
M.A. Abdel-Moneam
Departmnet of Agron, Faculty of Agricultural, Mansoura University, Egypt
M.S. Sultan
Departmnet of Agron, Faculty of Agricultural, Mansoura University, Egypt
S.E. Sadek
Department of Maize Research, Institute of Field Crop, ARC, Egypt
M.S. Shalof
Department of Maize Research, Institute of Field Crop, ARC, Egypt
Asian Journal of Crop Science
Year: 2014 | Volume: 6 | Issue: 2 | Page No.: 101-111







ABSTRACT
A diallel cross among inbred lines of maize (Zea mays L.) with medium maturity and an evaluation to estimate heterosis and genetic parameters for ear length (cm), ear diameter (cm), No. of rows ear-1, No. of kernels row-1, weight of 100-kernels and grain yield (ardab feddan-1) were carried out. Hybrids mean squares were highly significant for the six traits under both planting dates and combined analysis, except ear diameter. Indicating that the hybrids performance are differed from planting date to another. Results indicating that the parental inbred P5 gave high grain yield. The highest grain yield was obtained from crosses (P4xP1) and (P5xP4) in combined, these crosses were significantly out yielded the two checks SC 155. SC 162 at 5%. Moreover, crosses (P1xP4), (P1xP5),(P6xP1), (P4xP2), (P2xP3) and (P3x P6) were insignificantly better than the checks. For ear length, P1xP4 (56.67 and 67.65%) showed maximum positive heterosis over better-parent and mid-parent, respectively. For ear diameter P2xP1 gave maximum positive heterosis over better-parent and over mid-parent, for No. of Kernels row-1 26 crosses had highly significant and positive heterosis over mid-parents, for No. of Kernels row-1 six crosses (P1x P2),(P2x P1), (P4 x P1), (P1 x P5), (P5xP1) and (P2xP6) had highly positively significant heterosis over better-parent. The highest positive significant heterosis over mid-parents for 100-kernel weight were recorded by 14 crosses. Cross P1xP2 showed maximum positive and significant heterosis over better-parent and mid-parents and only one cross P1xP4 gave positive and significant over check varieties for 100-kernel weight. For grain yield, cross P2xP4 (289.55%) and (295.73%) showed maximum positive and significant heterosis followed by P4xP2 (267.22 and 273.04%). Heritability in the narrow sense was the highest in No. of rows ear-1, while heritability in the broad sense was the highest in ear diameter. Heritability estimates in broad sense were medium (33-66%) of all studied traits except for ear diameter was high estimate (71%). Hence it could be concluded that these crosses may be useful for improving maize grain yield program.
PDF Abstract XML References Citation
Received: December 20, 2013;
Accepted: January 07, 2014;
Published: March 08, 2014
How to cite this article
M.A. Abdel-Moneam, M.S. Sultan, S.E. Sadek and M.S. Shalof, 2014. Estimation of Heterosis and Genetic Parameters for Yield and Yield Components in Maize Using the Diallel Cross Method. Asian Journal of Crop Science, 6: 101-111.
DOI: 10.3923/ajcs.2014.101.111
URL: https://scialert.net/abstract/?doi=ajcs.2014.101.111
DOI: 10.3923/ajcs.2014.101.111
URL: https://scialert.net/abstract/?doi=ajcs.2014.101.111
INTRODUCTION
Many procedures have been used by plant breeders to an attempt to increase of maize yields Geadelmann and Peterson, 1980). The use of heterosis started in 1933 when in the USA approximately 1% of the total farming average was planted with heterosis maize hybrids, while in 1953 the heterosis of the maize hybrids were expanded up to 96% (Sprague, 1962). The choice of the most efficient breeding program depends on the said information) (Liao, 1989; Pal and Prodham, 1994). The effects of general Combining Abilities (GCA) and Specific Combining Abilities (SCA) are important indicators of potential value for inbred lines in hybrid combinations. Differences in GCA effects have been attributed to additive, the interaction of additive x additive and the higher-order interactions of additive genetic effects in the base population, while differences in SCA effects have been attributed to non-additive genetic variance (Falconer, 1981). The concept of GCA and SCA has become increasingly important to plant breeders because of the widespread use of hybrid cultivars in many crops. The evaluation of crosses among inbred lines is an important step towards the development of hybrid varieties in maize) (Hallauer, 1990). This process ideally should be through the evaluation of all possible crosses (diallel crosses), where the merits of each inbred line can be determined. A Diallel analysis provides good information on the genetic identity of genotypes especially on dominance-recessive relations and some other genetic interactions. Diallel crosses have been used in genetic research to determinate the inheritance of a trait among a set of genotypes and to identify superior parents for hybrid or cultivar development (Yan and Kang, 2003). The main objective of our study was to estimate the heterosis and genetic parameters among these maize inbred lines and, consequently, to identify superior single-cross hybrids (SCH) developed from the studied new maize inbred lines.
