all the cultivars did not produce aggregates of green shoot buds even after 8 weeks of culture in the absence of 6-BAP. The average number of multiple shoots produced per green shoot bud clumps following a 8-week culture period ranged from 3.00 shoots in Pokkali to 30.33 shoots in Sakon Nakhon (Fig. 1). The characteristics of shoots produced from germinated seed are depicted in Fig. 2. The multiple shoots induction was noted to be associated with the concentration of 6-BAP applied. MS solid medium containing 50 μM 6-BAP was most efficient in inducing formation of multiple shoots in all the cultivars with Sakon Nakhon (Fig. 1c and 3c) showing the highest number of multiple shoots (30.33 shoots) followed by RD 6 (Fig. 1b and 3b) with 16.00 shoots, Niaw Ubon 2 (Fig. 1d and 3d) with 15.00 shoots and Pokkali (Fig. 1a and 3a) with 8.00 shoots.
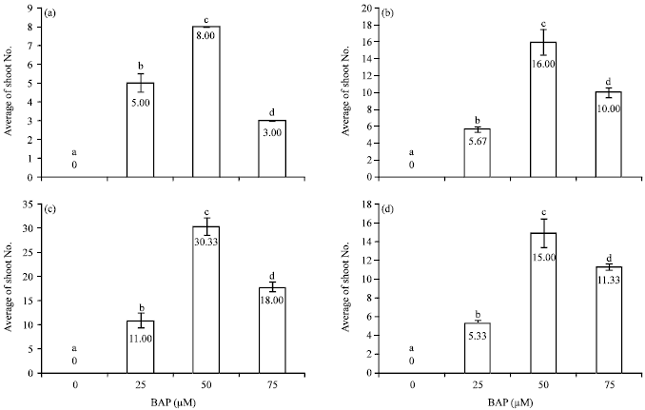 | |
| Fig. 1(a-d): | Mean number of shoots produced from germinated seeds of individual cultivars (a) Salt-tolerant Pokkali, (b) Salt-sensitive RD6, (c) Salt-sensitive Sakon Nakhon and (d) Moderately salt-tolerant Niaw Ubon 2. Different letters above bars indicate significant differences at p<0.05 analyzed by DMRT |
 | |
| Fig. 2(a-c): | Characteristics of shoot buds and shoots produced from germinated seed explants (a) Shoot buds clumps produced from germinated seed, (b) Control plants produced form germinated seeds and (c) Multiple shoots developed from shoot bud clumps |
 | |
| Fig. 3(a-d): | Multiple shoots produced from shoot segments of individual cultivars (a) Salt-tolerant Pokkali, (b) Salt-sensitive RD6, (c) Salt-sensitive Sakon Nakhon and (d) Moderately salt-tolerant Niaw Ubon 2 |
Shoot rooting and transplanting: Rooting of shoots was easily induced on hormone-free MS solid medium. Roots began to generate on the 5th day of culture. After 4 weeks, roots developed to about 4-8 cm.
 | |
| Fig. 4(a-d): | Rooting of the obtained shoots in individual cultivars (a) Salt-tolerant Pokkali, (b) Salt-sensitive RD6, (c) Salt-sensitive Sakon Nakhon and (d) Moderately salt-tolerant Niaw Ubon 2 |
The cultivar Sakon Nakhon (Fig. 4c) exhibited the highest root length (8.38 cm) followed by Niaw Ubon 2 (Fig. 4d), RD 6 (Fig. 4b) and Pokkali (Fig. 4a). Plants with roots were transplanted to the plastic pots and the survival rate of transplanted plants was up to 95%.
