
M.A. Abdel-Moneam
Department of Agron, Faculty of Agriculture, Mansoura University, Egypt
M.S. Sultan
Department of Agron, Faculty of Agriculture, Mansoura University, Egypt
S.M.G. Salama
Central Lab. for Design and Statistical Analysis Res. ARC, Egypt
A.M. El Oraby
Fine Seeds International Company, Egypt
Asian Journal of Crop Science
Year: 2014 | Volume: 6 | Issue: 2 | Page No.: 142-149







ABSTRACT
A half diallel cross among 5 inbred lines of maize was evaluated under two different nitrogen rates for yield and yield components characters i.e., ears No. plant-1, ear length, ear diameter, 100-kernel weight, grain yield plant-1 and shelling percentage to evaluate the role of GCA and SCA of inbred lines in hybrids performance under normal and stress of nitrogen levels and to establish the magnitude of heterosis. Mean squares of genotypes were highly significant for all studied traits under both nitrogen levels, except shelling percentage under normal nitrogen level. General and specific combining ability (GCA and SCA) mean squares were significant or highly significant for all studied traits, except shelling percentage under normal nitrogen level at GCA and SCA. GCA/SCA ratios revealed that the non-additive gene action for all studied traits under both nitrogen levels was detected. The best combiners were P4 (Inb.204) and P5 (Inb.213) for most of studied traits under normal and stress nitrogen levels. This result indicated that these inbred lines could be considered as good combiners for improving these traits. The best crosses for ears No. plant-1 were P1 (Inb.84) xP3 (Inb.144) and P3 (Inb.144) xP4 (Inb.204), for ear length was P2 (Inb.95) xP4 (Inb.204), for ear diameter was P1 (Inb.84) xP5 (Inb.213), for 100-kernel weight was P2 (Inb.95) xP4 (Inb.204) and for grain yield plant-1 was P1 (Inb.84) xP3 (Inb.144) under both nitrogen levels. These crosses could be selected and used in breeding programs for improving these traits. Results showed significant or highly significant heterosis over mid or better parents for all studied traits, except shelling percentage under normal nitrogen level. The best crosses over both their mid-parents and better-parent for ears No. plant-1 was P3 (Inb.144) xP4 (Inb.204), for ear length was P2 (Inb.95) xP4 (Inb.204), for ear diameter was P1 (Inb.84) xP5 (Inb.213), for 100-kernel weight was P2 (Inb.95) xP4 (Inb.204), for grain yield plant-1 was P1 (Inb.84) xP3 (Inb.144) under both nitrogen levels and for shelling percentage was P1 (Inb.84) xP3 (Inb.144) under stress nitrogen level.
PDF Abstract XML References Citation
Received: December 20, 2013;
Accepted: January 10, 2014;
Published: March 08, 2014
How to cite this article
M.A. Abdel-Moneam, M.S. Sultan, S.M.G. Salama and A.M. El Oraby, 2014. Evaluation of Combining Ability and Heterosis for Yield and its Components Traits of Five Maize Inbreds under Normal and Stress Nitrogen Fertilization. Asian Journal of Crop Science, 6: 142-149.
DOI: 10.3923/ajcs.2014.142.149
URL: https://scialert.net/abstract/?doi=ajcs.2014.142.149
DOI: 10.3923/ajcs.2014.142.149
URL: https://scialert.net/abstract/?doi=ajcs.2014.142.149
INTRODUCTION
Based on area and production, maize is the 3rd most important cereal crop after wheat and rice, all over the world for production and consumption. In addition to its use as a human food, it is also utilized as a poultry and livestock feed. Moreover, it is also used for many industrial purposes. Nitrogen is the most important nutritive element for the worldwide production of cereals (wheat, barley, rice, sorghum and maize). It is mostly supplied to the soil in the form of inorganic fertilizers and to a lesser extent as organic manure. A considerable portion of N-fertilizer is lost through gaseous plant emissions, soil denitrification, surface runoff, ammonia volatilization and leaching (Akintoye et al., 1999; Raun and Johnson, 1999).
The affordability of N in the developed countries has led to its misuse and over application (Raun and Johnson, 1999) and created growing environmental concerns from increased nitrate leaching that may lead to ground water contamination. In contrast, the rates of N fertilizers in most developing countries are considerably low because of the limited access to fertilizers and the low purchasing power of small farmers due a high fertilizer/cereal grain price ratio. El-Badawy (2013) found that nitrogen rates, genotype, parents, hybrids and parents vs. crosses mean squares were significant for all traits. Sallah et al. (1996) found that genotypic variance was greater at high N for grain yield and at low N for the other traits.