MATERIALS AND METHODS
The following six new yellow parental inbred lines were studied: 10RF, 11RF, 39RF, 45RF, 48RF and 50RF. These lines were differed considerably in expression of various agronomy traits. Six inbred lines were crossed at Gemmeiza in a full diallel to give 30 crosses including reciprocal crosses in the summer of 2010 at Agricultural Research Center in Egypt (A.R.C.). The parents and their 30 F1 hybrids as well as two check hybrids (single cross 155 and single cross 162) were evaluated at Gemmeiza location on randomized complete block design (RCBD) with four replications in two different planting dates in 15 April and 15 May 2011. Kernels were hand-sown at 3 to 4 grains per hill then, thinned at two plants per hill after emergence. Each replication contained 38 plots and each plot consisted of one ridge with 6 m a long and spacing of 35 cm between plants within ridge and 80 cm between ridges. In Experiments, data were recorded on the following characters on plot basis: Ear length (cm), ear diameter (cm), No. of rows ear-1, No. of kernels row-1, weight of 100-kernels and grain yield which was adjusted to 15.5% moisture content (estimated in an ard fed-1).
Statistical analysis procedure: Analysis of variance for mean of performance according to the method outlined by Snedecor and Cochran (1977) was used for each experiment and then combined over the two planting dates. The L.S.D. test at 5 and 1% according to Steel and Torrie (1980) was used for comparison the mean performance of the different genotypes.
General Combining Ability (GCA) and Specific Combining Ability (SCA) effects were estimated according to Griffings (1956) Method 1 Model 1. In addition the mathematical models for a single inbred cross were tested for normality by statistical software. Then, data were analyzed using AGR 21 statically software the evaluating main genotype effects obtain GCA, SCA, reciprocal, maternal and non-maternal effects and their interaction with environment.
Broad sense heritability h2b and narrow sense heritability h2n for mean values over environments were calculated following the components of variance (Teklewold and Becker, 2005):
| r | = | No. of replications |
| g^i | = | General combining ability effects |
| s^ij | = | Specific combining ability effects |
| σ2e | = | Error mean squares |
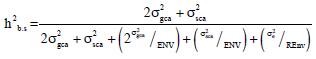
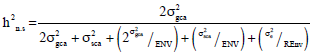
Baker (1978) suggested genetic ratio that the progeny performances could be predicted by the use of the ratio of combining ability variance components:
Genetic ratio = 2 MSgca/(2MSgca+MSsca)
Heterosis for all traits was estimated based on the behavior of the most outstanding parent, given that such estimation is useful to justify the use of hybrid seed (Fehr, 1991):
Heterosis (BP) (Heterobeltosis)(%) = [( F1-BP )/BP]x100
Heterosis (MP) (%) = [( F1-MP )/MP]x100
Heterosis (CV))(%) = [( F1-CV)/CV]x100
where, F1 = Performance of F1 hybrid; HP = Performance of the best parent, MP = Mide-parents and Cv = Check variety.
The difference of F1 means from the respective better parent value and check variety were evaluated as follows: LSD = t (2MSe/r)1/2.
The difference of F1 means from the respective mid- parents value was evaluated as follows: LSD =t (3MSe/2r)1/2.
where, MSe= the error mean squares; r = No. of replication and t = the table value of t at 5 or 1% level of significance. Combined analyses of variance based on RCBD, genetic parameters and comparison of quantitative traits means based on Duncan’s new multiple range test, were performed in Agrobase (2001) and Griffings (1956) method 1 (Model 1) diallel analysis.
