
H. Zakikhani
University Putra Malaysia, Malaysia
Y.M. Khanif
University Putra Malaysia, Malaysia
A.R. Anuar
University Putra Malaysia, Malaysia
O. Radziah
University Putra Malaysia, Malaysia
A. Soltangheisi
University Putra Malaysia, Malaysia
Asian Journal of Crop Science
Year: 2014 | Volume: 6 | Issue: 3 | Page No.: 236-244







ABSTRACT
A hydroponic trial was carried out to determine the effect of molybdenum (Mo) on utilization and uptake of macro and micronutrients in different rice cultivars. The experiment was conducted using a randomized complete-block design, with a split-plot arrangement of treatments and three replications. Four rates of Mo (0, 0.01, 0.1 and 1 mg L-1) and five cultivars (MR219, HASHEMI, MR232, FAJRE and MR253) provided the main and sub-plots, respectively. The results showed that the enhancement of Mo levels (from 0.01 to 0.1 and 0.1 to 1mg L-1) led to increase of root Mo uptake by 81.7 and 61.6% and shoot Mo uptake by 43 and 87%, respectively. Also, Mo application significantly affected shoot Phosphorus uptake so that highest shoot Phosphorus (P) uptake (0.43% plant-1) was achieved at 1mg Mo L-1 but there was no significant influence on root P uptake. Shoot Iron (Fe) uptake was inversely proportional to increment of Mo rates therefore, the highest rate of shoot Fe (59.05 μg plant-1) was obtained in treatments with least amount of applied Mo. Also, increase in Mo application enhanced manganese (Mn) uptake of shoot but there was no significant effect on Mn root uptake.
PDF Abstract XML References Citation
Received: January 13, 2014;
Accepted: February 25, 2014;
Published: April 14, 2014
How to cite this article
H. Zakikhani, Y.M. Khanif, A.R. Anuar, O. Radziah and A. Soltangheisi, 2014. Effects of Different Levels of Molybdenum on Uptake of Nutrients in Rice Cultivars. Asian Journal of Crop Science, 6: 236-244.
DOI: 10.3923/ajcs.2014.236.244
URL: https://scialert.net/abstract/?doi=ajcs.2014.236.244
DOI: 10.3923/ajcs.2014.236.244
URL: https://scialert.net/abstract/?doi=ajcs.2014.236.244
INTRODUCTION
Rice is one of the main crop worldwide which is a staple food of nearly half of the world’s population. A great portion of rice is produced in Asia. Importance of Molybdenum (Mo) as a trace element in plant and animal growth is of critical importance (Mengel et al., 2001). Deficiency of molybdenum causes tasseling disruption, flowering delay and anthesis inhibition (Agarwala, 1978) and could lead to lower dormancy levels of wheat (Cairns and Kritzinger, 1992). Mo exists in soil in different oxidation states from II to VI, where VI is predominant. Available form of Mo to plants is MoO4-2 which is mobile in plants. A numerous factors affect the availability and uptake of Mo to plants including soil pH, drainage, soil organic matter, plant species and parts, soil texture and type and oxides presence (Gupta, 1997). Within the cells, Mo anions incorporated by complexes into cofactors (Schwarz, 2005). Molybdenum cofactors (Moco) also bind to molybdoenzymes including nitrate reductase, nitrogenase, xanthine dehydrogenase, aldehyde oxidase and sulfite oxidase (Schwarz et al., 2009; Mendel and Hansch, 2002). It has shown that plant uptake of Mo could be directly proportional with enhancement of Phosphorus (P) and can be declined by presence of available Sulfur (S) (Mortvedt and Cunningham, 1971). High availability of P and Mo ions can be due to ligand exchange mechanism and then hydroxyl ion increase (Barrow et al., 2005). Enhancement of soil potassium (K) concentration could decline plant Mo content (Villora et al., 2003) and an antagonistic influence of Mo on Fe nutrition of different plants has been observed (Berry and Reisenauer, 1967). In a culture study, enhancement of Mo at lower levels of Fe declined the yield of tomato but at higher Fe levels, addition of Mo raised the yield (Kirsch et al., 1960). Other nutrients such as Copper (Cu) and Manganese (Mn) also have antagonistic impacts on plant Mo uptake (Jackson, 1967; Nayyar et al., 1980). Also, at both lower amount of Mn and Fe, addition of Mo had not had any effect on uptake of Mn but at higher amounts of Fe and Mn, Mo enhanced the uptake of Mn. (Kirsch et al., 1960). The main objective of this study were 1- to determine the effects of Mo on rice yield and uptake of other nutrients 2- to screen for highly Mo efficient variety.