Callus induction and subculture: Calli started to form 7-10 days after culture of shoot segments on MS solid medium supplemented with various concentrations of 2,4-D. Initially, induced calli were compact and yellow (Fig. 5a-b) but most of them became watery and sticky light-yellow following 4-5 weeks of subculture. Shoot segments did not produce calli even after 4 weeks of culture in the absence of 2,4-D. The percentage of callus induction after 4 weeks of culture ranged from 32.0% in the cultivar Niaw Ubon 2-88% in the cultivar Pokkali (Table 1). Accompanied by growth, compact calli produced phenolic compounds that resulted in browning and subsequent death of the calli. Although, all the cultivars produced healthy calli, significant quality differences were observed between the cultivars. Fresh weight of the obtained callus clumps was highest (1.02 g) in RD6 and lowest (0.13 g) in Niaw Ubon2. Similar results were noted for dry weight of the obtained callus clumps, where the highest value (0.14 g) was observed in RD6 and the lowest value (0.02 g) was obtained in Niaw Ubon2. However, water content of callus clumps was highest (90.01%) in Sakon Nakhon and lowest (73.10%) in Pokkali.
 | |
| Fig. 5(a-i): | Callus induction and plant regeneration in individual cultivars. (a-b) Yellow,compact calli induced from shoot segments, (c) Aggregates of green shoot buds, (d) Multiple shoots developed from green shoot buds, (e) 5-8 week-old whole plants and (f-i) Charateristics of regenerated plants of the salt-tolerant Pokkal the salt-sensitive RD6, the salt-sensitive SakonNakhon and the moderatelysalt-tolerant Niaw Ubon 2, respectively |
| Table 1: | Frequency of callusing, callus fresh and dry weight and callus water content observed in four rice cultivars on MS solid medium supplemented with various concentrations of 2,4-D |
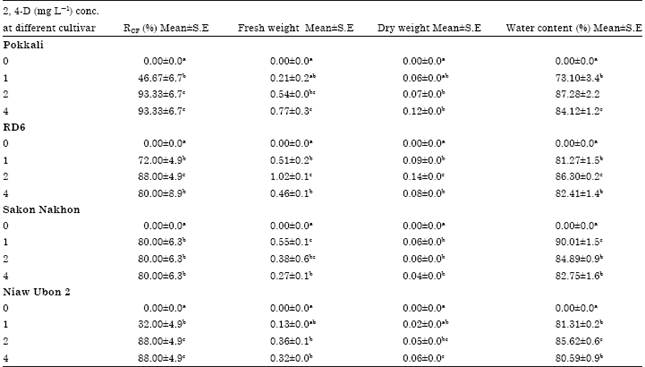 | |
| *Significance at p<0.05. Means followed by the same letter within each column are not significantly different using Duncan’s multiple range test at 95% | |
Plant regeneration from shoot segments-derived calli: In all the cultivars tested, globular bodies initially produced from shoot segments-derived calli did not develop into plantlets on callus induction medium. To induce organogenesis, globular bodies were transferred onto MS solid medium amended with 0.5 mg L-1 NAA, 300 mg L-1 casein hydrolysate and different combinations of 6-BAP and Kn. After 4 weeks of culture, globular bodies of all the cultivars formed adventitious roots on MS medium supplemented with 0.5 mg L-1 NAA and 300 mg L-1 casein hydrolysate but shoots were not produced except in the cultivar Sakon Nakhon (Table 2). The addition of 6-BAP and/or Kn promoted the formation of roots and shoots in all the cultivars. Under preferred conditions, globular bodies produced aggregates of green shoot buds (Fig. 5c) and subsequently the obtained shoot buds further developed into multiple shoots (Fig. 5d). After 5-8 weeks of culture, whole plants were obtained (Fig. 5e). MS solid medium supplemented with 0.5 mg L-1 NAA, 300 mg L-1 casein hydrolysate and 2 mg L-1 6-BAP was most effective in promoting plant regeneration in all the cultivars with the cultivar Sakon Nakhon (Fig. 5h) showing the highest frequency of plant regeneration (31.1%) followed by RD6 (Fig. 5g) (22.2%) and Pokkali (Fig. 5f) and Niaw Ubon 2 (Fig. 5i) (17.8%).