RESULTS AND DISCUSSION
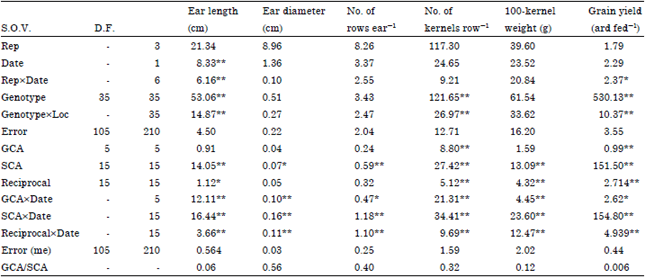
The analysis of variance for ordinary analysis and combining ability in two planting dates and combined data over two planting dates for ear length (cm), ear diameter (cm), No. of rows ear-1, No. of kernels row-1, Weight of 100-kernels and grain yield (ardab feddan-1) is presented in Table 1.
Results in Table 1 show that both General (GCA) and Specific (SCA) combining ability mean squares were significant or highly significant for all studied traits, except for ear length, ear diameter, No. of rows ear-1 and 100-kernel weight for (GCA).
| Table 1: | Analysis of variance for ordinary analysis and combining ability based combined data over two planting dates for studied traits |
 | |
| *,**Significant at 0.05 and 0.01 level of probability, respectively | |
These results indicated that both additive and non-additive types of gene effects were involved in the inheritance of these traits. The ratio of GCA/SCA was less than unity for all studied traits. Also results indicating that the non-additive genetic effects were more important and played the major role in the inheritance of all studied traits indicating the non-additive gene under both planting dates and combined analysis with a few previous exceptions.
On the other hand, reciprocals mean squares were significant or highly significant for all studied traits under both planting dates and combined analysis except ear diameter and No. of rows ear-1 under planting dates and combined data and ear length in the combined analysis. These results indicating that the maternal effect played an important role in the expression. These results are in agreement with those obtained by Sadek et al. (2001), Ji et al. (2006) and Kumar et al. (2006).
The interactions between GCA, SCA and reciprocals with planting dates (Table 1) were significant for all studied traits. The magnitude of the interaction was lowest for GCAxplanting dates than the SCAxplanting dates for all studied traits. This indicates that non-additive genetic variance was influenced by environment. The non-additive effect component interacted more with the environment than the additive. This conclusion supports the findings by Motawei (2005) and Singh and Roy (2007).
The closer of GCA/SCA genetic ratio (Baker, 1978) to unity shows the predictability based on GCA alone. Also the GCA/SCA ratio reveals that different traits show an additive or non-additive genetic effect. A GCA/SCA ratio with a value greater than one indicates additive genetic effect, whereas a GCA/SCA ratio with a value lower than one indicates dominant genetic effect. Mean squares of GCA/SCA ratio were less than unit for all studied traits.
Mean performance: The combined data of mean performance across the two planting dates for grain yield and other agronomic traits of the six parental inbred lines , 30 F1 crosses and two check hybrids were presented in Table 2. Results indicate that the parental inbred P5 was highly grain yield parent. Considering of ear length for genotypes are presented in Table 2. Ear length for parents ranged from13.8 to 17.5 cm over the two dates. The highest parental inbred line was P6 over the two dates.