MATERIALS AND METHODS
Five rice (Oryza sativa) cultivars (V1: MR219, V2: HASHEMI, V3: MR232, V4: FAJRE and V5: MR253) were used and soaked in tap water for 24 h, then were placed between two damp filter paper within petri dishes. The dishes placed in oven at 25°C for 3 days. Germinated seed were selected and transplanted to plastic trays (36x26x8) containing nutrient solution at the rate of 25 seedlings per tray. The Yoshida et al. (1976) nutrient solution was used as solution medium. Molybdenum (Mo) combination was added to nutrient solution as ammonium molybdate [(NH4)6Mo7O24•4H2O] at four rates: [M0: 0, M1: 0.01, M2: 0.1 and M3: 1.0 mg L-1]. The pH of nutrient solution was monitored to maintain it between 5.5 and 5.8 using 0.1 M HCl solution or 1.0 M KOH. The nutrient solution was changed regularly at 3 days intervals. The experiment was a Split Plot Design (SPD) in Randomized Complete Block Design (RCBD) using three replications. Applied molybdenum and varieties were considered as main and subplots, respectively.
Plants were taken from the treatments at 35th day after transplantation, washed with deionized water and oven dried at 70°C for 48 h. Then, separated as roots and shoots for micro and macro nutrients analysis and dry matter weights (DM) determination. Also, roots were washed in distilled water several times before scanning procedure. Root length, volume and diameter were determined using WinRhizo (STD 4800, Regent, Canada). Evaluation of plant height and number of tillers were manually done. Dried samples weighed, placed in glass tubes and concentrated HNO3 was added. In order to increase the speed of reaction concentrated H2SO4 added (Jones and Case, 1990). Glass tubes placed on hot plate, heated at 125°C up to 4 h and allowed to cool. To maintain wet digestion and clean digested tissues, 30% H2O2 added.
Mo was determined by inductively coupled plasma optical emission spectrometry (ICP-OES) (Optima 8300, PerkinElmer, USA) and Phosphorus (P) was measured by autoanalyzer (8000 series, Lachat QuickChem FIA+, USA).
All data were statistically analyzed using Excel 2010 and ANOVA procedure in SAS 9.2 (SAS institute, Cary, NC, USA). Also, Interactions and main effects of factors were analyzed following the principle of F statistics.
RESULTS AND DISCUSSION
Molybdenum: Enhancement of shoot Mo uptake was observed when Mo rates increased from 0.01 to 0.1 and 0.1 to 1 mg L-1 by 47.7 and 83.3%, respectively. Also, the enhancement of Mo levels (from 0.01 to 0.1 and 0.1 to 1 mg L-1) led to increase of root Mo uptake by 81.7 and 61.6%, respectively. According to results, uptake of Molybdenum in root M1 and M2 were higher than shoot M1 and M2 levels but the uptake of molybdenum at M3 levels was higher in shoots than roots. At level of M1 and M2, highest amount of root Mo uptake was seen in cultivar V4 but when Mo level increased (M3) in the medium, the maximum root Mo uptake was achieved in V3 cultivar (Fig. 2).