DISCUSSION
Plant tissue culture systems offer the opportunity to select and regenerate plants with desirable characteristics and are effectively utilized to produce plants with desired phenotypes in many plants species (Aazami et al., 2010; Soliman and Hendawy, 2013; Verma et al., 2013). In general, successful improvement of plants in vitro requires a simple, efficient and reproducible protocol for plant regeneration (Joshi et al., 2011; Lu et al., 2007) which is affected by several parameters, including explant materials.
| Table 2: | Frequency of regeneration and rooting in four rice cultivars on MS solid medium supplemented with various concentrations of 6-BAP and Kn |
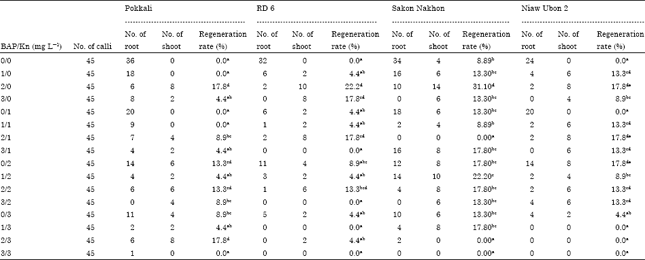 | |
| *Significance at p<0.05. Means followed by the same letter within each column are not significantly different using Duncan’s multiple range test at 95% | |
Even though, in vitro technique has been employed for successful improvement in several rice genotypes (Khanna and Raina, 1999; Mohanty et al., 2002; Zinnah et al., 2013), the possibility of successful improvement using this technique is limited in indica rice varieties because of low regeneration efficiency (Raghavendra et al., 2009).
As has been reported that successful plant regeneration can be achieved via direct shoot induction and several protocols have been established for many important cereal crops including oat (Zhang et al., 1996), maize (Zhang et al., 2002), sorghum (Baskaran and Jayabalan, 2005; Baskaran et al., 2006; Bhaskaran et al., 1992), barley (Sharma et al., 2004) and wheat (Sharma et al., 2005), we performed direct shoot induction in four rice genotypes to investigate whether this technique should be used in rice. As regeneration capability in rice varies greatly across genotypes, it is essential to optimize the in vitro regeneration conditions for each cultivar since it is a key step for plant manipulation. In our study, germinated seeds were used as explants for direct shoot regeneration and a great number of multiple shoots were obtained on a multiple shoots-induction medium amended with the cytokinin 6-BAP alone. The proposed protocol, in our study, was quite similar to the protocol established in previous studies that the cytokinin thidiazuron (TDZ) was applied alone or in combination with 6-BAP for improving plant regeneration in rice and other cereal crops (Ganeshan et al., 2006; Raghavendra et al., 2009). Based on our study, neither of albino plants and abnormal phenotypes was regenerated, irrespective of the plant growth regulators or combinations thereof. The obtained plants grew normally after being transferred to new medium for rooting.
Calli have been have been mainly used as explant materials for in vitro improvement of rice through mutant induction (Bajji et al., 2000; Mahmood et al., 2014; Matheka et al., 2008). In general, mutation induction in rice is generally on the basis of prolonged exposure of callus explants to a selected mutagenic substance along with repeated subculturing which usually leads to decreased regeneration efficiency of the callus explants. A suitable methodology to enhance plant regeneration efficiency during improvement of rice via mutation induction is therefore, urgently needed. In our study, compact calli were induced from shoot segments with high frequency using 2,4-D alone, indicating that these explants are a good source for callus induction in the four rice cultivars. It is well-known that plant growth regulators play a crucial role in callus induction. In most cases, callus induction in monocots, particularly rice, usually requires a strong auxin such as 2,4-D. Manimaran et al. (2013) induced callus formation on MS solid medium supplemented with 2 mg L-1 2,4-D and 0.5 mg L-1 Kn as callus-inducing agents. Che et al. (2006) observed that gene expression patterns change extensively during somatic embryo maturation and germination and a progressive decline in the expression of genes involved in cell proliferation and growth, such as genes encoding histones and ribosomal proteins throughout embryo maturation was noted when the calli were transferred to medium lacking 2,4-D. Bronsema et al. (2001) described that atleast 0.2 mg L-1 2,4-D is required to exceed the threshold level for the transition from germination to callus induction. Another study has supported that the use of 1-3 mg L-1 2,4-D to induce callus from immature embryos of maize is a critical factor. Moreover, Armstrong and Green, (1985) supported that 1-3 