| Table 2: | Mean performance of maize genotypes at their combined for the traits studied during 2011 season |
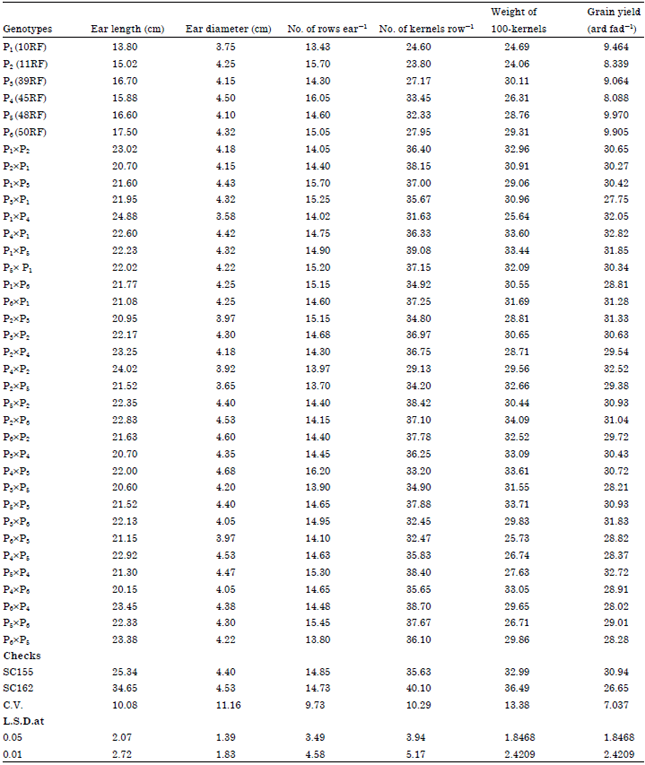 | |
The differences between ear length for crosses were significant and ranged from 20.15 to 24.88 in over two dates. ear diameter for parental inbred line ranged from 3.75 to 4.5 cm in combined data over two dates. The highest value was recorded by P4 in combined data. Meanwhile, the lowest value was recorded by P1 in combined data. The differences between ear diameter for all crosses studied were non-significant in both planting dates and combined over them. Ear diameter ranged from 3.58 to 4.68 cm over the two dates in combined data, Singh (2005), Machado et al. (2009) and Sultan et al. (2011) came to similar results. Rows number for genotypes ranged from 13.43 to 16.05 over the two dates. The highest value was recorded by P4 in combined data. Meanwhile, the lowest value was recorded by P1 in two planting dates and combined data. The highest value was recorded by (P4xP3) in combined data, Meanwhile, the lowest value was recorded by cross (P2xP5) in combined data. Number of Kernels row-1 for parents ranged from 23.80 to 33.45 in over the two dates in combined data. The highest value was recorded by P4 in combined data. Meanwhile, the lowest value was recorded by P2 in combined data. Number of Kernels row-1 for all studied crosses were significant compared with S.C.162. 100-kernel weight for parents ranged from 24.06 to 30.11g in combined data. The difference among crosses for 100-kernel weight were significant. The highest grain yield was obtained from crosses (P4xP1) 32.82 ard fed-1 and (P5xP4) 32.72 ard fed-1 in combined , these crosses were significantly out yielded the two checks SC 155 and SC 162 at 5%. Moreover, crosses (P1xP4) 32.05, (P1xP5) 31.85, (P6xP1) 31.28, (P4xP2) 32.52, (P2xP3) 31.33 and (P3x P6) 31.83 ard fed-1 were insignificantly better than the checks. Hence, it could be concluded that these crosses may be useful for improving maize grain yield program.
HETEROSIS STUDIES
Heterosis is a major reason for the commercial maize industry as well as for the success of breeding efforts in many other crops. Beginning in the early 1900s, scientists began designing experiments to determine the mechanism of heterosis. Over the years, the majority of the scientific community has attributed heterosis to dominance or over dominance and recently scientists have reported that epistasis and linkage are major contributors. One common theme throughout the last century has been that no one hypothesis of heterosis holds true for every experiment or every organism (Budak et al., 2002).
Results given in Table 3 indicated that all the crosses manifested highly significant and positive heterosis over mid-parent and better-parent for ear length, P1xP4 (56.67 and 67.65%) showed maximum positive heterosis over better-parent and mid-parent, respectively. All crosses show highly significant and negative heterosis over check varieties for ear length, P4xP6 (-41.84%) showed maximum negative heterosis over check varieties, Katta et al. (2007); Alam et al. (2008) and Patel et al. (2009) came to the similar conclusion. Data in Table 3 clear that the highest significant and positive heterosis over mid-parents for ear diameter was recorded by P1xP3 (15.06%) followed by P3xP1 (12.20%). These results are in conformity by the finding of Katta et al. (2007) and Alam et al. (2008).
For number of Kernels row-1 in Table 3 regarding that the significant and positive heterosis over mid-parent and better-parent value were recorded but, showed significant or highly significant and positive heterosis by 21 crosses over better- parent, for number of Kernels row-1. 26 crosses had highly significant and positive heterosis over mid-parent. These results are in agreement with those obtained by Katta et al. (2007) and Alam et al. (2008).