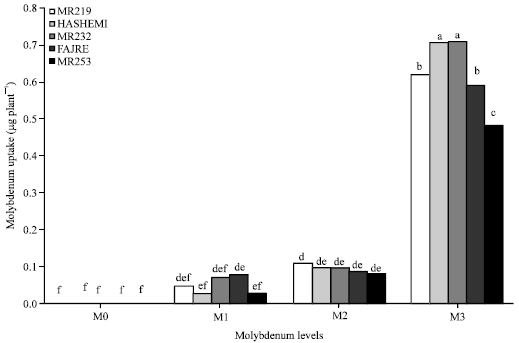 | |
| Fig. 1: | Interaction of molybdenum and varieties on shoot molybdenum uptake |
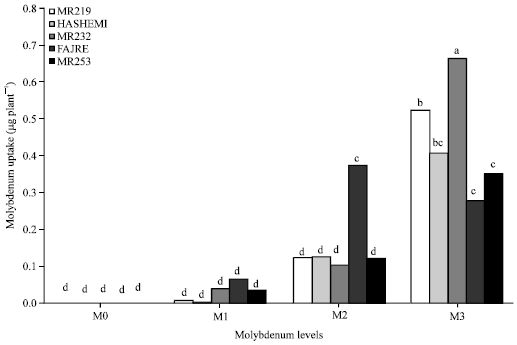 | |
| Fig. 2: | Interaction of molybdenum and varieties on root molybdenum uptake |
Enhancement of shoot molybdenum uptake in the cultivars at levels of M1 and M2 was low but it drastically increased when Mo application rate reached maximum (Fig. 1). The interaction of Molybdenum (Mo) and Varieties (V) on Mo uptake in shoots and roots were significant (p≤0.01) so that highest molybdenum uptake in shoots (0.708 μg plant-1) and roots (0.669 μg plant-1) were observed in MoV3 treatment (Table 1). This might account for the high efficiency in cultivar V3. Also, binding sites of Mo might be controlled genetically, which needed to be studied further. Mandai et al. (1998) stated that application of Mo increased uptake of Mo in lentil plants grown in soil and Moore and Patrick (1991) reported that Mo contents of rice leaves were positively correlated with activities of molybdate ions.
| Table 1: | Analysis of variance for the effects of molybdenum applications and varieties on shoot and root Mo uptake, shoot P, Fe and Mn uptake and root volume |
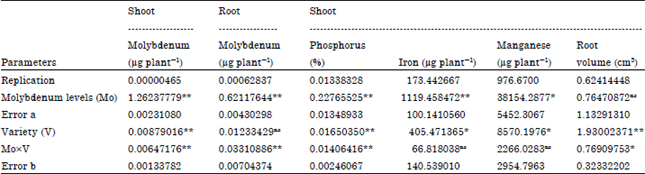 | |
| ns: Non-significance, *Significant at the 0.05 probability levels, **Significant at the 0.01 probability levels | |
Also, other results revealed that concentration of Mo in clover tissues enhanced when application of Mo increased at different cutting stages (Nayyar et al., 1980). Our results showed that an increase in each level of applied Mo led to enhancement of Mo uptake so that the highest shoot Mo uptake reached maximum at 1 mg L-1 Mo application. This might be due to influence of prevalence of phosphorus in solution which is in agreement with the results of Stout and Meagher (1948). In contrast, Weng et al. (2009) presented that an ascending trend in accumulation of Mo content in round leaf cassia plants was seen with increasing the Mo application (up to 2 mg kg-1) but plant Mo content at higher amounts (4 mg kg-1) were lower than that at 2 mg kg-1. This diminution could be due to the process of active adsorption of Mo. Also, Stout et al. (1951) reported that P accounts for Mo release from the root cells into system of translocation.