The two main genetic parameters of diallel analysis are GCA and SCA which are essential in developing breeding strategies. El-Absawy (2002) cleared that GCA mean squares were significant for grain yield per plant-1, ear diameter and 100-grain weight and also added that the mean squares of SCA were significant for ear diameter and 100-grain weight. In this concern, several investigators reported that additive gene action was responsible for the inheritance of growth characters (Sedhom, 1994; Ahmed et al., 2000; Al-Naggar et al., 2002; Alamnie et al., 2006; El-Badawy, 2006). However, Dadheech and Joshi (2007), Barakat and Osman (2008) and Irshad-El-Haq et al. (2010) reported that non-additive gene action was more important in the inheritance of grain yield and most other agronomic traits in maize.
Therefore, the objectives of this study were to evaluate the role of GCA and SCA of inbred lines in hybrids performance and to establish the magnitude of heterosis under high and low nitrogen fertilization rates.
MATERIALS AND METHODS
Plant materials: Five inbred lines of corn (Zea mays L.) were used as parents in this study i.e., P1(Inb. 84), P2(Inb95), P3(Inb. 144), P4(Inb. 204) and P5(Inb. 213). These inbred lines were obtained from Agriculture Research Center (ARC), Egypt.
Field experiments: The field trials were started in the 2011 growing season in the Experimental Farm of the Faculty of Agric., Mansoura Univ. and lasted in 2012. In 2011 growing season, the five parental inbred lines were planted on April 21st and May 7th and each inbred line was grown in two rows, to overcome the differences in flowering date and to secure enough hybrid seeds. During this season, all possible cross combinations, without reciprocals, were made giving a total of 10 F1’s hybrid seeds. In 2012 growing season, 16 entries (10 F1’s along with their 5 parental inbred lines plus one check cultivar (S.C.10)) were grown in two experiments representing two different nitrogen levels, which were 60 kg N fad-1 (stress) and 120 kg N fad-1 (normal) by using distance of 70 cm between ridges and 25 cm between hills. Each experiment was designated in a Randomized Complete Blocks Design (RCBD) with three replicates. Each plot consisted of one ridge three meters long. Hills were thinned after seedling emergence to secure one plant per hill. Each experiment was hoed twice, before first and second irrigations. Phosphorus in the form of calcium super phosphate (15.5% P2O5) at a rate of 200 kg fad-1, was added to the soil during seedbed preparation and potassium sulphate (48% K2O) at a level of 50 kg fad-1 was applied after thinning. Moreover, nitrogen in the form of Urea (46% N) at a rate studied (60 and 120 kg N fad-1) was added in two equal split doses, before the first and the second irrigation. Other agriculture practices were applied as recommended.
Studied traits: The following measurements were recorded: The ears No. plant-1, ear length (cm), ear diameter (cm), 100-kernel weight (g), grain yield plant-1 (g) and shelling percentage.
Statistical analysis: The obtained data were statistically analyzed for analysis of variance by using computer statistical program MSTAT-C. The Ten single crosses comprise a half diallel among 5 inbred parents. Data of all 10 single crosses at each nitrogen level treatment were analyzed as randomized blocks. The sum squares of genotypes were partitioned to general and specific combining ability, following method 2 model 1 (fixed) of Griffing (1956).
RESULTS AND DISCUSSION
Results in Table 1 indicated that mean square of genotypes were highly significant for all studied traits under both nitrogen levels, except shelling percentage under normal nitrogen level. Mean squares of General Combining Ability (GCA) were significant or highly significant for all studied traits under both nitrogen levels, except shelling percentage under normal nitrogen level which was non-significant. Mean squares of Specific Combining Ability (SCA) were highly significant for all studied traits under both nitrogen levels, except shelling percentage under normal nitrogen level. The GCA/SCA ratio was less than unity for all studied traits, this means that these traits are predominantly controlled by non-additive gene action under both nitrogen levels, as shown in Table 1. Similar results were obtained by El-Hosary et al. (1994), Hoballah and Radwan (1996), Gado et al. (2000), Abdel-Moneam et al. (2009) and El-Badawy (2013).