In Table 3 regarding to 100-kernel weight, the highest significant heterosis and positive over better-parent was recorded for 6 crosses (P1xP2, P2xP1, P4xP1, P1xP5, P5xP1 and P2xP6) had highly positively significant. The highest positive significant heterosis for 100-kernel weight were recorded by 14 crosses, had highly significant and positively effect. Cross P1xP2 (33.49%) and (35.24%) showed maximum positive and significant heterosis over better-parent and mid-parents, respectively and only one cross P1xP4 (29.73%) gave positive and significant heterosis over check varieties for 100-kernel weight, similar results were obtained by Katta et al. (2007); Alam et al. (2008) and Abdel-Moneam et al. (2009). In Table 3, data for grain yield (ard fed-1) revealed that the highest significant and positive heterosis effect over better-parent and mid-parents was recorded in all crosses. Cross P2xP4 (289.55%) and (295.73%) showed maximum positive and significant heterosis followed by P4xP2 (267.22 and 273.04%) over better-parent and mid-parents, respectively. These results are in accordance with those of El-Ghonemy and Ibrahim (2010); Sultan et al. (2010) and Amanullah et al. (2011).
| Table 3: | Estimates of heterotic effects of 30 yellow single crosses maize genotypes at Gemmeiza in their combined for the studied traits in growing season 2011 |
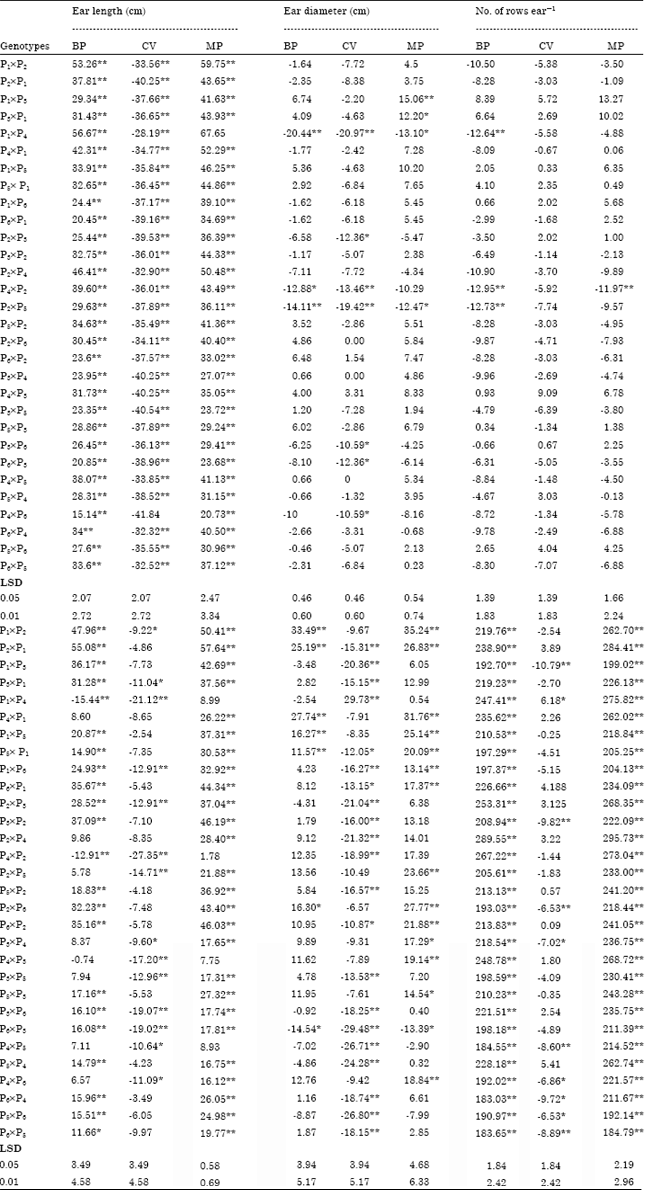 | |
| *, **Significant at 0.05 and 0.01 level of probability, respectively | |
Heritability and genetic parameters: Estimates of variance for general (δ2gca) and specific (δ2sca) combining ability and their interaction with planting dates showed that in Table 4 (δ2gca) was higher than (δ2 sca) for all studied traits except ear length, this indicated that the additive gene action was more important in the inheritance of all studied traits. On the other side the δ2 gcaEnv was higher than δ2 scaEnv for all studied traits (Osman and Ibrahim, 2007). For δ2 gca/δ2 sca and genetic ratio estimation of genetic parameters is given in Table 4 revealed that the closer genetic ratio (Baker, 1978) to unity shows the predictability based on GCA alone. Also the GCA/SCA ratio reveals that different traits show an additive or non-additive genetic effect.