Copper, zinc: The effects of Mo rates on the shoot and root zinc uptake were not significant. Also, the effects of varieties on shoot zinc uptake were not significant but the effect of varieties on root zinc uptake was significant (p≤0.01) (Table 1). The results showed that the highest and lowest root zinc uptake were observed in V1 (7.66 μg plant-1) and V5 (2.38 μg plant-1), respectively. The effects of Mo on the shoot and root copper uptake rate were non-significant but the effects of varieties on copper uptake in shoot and root were significant. The results showed that V1 and V5 had the highest and lowest copper uptake in shoots and roots, respectively.
Iron: The effects of Mo and V on shoot iron uptake were significant (Table 1) but interaction of Mo and V were non-significant. The results showed that highest rate of shoot Fe (59.05 μg plant-1) was seen in treatment containing lowest amount of applied Mo and enhancement of Mo application did not lead to increase of Fe uptake. It had shown that Fe and Mo uptake are inversely proportional so that addition of Mo resulted in enhanced Fe-chlorosis and tomato yield diminution (Gerloff et al., 1959) or application of higher levels of Mo in growth solution adversely affected translocation of iron from red clover roots to shoots (Hangar, 1965). Because of strong adsorption of molybdate by iron oxides and decline of its surface activity to water, uptake of Fe could be depressed. Thus, immobilization of a considerable amount of Fe due to adsorption of minute amounts of Mo can be accounted for the Fe uptake depression even at low levels of Mo. According to the results, lowest rate of shoot iron uptake (39.93 μg plant-1) was observed in treatments contained highest amount of applied Mo. Highest rate of iron in shoot was seen in V4 and application of Mo and V did not affect root Fe uptake. According to results, the main effects of Mo and variety on root Fe uptake were not significant.
Manganese: The effects of Mo application and variety on uptake of Mn in shoots were significant (p≤0.01) (Table 1). According to the results, an ascending trend of Mn uptake in shoots was found when amount of applied Mo increased. The results showed that V2 had taken up Mn at the highest point (183.13 μg plant-1) and the lowest Mn content (110.79 μg plant-1) was achieved in V1. The effects of Mo rates on Mn uptake were non-significant in plant roots but the effects of varieties on root Mn uptake were significant (p≤0.01). The results showed that the highest and lowest Mn uptake rates were seen in V2 (44.5 μg plant-1) and V5 (19.2 μg plant-1), respectively. In contrast to current results, uptake of Mo by cauliflower declined when Mn applied into acidic soil. Mulder (1954) and Sims et al. (1975), reported that Mn halted uptake of Mo in burley tobacco grown in soil solution.
Nitrogen and potassium: The effects of Mo rates on percentage of shoot nitrogen (N) and potassium (K) were not significant but the effects of Variety on percentage of shoot N and k were significant .The results also showed that the effects of Mo levels and varieties on percentage of root N and K were not significant either. According to results, the highest percentage of shoot N (3.67% plant-1) and K (3.78% plant-1) was seen in V5.
Phosphorus: The results showed that increase in percentage of shoot Phosphorus (P) was achieved when Mo rates enhanced. The highest percentage of P (0.43% plant-1) was seen in the maximum rate of applied Mo and lowest (0.18% plant-1) was observed in treatments without applied Mo (Fig. 5). Basak et al. (1982) represented that application of Mo enhanced P concentration in rice plants and uptake of Mo increased by application of P in solution culture (Mortvedt and Cunningham, 1971). These reports show that synergistic effects exist between Mo and P application so that formation of anionic complexes between P and Mo, which are more likely to be absorbed by plant roots, could be accounted for such effects. In contrast, Chatterjee et al. (1985) reported that application of Mo decreased total amount of P in mustard due to hindering of P enzyme activity such as phosphatase. According to the results, enhancement of applied Mo increased shoot P contents at each level of Mo in all varieties (Fig. 3). This is in agreement with those of Liu et al. (2010) who found higher amount of Mo in solution culture of canola increased P effectiveness. The interaction of Mo and V on percentage of shoot P was significant (p≤0.01) so that highest percentage of P (0.6% plant-1) was seen in Mo3V3 treatment (Table 1). This implies that responses of varieties to absorb P during application of various levels of Mo are different. According to results the main effects of Mo and V and their interactions on root percentage of P were not significant. While levels of Mo increased, the trends of P content enhanced in rice cultivars so that trends of enhancement were observed in the order of V3, 5, 4, 1, 2. According to results, the main effects of Mo and V and their interactions on root P concentration were not significant.