Mean performance of traits: Means of ears No. plant-1 as affected by normal and stress nitrogen levels are presented in Table 2. The better parent in ears No. plant-1 was P2(Inb.95) under both nitrogen levels. The better cross in ears No. plant-1 was p3xp4 under both nitrogen levels. The better parent in ear length was P5 (Inb.213) under both nitrogen levels. The better cross in ear length was check S.C.10 under both nitrogen levels. The better parent in ear diameter was P2(Inb.95) under both nitrogen levels. The better cross in ear diameter was P1xP5 under both nitrogen levels. The better parent in 100-kernel weight was P1(Inb.84) under both nitrogen levels. The better cross in 100-kernel weight was check S.C.10 under both nitrogen levels. The better parent in grain yield plant-1 was P4(Inb.204) under both nitrogen levels.
| Table 1: | Mean squares of genotypes, GCA and SCA for studied maize yield and yield components traits under normal and stress nitrogen levels conditions |
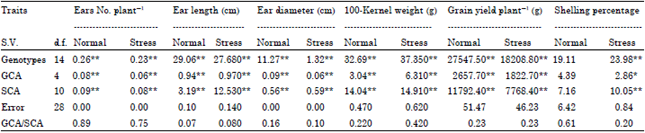 | |
| *and**significant at 5 and 1% probability levels, respectively | |
| Table 2: | Means of studied traits for maize inbreds and their F1 crosses under normal and stress nitrogen levels conditions |
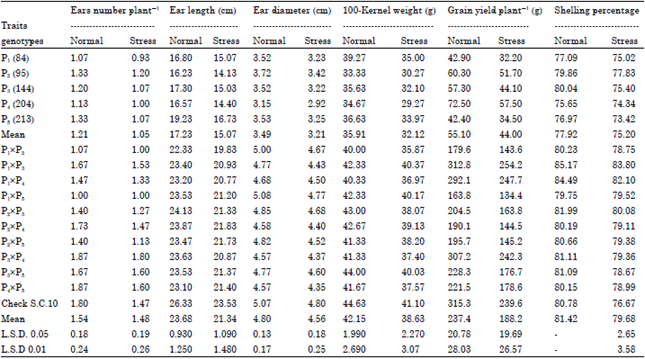 | |
| Table 3: | Estimates of G.C.A. effects of five parents of maize genotypes for all studied yield and yield components traits |
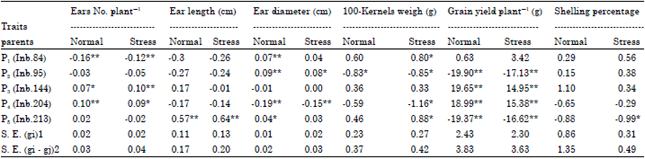 | |
| *, **Significant at 0.05 and 0.01 level of probability, respectively, S.E.(gi), standard error for an GCA effects, S.E.(gi-gj), standard error for the difference between two estimates of GCA effects | |
The better cross in grain yield plant-1 was P1xP3 under stress nitrogen level and cross check S.C.10 under normal nitrogen level. It may indicated that grain yield plant-1 is greatly influenced by different agronomic treatments. Shelling percentage ranged from 73.42 to 77.83% under stress nitrogen level. The highest value was recorded by P2(Inb.95) under stress nitrogen levels. Shelling percentage ranged from 76.67 to 83.80% under stress nitrogen level. The highest value was recorded by P1xP3 under stress nitrogen level. These results are in agreement with findings of Abdel-Moneam (2000), Soares et al. (2011) and El-Badawy (2013).
General combining ability: Based on GCA estimates, it could be concluded that the best combiners for ears No. plant-1 were inbred lines P3 (Inb144) and P4(Inb.204); for ear length was P5(Inb.213); for ear diameter was P2(Inb.95); for 100-kernel weight was P5(Inb.213) and for grain yield plant-1 was P3(Inb.144) and P4 (Inb. 204) under both nitrogen levels, (Table 3). These results indicated that these inbred lines could be considered as good combiners for improving these traits.