mg L-1 2,4-D was considered a critical factor for inducing callus formation from immature embryos of maize and the addition of 2.0-3.0 mg L-1 2,4-D in culture medium was essential for embryogenic callus formation. Although the application of 2,4-D alone could induce callus formation from rice embryos (Ge et al., 2006; Lee et al., 2002; Thadavong et al., 2002), enhanced efficiency of callus formation could be achieved by the addition of some organic substances (such as tryptophan, casein hydrolysate, proline) together with 2,4-D to callus-induction medium (Khaleda and Al-Forkan, 2006; Rattana et al., 2012; Thadavong et al., 2002). Plant regeneration from calli has been successfully achieved in many monocots (Ntui et al., 2010; Yuan et al., 2009; Zhong et al., 2011). However, plant regeneration from calli has been reported to be relatively low in rice (Basu et al., 2002; Winicov, 1996; Zinnah et al., 2013). In our study, the frequency of plant regeneration from calli under non-optimized conditions was quite low. The frequency was, however, enhanced by the addition of NAA and Kn to a regeneration medium; the frequency under optimal conditions was found to increase up to 31.1%. The findings in our study were consistent with an earlier study in rice (Manimaran et al., 2013) that the supplementation of NAA and Kn to a regeneration medium could regenerate more plants with good number of roots.
It has been well-known that several factors play a crucial role in callus development into whole plants, including callus age, type and callus pretreatment prior to plant regeneration induction. Manimaran et al. (2013) noticed that young calli had a higher tendency of callus development into whole plants compared with aged calli, while compact calli tended to develop into whole plants better than friable calli. It has also been well-documented that dehydration of calli prior to plant regeneration induction contributes to high regeneration capacit, because calli that lose water from the cells to become at quiescent stage during dehydration can re-absorb more water and nutrient when transferred to regeneration medium (Gray, 1987). By contrast, non-dehydrated calli absorb less nutrients from regeneration medium but release excess water from the cells to the surface of calli which restrains the regeneration process of the calli (Rueb et al., 1994). Several studies have been conducted to clarify that the frequency of callus induction and plant regeneration depends largely on genotypes and its interaction with culture conditions (Abe and Futsuhara, 1986; Mathias and Fukui, 1986; Tomes and Smith, 1985). The indica rice variety is one good example to support that genotypes and culture conditions determine the frequency of callus induction and plant regeneration, because it has been elucidated that the prolonged culturing of indica rice explants on induction medium results in callus browning and low frequency of subsequent regeneration (Manimaran et al., 2013).
The procedure for multiple shoots induction that allowed for high regeneration frequency, proposed here, is simple, efficient and reproducible and is expected to be useful in trials to produce rice plants with increased stress tolerance in vitro by avoiding problems associated with plant regeneration from calli following selection, as evidenced by earlier studies that the regeneration frequency from calli was quite low after salt treatment (Basu et al., 2002; Winicov, 1996; Zinnah et al., 2013).
CONCLUSIONS
In the present study, two different protocols for regeneration of rice employing single germinated seed as an explant source has been established, including (1) Multiple shoots induction and rooting and (2) Callus induction and plant regeneration. A great number of multiple shoots induced from germinated seed could be achieved on MS solid medium supplemented with 50 μM 6-BAP alone, whereas rooting of the obtained shoots was easily induced on hormone-free MS solid medium. For compact callus induction, the best result was obtained on MS solid medium amended with 2 mg L-1 2,4-D. A combination of 0.5 mg L-1 NAA and 2.0 mg L-1 6-BAP yielded the best shoot regeneration from compact calli. However, the regeneration frequency obtained via callus induction was inferior to that yielded through multiple shoots induction. The procedure reported here is expected to be useful in micropropagation and in vitro breeding and selection for drought and/or salt tolerance in rice for increasing the yield potential of this vital crop.