| Table 4: | Estimation of genetic parameters of maize in a 6x6 diallel crosses for the studied traits in growing season 2011 |
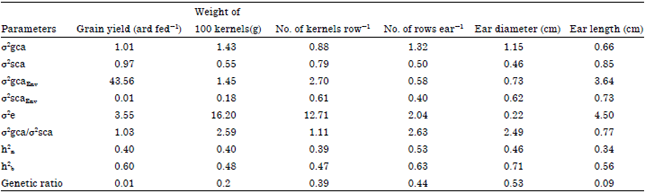 | |
A δ2gca/δ2sca ratio with a value greater than one indicates additive genetic effect, whereas a genetic ratio with a value lower than one indicates dominant genetic effect. The predominance of SCA variance denotes that non-additive genetic effects were largely influencing the expression of these traits; hence, heterosis and use of hybrid vigor could be applied for improving them. These results were in agreement with reports of other researchers about predominance of non-additive genetic effects (Alam et al., 2008) However, in contrast to our results, other researchers indicated predominance of additive genetic effects (Srdic et al., 2007).
Heritability is considered to be one of the important parameters to express relative genetic variability whether in a broad or narrow sense. Estimates of heritability in broad and narrow sense for all traits study full-sib families at combined analysis are presented in Table 4.
Data in Table 4 for studied traits showed that heritability estimates in broad sense were generally higher at combined data. In the combined data percentage of heritability in the narrow sense for studied traits ranged from 34% for ear lenght to 53% for No. of rows ear-1, in broad sense ranged from 47% for No. of Kernels row-1 to 71% for ear diameter. On the other hand heritability in the narrow sense was the highest in No. of rows ear-1, while heritability in the broad sense was the highest in ear diameter. Heritability estimates in broad sense were medium (47-63%) for all studied traits except for ear diameter was high estimate (71%), Mostafavi et al. (2009) and Wannows et al. (2010) came to the similar conclusion.
REFERENCES
- Abdel-Moneam, M.A., A.N. Attia, M.I. El-Emery and E.A. Fayed, 2009. Combining ability and heterosis for some agronomic traits in crosses of maize. Pak. J. Biol. Sci., 12: 433-438.
CrossRefPubMedDirect Link - Alam, A.K.M.M., S. Ahmed, M. Begum and M.K. Sultan, 2008. Heterosis and combining ability for grain yield and its contributing characters in maize. Bangladesh J. Agric. Res., 33: 375-379.
Direct Link - Amanullah, S.J., M. Mansoor and M.A. Khan, 2011. Heterosis studies in diallel crosses of maize. Sarhad J. Agric., 27: 207-211.
Direct Link - Griffing, B., 1956. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci., 9: 463-493.
Direct Link - Budak, H., L. Cesurer, Y. Bolekt, T. Dokuyucu and A. Akkaya, 2002. Understanding of heterosis. KSU J. Sci. Eng., 5: 68-75.
Direct Link - Machado, J.C., J.C. Souza, M.A.P. de Ramalho and J.L. Lima, 2009. Stability of combining ability effects in maize hybrids. Scientia Agricola, 66: 494-498.
Direct Link - Kumar, R., M. Singh and M.S. Narwal, 2006. Combining ability analysis for grain yield and its contributing traits in maize (Zea mays L.). Natl. J. Plant Improv., 8: 62-66.
Direct Link - Singh, P.K. and A.K. Roy, 2007. Diallel analysis of inbred lines in maize (Zea mays L.). Int. J. Agric. Sci., 3: 213-216.
Direct Link - Srdic, J., Z. Pajic and S. Drinid-Mladenovic, 2007. Inheritance of maize grain yield components. Maydica, 52: 261-264.
Direct Link - Teklewold, A. and H.C. Becker, 2005. Heterosis and combining ability in a diallel cross of Ethiopian mustard inbred lines. Crop Sci., 45: 2629-2635.
Direct Link - Wannows, A.A., H.K. Azzam, S.A. Al-Ahmad, 2010. Genetic variances, heritability, correlation and path coefficient analysis in yellow maize crosses (Zea mays L.). Agric. Biol. J. N. Am., 1: 630-637.
Direct Link