| Table 4: | Estimates of S.C.A. effects of 10 single crosses maize for yield and yield component traits under normal and stress nitrogen levels |
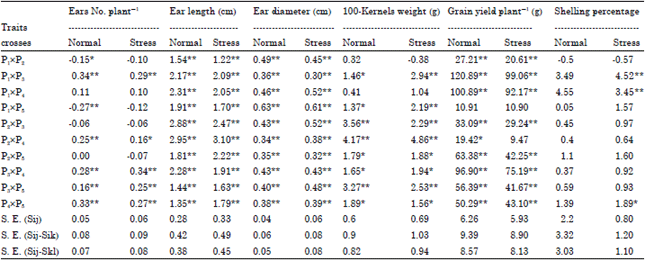 | |
| *, **Significant at 0.05 and 0.01 level of probability, respectively, SE(Sij), standard error for an SCA effects, SE(Sij-Sik), standard error for the difference between two SCA effects for a common parent, SE(Sij-Skl), standard error for the difference between two SCA effects for a non-common parent | |
Specific combining ability effects (Sij): Significant SCA effects were found in all studied traits for most crosses under both nitrogen levels (Table 4). Based on SCA effects, it could be concluded that under both nitrogen levels significant or highly significant positive SCA effects for ears No. plant-1 was recorded with P1xP3, P2xP4, P3xP4, P3xP5 and P4xP5; for ear length and ear diameter were all studied crosses; for 100-kernel weight was P1xP3, P1xP5, P2xP3, P2xP4, P2xP5, P3xP4, P3xP5 and P4xP5; for grain yield plant-1 was all studied crosses, except P1xP5 under both nitrogen levels and P2xP4 under stress nitrogen level and for shelling percentage was P1xP3, P1xP4 and P4xP5 under stress nitrogen level. These crosses could be selected and used in hybridization programs for improving these traits.
Heterosis over mid-parents and better-parent: Results showed that the highest positive and significant heterosis over mid-parents was demonstrated by cross P3xP4 (60.5 and 73.9%) followed by P4x5 (52 and 54.6%) under both normal and stress nitrogen levels, respectively. For better parent of ears No. plant-1, cross P3xP4 (55.8 and 68.2%) followed by P4xP5 (40.6 and 49.5%) showed maximum positive and significant heterosis under both nitrogen levels, respectively. The highest positive significant heterosis over mid-parents were recorded by P2xP4 (45.5%) followed by P2xP3 (43.9%) under normal nitrogen level, whereas under stress nitrogen level, cross P2xP4 (53%) gave maximum over mid-parents for ear length followed by cross P2xP3 (46.3%). For better-parent of ear length under normal nitrogen level, cross P2xP4 (44.1%) showed maximum positive and significant heterosis followed by P2xP3 (39.5%), whereas under stress nitrogen level, maximum positive and highly significant heterosis of better-parent were showed by crosses P2xP4 (51.6%) followed by P2xP3 (41.9%). The highest positive significant heterosis over mid-parents was recorded by P1xP5 (44.1%) followed by P1xP4 (40.3%) under normal nitrogen level; whereas under stress nitrogen level, cross P1xP5 (47.2%) gave maximum over mid-parents for ear diameter followed by crossP1xP4 (46.3%). For better-parent of ear diameter under normal nitrogen level, cross P1xP5 (43.9%) showed maximum positive and significant heterosis followed by P1xP3 (35.5%); whereas under stress nitrogen level, maximum positive and highly significant heterosis of better-parent were showed by crosses P1xP5 (46.8%) followed by P3xP5 (41.5%). The highest positive significant heterosis over mid-parents was recorded by P2xP4 (25.5%) followed by P2xP3 (24.7%) under normal nitrogen level, whereas under stress nitrogen level, cross P2xP4 gave maximum (31.4%) over mid-parents for 100-kernel weight followed by cross P2xP3 (22.1%). For better-parent of 100-kernel weight under both nitrogen levels, cross P2xP4 (23.1 and 29.3%) showed maximum positive and significant heterosis followed by P2xP3 (20.7 and 18.6%) under normal and stress nitrogen levels, respectively. For grain yield plant-1, the highest positive significant heterosis over mid-parents was recorded by P1xP3 (524.2 and 566.5%) followed by P1xP4 (406 and 452.1%) under normal and stress nitrogen levels, respectively. For better-parent of grain yield plant-1 under both nitrogen levels, cross P1xP3 (445.9 and 476.7%) showed maximum positive and significant heterosis followed by P3xP4 (323.5%) and P1xP4 (330.6%), respectively. The highest positive significant heterosis over mid-parents was recorded under stress nitrogen level for cross P1xP3(11.4%) over mid-parents for shelling percentage followed by cross P1xP4 (9.9%). For better- parent of shelling percentage under normal nitrogen level, cross P1xP4 (9.6%) showed maximum positive and significant heterosis; whereas under stress nitrogen level, cross P1xP3 (11.1%) showed maximum positive and significant heterosis followed by P1xP4 (9.4%). It could be concluded that cross P1xP3 recorded maximum positive and significant heterosis over mid and better-parents for shelling (%) under stress nitrogen level, (Table 5).