ACKNOWLEDGMENT
This study was financially supported by Ph.D. scholarships awarded to the first author from the Center of Excellence on Agricultural Biotechnology, Science and Technology Postgraduate Education and Research Development Office, Office of Higher Education Commission, Ministry of Education (AG-BIO/PERDO-CHE), the Agricultural Biotechnology Research Center for Sustainable Economy, Khon Kaen University and the Thailand Institute of Scientific and Technological Research (TISTR) and study funding from Khon Kaen University to the corresponding author and Research Group for improvement of salt-tolerant rice.
REFERENCES
- Abe, T. and Y. Futsuhara, 1986. Genotypic variability for callus formation and plant regeneration in rice (Oryza sativa L.). Theor. Applied Genet., 72: 3-10.
CrossRefDirect Link - Anzoua, K.G., K. Junichi, H. Toshihiro, I. Kazuto and J. Yutaka, 2010. Genetic improvements for high yield and low soil nitrogen tolerance in rice (Oryza sativa L.) under a cold environment. Field Crops Res., 116: 38-45.
CrossRef - Armstrong, C.L. and C.E. Green, 1985. Establishment and maintenance of friable, embryogenic maize callus and the involvement of L-proline. Planta, 164: 207-214.
CrossRefDirect Link - Bajji, M., S. Lutts and J.M. Kinet, 2000. Physiological changes after exposure to and recovery from polyethylene glycol-induced water deficit in callus cultures issued from durum wheat (Triticum durum Desf.) cultivars differing in drought resistance. J. Plant Physiol., 156: 75-83.
CrossRefDirect Link - Baskaran, P., B.R. Rajeswari and N. Jayabalan, 2006. Development of an in vitro regeneration system in sorghum (Sorghum bicolor (L.) Moench) using root transverse Thin Cell Layers (tTCLs). Turk. J. Bot., 30: 1-9.
Direct Link - Basu, S., G. Gangopadhyay and B.B. Mukherjee, 2002. Salt tolerance in rice in vitro: Implication of accumulation of Na+, K+ and proline. Plant Cell Tissue Org. Cult., 69: 55-64.
CrossRefDirect Link - Boling, A.A., B.A.M. Bouman, T.P. Tuong, Y. Konboon and D. Harnpichitvitaya, 2011. Yield gap analysis and the effect of nitrogen and water on photoperiod-sensitive Jasmine rice in North-East Thailand. NJAS-Wageningen J. Life Sci., 58: 11-19.
CrossRefDirect Link - Breitenbach, J., C. Bai, S.M. Rivera, R. Canela and T. Capell et al., 2014. A novel carotenoid, 4-keto-α-carotene, as an unexpected by-product during genetic engineering of carotenogenesis in rice callus. Phytochemistry, 98: 85-91.
CrossRefDirect Link - Bronsema, F.B.F., W.J.F. van Oostveen and A.A.M. van Lammeren, 2001. Influence of 2,4-D, TIBA and 3,5-D on the growth response of cultured maize embryos. Plant Cell Tissue Org. Cult., 65: 45-56.
CrossRefDirect Link - Che, P., T.M. Love, B.R. Frame, K. Wang, A. L. Carriquiry and S. H. Howell, 2006. Gene expression patterns during somatic embryo development and germination in maize Hi II callus cultures. Plant Mol. Biol., 62: 1-14.
CrossRefDirect Link - Ganeshan, S.V., S. Chodaparambil, M. Baga, D.B. Fowler, P. Hucl, B.G. Rossnagel and Chibbar, 2006. In vitro regeneration of cereals based on multiple shoot induction from mature embryos in response to thidiazuron. Plant Cell Tissue Org. Cult., 85: 63-73.
CrossRefDirect Link - Ge, X., Z. Chu, Y. Lin and S. Wang, 2006. A tissue culture system for different germplasms of indica rice. Plant Cell Rep., 25: 392-402.
CrossRefPubMedDirect Link - Joshi, R., A. Shukla and R. Sairam, 2011. In vitro screening of rice genotypes for drought tolerance using polyethylene glycol. Acta Physiol. Plant., 33: 2209-2217.