Root length, volume and diameter: Root volume was significantly affected by interaction of Mo and V (p≤0.05). The root volume of V3 at the high level of molybdenum (M2) was 3.02 cm3 (Fig. 4). According to results, the higher amounts of applied Mo greatly increased root volume of V3 in comparison with other varieties. The results also showed that Mo levels did not have effect on root length and root average diameter.
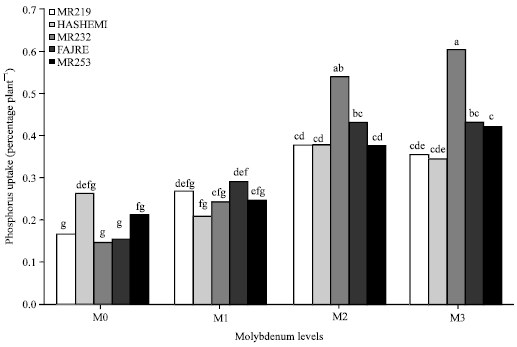 | |
| Fig. 3: | Interaction of molybdenum and varieties on shoot phosphorus uptake |
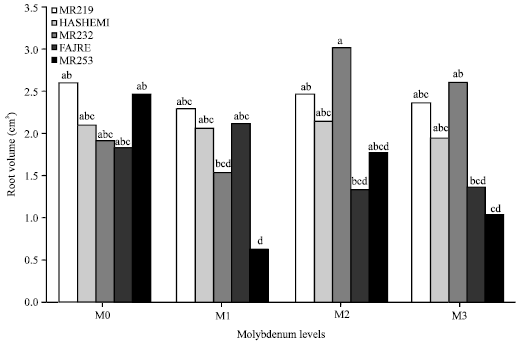 | |
| Fig. 4: | Interaction of molybdenum and varieties on root volume |
Variety also did not have effect on root average diameter but its effect on root length was significant (p≤0.01). The results showed that V1 had the highest root length comparing with the other varieties.
Shoot and root dry weight and plant height and tiller: Molybdenum rates didn’t affect shoot and root dry weights, plant heights and tiller numbers but the effects of variety on shoot and root dry weights, plant heights and tiller numbers were significant (p≤0.01).
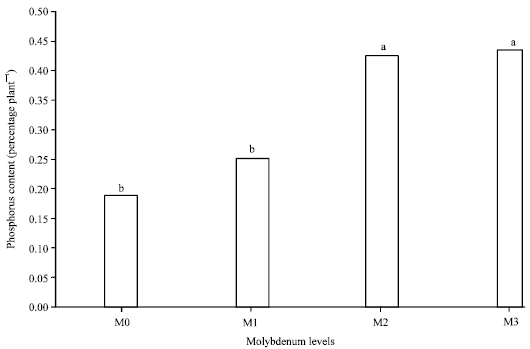 | |
| Fig. 5: | Effects of molybdenum on shoot phosphorus content |
The highest shoot and root dry weights were seen in V3 (0.602 g) and V1 (0.168 g), respectively. Also, the maximum and minimum plant heights were observed in V2 (55.34 cm) and V5 (44.89 cm), respectively. According to results, the highest and lowest number of tillers were seen in V3 (4.3) and V2 (2.09), respectively.
ACKNOWLEDGMENT
This research was supported by Long Term Research Grant Scheme (LRGS) from Ministry of Education and Faculty of Agriculture, Department of Land management, UPM.