| Table 5: | Percentage of heterosis over mid-parent (M.P) and better parent (B.P) for F1 crosses of studied maize flowering and vegetative traits under normal and stress nitrogen levels |
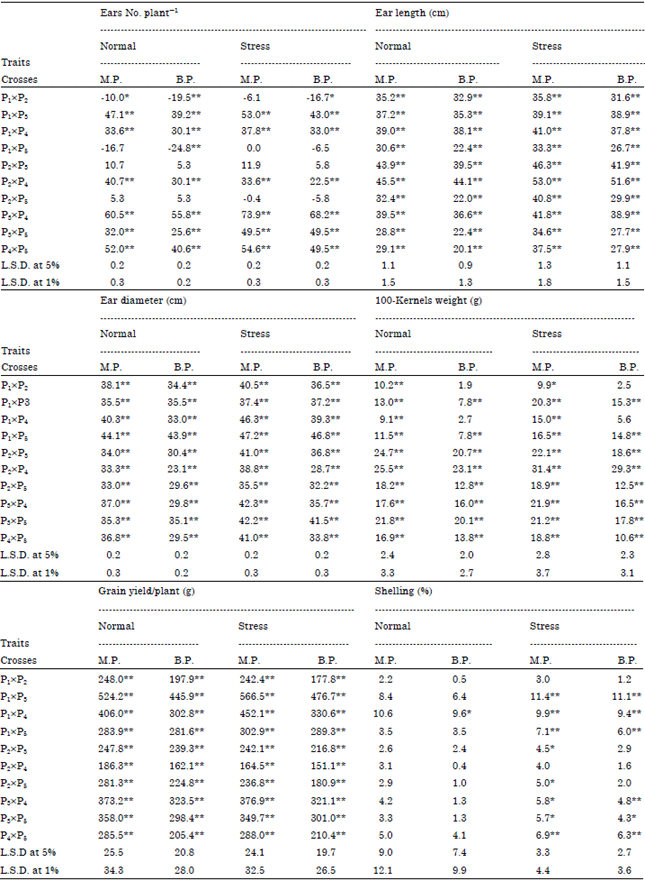 | |
| *, **Significant at 0.05 and 0.01 level of probability, respectively | |
REFERENCES
- Abdel-Moneam, M.A., A.N. Attia, M.I. El-Emery and E.A. Fayed, 2009. Combining ability and heterosis for some agronomic traits in crosses of maize. Pak. J. Biol. Sci., 12: 433-438.
CrossRefPubMedDirect Link - Akintoye, H.A., J.G. Kling and E.O. Lucas, 1999. N-use efficiency of single, double and synthetic maize lines grown at four N levels in three ecological zones of West Africa. Field Crops Res., 60: 189-199.
CrossRefDirect Link - Dadheech, A. and V.N. Joshi, 2007. Heterosis and combining ability for quality and yield in early maturing single cross hybrids of maize (Zea mays L.). Indian J. Agric. Res., 41: 210-214.
Direct Link - El-Badawy, M.E.M., 2013. Heterosis and combining ability in maize using diallel crosses among seven new inbred lines. Asian J. Crop Sci., 5: 1-13.
CrossRefDirect Link - Griffing, B., 1956. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci., 9: 463-493.
Direct Link - Irshad-Ul-Haq, M., S.U. Ajmal, M. Munir and M. Gulfaraz, 2010. Gene action studies of different quantitative traits in maize. Pak. J. Bot., 42: 1021-1030.
Direct Link - Soares, M.O., G.V. Miranda L.J.M., Guimaraes, I.E. Marriel and C.T. Guimaraes, 2011. Genetic parameters of a maize population in contrasting nitrogen levels. Rev. Cienc. Agron., 42: 168-174.
CrossRefDirect Link - Raun, W.R. and G.V. Johnson, 1999. Improving nitrogen use efficiency for cereal production. Agron. J., 91: 357-363.
CrossRefPubMedDirect Link - Sedhom, S.A., 1994. Estimation of general and specific combining ability In maize under two different planting dates. Ann. Agric. Sci. Moshtohor., 32: 119-130.
Direct Link