CrossRefDirect Link - Karthikeyan, A., S.K. Pandian and M. Ramesh, 2011. Agrobacterium-mediated transformation of leaf base derived callus tissues of popular indica rice (Oryza sativa L. sub sp. indica cv. ADT 43). Plant Sci., 181: 258-268.
CrossRefDirect Link - Khaleda, L. and M. Al-Forkan, 2006. Stimulatory effects of casein hydrolysate and proline in in vitro callus induction and plant regeneration from five deepwater rice (Oryza sativa L.). Biotechnology, 5: 379-384.
CrossRefDirect Link - Khanna, H.K. and S.K. Raina, 1999. Agrobacterium-mediated transformation of indica rice cultivars using binary and superbinary vectors. Aust. J. Plant Physiol., 26: 311-324.
Direct Link - Lee, K., H. Jeon and M. Kim, 2002. Optimization of a mature embryo-based in vitro culture system for high-frequency somatic embryogenic callus induction and plant regeneration from Japonica rice cultivars. Plant Cell Tissue Org. Cult., 71: 237-244.
CrossRefDirect Link - Lu, S., X. Peng, Z. Guo, G. Zhang and Z. Wang et al., 2007. In vitro selection of salinity tolerant variants from triploid bermudagrass (Cynodon transvaalensis × C. dactylon) and their physiological responses to salt and drought stress. Plant Cell Rep., 26: 1413-1420.
CrossRefDirect Link - Mahmood, I., A. Razzaq, M. Rasheed, A. Qayyum and M. Ahmad, 2014. Employment of immature embryo culture for in vitro selection of drought tolerant somaclones of wheat. Bulg. J. Agric. Sci., 20: 155-161.
Direct Link - Manimaran, P., G.R. Kumar, M.R. Reddy, S. Jain and T.B. Rao et al., 2013. Infection of early and young callus tissues of indica rice BPT 5204 enhances regeneration and transformation efficiency. Rice Sci., 20: 415-426.
CrossRefDirect Link - Matheka, J.M., E. Magiri, A.O. Rasha and J. Machuka, 2008. In vitro selection and characterization of drought tolerant somaclones of tropical maize (Zea mays L.). Biotechnology, 7: 641-650.
CrossRefDirect Link - Mathias, R.J. and K. Fukui, 1986. The effect of specific chromosome and cytoplasm substitutions on the tissue culture response of wheat (Triticum aestivum) callus. Theor. Applied Genet., 71: 797-800.
CrossRef - Mazid, M.S., M.Y. Rafii, M.M. Hanafi, H.A. Rahim, M. Shabanimofrad and M.A. Latif, 2013. Agro-morphological characterization and assessment of variability, heritability, genetic advance and divergence in bacterial blight resistant rice genotypes. South Afr. J. Bot., 86: 15-22.
CrossRefDirect Link - Mohanty, A., H. Kathuria, A. Ferjani, A. Sakamoto, P. Mohanty, N. Murata and A. Tyagi, 2002. Transgenics of an elite indica rice variety pusa basmati 1 harbouring the codA gene are highly tolerant to salt stress. Theoret. Applied Genet., 106: 51-57.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Nounjan, N., P.T. Nghia and P. Theerakulpisut, 2012. Exogenous proline and trehalose promote recovery of rice seedlings from salt-stress and differentially modulate antioxidant enzymes and expression of related genes. J. Plant Physiol., 169: 596-604.
CrossRef - Ntui, V.O., P. Azadi, H. Supaporn and M. Mii, 2010. Plant regeneration from stem segment-derived friable callus of fonio (Digitaria exilis (L.) Stapf.). Scient. Horticult., 125: 494-499.
CrossRefDirect Link - Raina, S.K., 1989. Tissue culture in rice improvement: Status and potential. Adv. Agron., 42: 339-398.
CrossRefDirect Link - Ramesh, M., V. Murugiah and A.K. Gupta, 2009. Efficient in vitro plant regeneration via leaf base segments of Indica rice (Oryza sativa L.). Indian J. Exp. Biol., 47: 68-74.