REFERENCES
- Agarwala, S., C.P. Sharma, S. Farooq and C. Chatterjee, 1978. Effect of molybdenum deficiency on the growth and metabolism of corn plants raised in sand culture. Can. J. Bat., 56: 1905-1908.
CrossRefDirect Link - Barrow, N.J., P. Cartes and M.L. Mora, 2005. Modifications to the Freundlich equation to describe anion sorption over a large range and to describe competition between pairs of ions. Eur. J. Soil Sci., 56: 601-606.
CrossRef - Basak, A., L.N. Mandal and M. Haldar, 1982. Interaction of phosphorus and molybdenum in relation to uptake and utilization of molybdenum, phosphorus, zinc, copper and manganese by rice. Plant Soil, 68: 261-269.
CrossRef - Berry, J.A. and H.M. Reisenauer, 1967. The influence of molybdenum on iron nutrition of tomato. Plant Soil, 27: 303-313.
CrossRef - Cairns, A.L.P. and J.H. Kritzinger, 1992. The effect of molybdenum on seed dormancy in wheat. Plant Soil, 145: 295-297.
CrossRef - Chatterjee, C., N. Nautiyal and S.C. Agarwala, 1985. Metabolic changes in mustard plants associated with molybdenum deficiency. New Phytol., 100: 511-518.
CrossRefDirect Link - Kirsch, R.K., M.E. Harward and R.G. Petersen, 1960. Interrelationships among iron, manganese and molybdenum in the growth and nutrition of tomatoes grown in culture solution. Plant Soil, 12: 259-275.
CrossRef - Liu, H., C. Hu, X. Hu, Z. Nie, X. Sun, Q. Tan, and H. Hu, 2010. Interaction of molybdenum and phosphorus supply on uptake and translocation of phosphorus and molybdenum by brassica napus. J. Plant Nutr., 33: 1751-1760.
CrossRef - Mandai, B., S. Pal and L.N. Mandai, 1998. Effect of molybdenum, phosphorus and lime application to acid soils on dry matter yield and molybdenum nutrition of lentil. J. Plant Nutr., 21: 139-147.
CrossRef - Moore Jr., P.A. and W.H. Patrick Jr., 1991. Aluminium, boron and molybdenum availability and uptake by rice in acid sulfate soils. Plant Soil, 136: 171-181.
CrossRef - Mulder, E.G., 1954. Molybdenum in relation to growth of higher plants and micro-organisms. Plant Soil, 5: 368-415.
CrossRefDirect Link - Schwarz, G., R.R. Mendel and M.W. Ribbe, 2009. Molybdenum cofactors, enzymes and pathways. Nature, 460: 839-847.
PubMed - Stout, P.R., W.R. Meagher, G.A. Pearson and C.M. Johnson, 1951. Molybdenum nutrition of crop plants. I. The influence of phosphate and sulfate on the absorption of molybdenum from soils and solution cultures. Plant Soil, 3: 51-87.
CrossRefDirect Link - Villora, G., D.A. Moreno and L. Romero, 2003. Potassium supply influences molybdenum, nitrate and nitrate reductase activity in eggplant. J. Plant Nutr., 26: 659-669.
CrossRef - Weng, B.Q., D.F. Huang, D.Z. Xiong, Y.X. Wang, T. Luo, Z.Y. Ying and H.P. Wang, 2009. Effects of molybdenum application on plant growth, molybdoenzyme activity and mesophyll cell ultrastructure of round leaf cassia in red soil. J. Plant Nutr., 32: 1941-1955.
CrossRef - Yoshida, S., D.A. Forno, J.H. Cock and K.A. Gomez, 1976. Laboratory Manual for Physiological Studies of Rice. 3rd Edn., Internatioanl Rice Research Institute, Los Banos, Philippines, Pages: 83.
Direct Link