Direct Link - Rattana, K., P. Theerakulpisut and S. Bunnag, 2012. The effect of plant growth regulators and organic supplements on callus induction and plant regeneration in rice (Oryza sativa L.). Asian J. Plant Sci., 11: 182-189.
CrossRef - Rueb, S., M. Leneman, R.A. Schilperoort and L.A.M. Hensgens, 1994. Efficient plant regeneration through somatic embryogenesis from callus induced on mature rice embryos (Oryza sativa L.). Plant Cell Tissue Org. Cult., 36: 259-264.
CrossRefDirect Link - Sharma, V.K., R. Hansch, R.R. Mendel and J. Schulze, 2004. A highly efficient plant regeneration system through multiple shoot differentiation from commercial cultivars of barley (Hordeum vulgare L.) using meristematic shoot segments excised from germinated mature embryos. Plant Cell Rep., 23: 9-16.
CrossRef - Sharma, V.K., R. Hansch, R.R. Mendel and J. Schulze, 2005. Influence of Picloram and Thidiazuron on high frequency plant regeneration in elite cultivars of wheat with long-term retention of morphogenecity using meristematic shoot segments. Plant Breed., 124: 242-246.
CrossRefDirect Link - Soliman, H.I.A. and M.H. Hendawy, 2013. Selection for drought tolerance genotypes in durum wheat (Triticum durum Desf.) under in vitro conditions. Middle-East J. Sci. Res., 14: 69-78.
Direct Link - Thadavong, S., P. Sripichitt, W. Wongyai and P. Jompuk, 2002. Callus induction and plant regeneration from mature embryos of glutinous rice (Oryza sativa L.) cultivar TDK1. Agric. Nat. Resour., 36: 334-344.
Direct Link - Tomes, D.T. and O.S. Smith, 1985. The effect of parental genotype on initiation of embryogenic callus from elite maize (Zea mays L.) germplasm. Theor. Applied Genet., 70: 505-509.
CrossRef - Tsubo, M., S. Fukai, J. Basnayake and M. Ouk, 2009. Frequency of occurrence of various drought types and its impact on performance of photoperiod-sensitive and insensitive rice genotypes in rainfed lowland conditions in Cambodia. Field Crops Res., 113: 287-296.
CrossRefDirect Link - Verma, D., M.W. Ansari, G.K. Agrawal, R. Rakwal, A. Shukla and N. Tuteja, 2013. In vitro selection and field responses of somaclonal variant plants of rice cv PR113 for drought tolerance. Plant Signal. Behav., 8: 1-5.
Direct Link - Winicov, I., 1996. Characterization of rice (Oryza sativa L.) plants regenerated from salt-tolerant cell lines. Plant Sci., 113: 105-111.
CrossRefDirect Link - Wu, W., L. Nie, Y. Liao, F. Shah and K. Cui et al., 2013. Toward yield improvement of early-season rice: Other options under double rice-cropping system in central China. Eur. J. Agron., 45: 75-86.
CrossRef - Yuan, X., Z. Wang, J. Liu and J. She, 2009. Development of a plant regeneration system from seed-derived calluses of centipedegrass [Eremochloa ophiuroides (Munro.) Hack]. Scient. Horticult., 120: 96-100.
CrossRefDirect Link - Zhang, S., R. Williams-Carrier and P.G. Lemaux, 2002. Transformation of recalcitrant maize elite inbreds using in vitro shoot meristematic cultures induced from germinated seedlings. Plant Cell Rep., 21: 263-270.
Direct Link - Zhang, S., H. Zhang and M.B. Zhang, 1996. Production of multiple shoots from shoot apical meristems of oat (Avena sativa L.). J. Plant Physiol., 148: 667-671.
CrossRefDirect Link - Zhao, F., Q. Li, M. Weng, X. Wang and B. Guo et al., 2013. Cloning of TPS gene from eelgrass species Zostera marina and its functional identification by genetic transformation in rice. Gene, 531: 205-211.
CrossRefDirect Link