
P. Theerakulpisut
Department of Biology, Faculty of Science, Khon Kaen University, Khon Kaen 40002, Thailand
S. Phongngarm
Department of Biology, Faculty of Science, Khon Kaen University, Khon Kaen 40002, Thailand
Asian Journal of Crop Science
Year: 2013 | Volume: 5 | Issue: 4 | Page No.: 405-415







ABSTRACT
Synthesis and accumulation of small organic osmolytes is a mechanism which protects plants from damaging effects of abiotic stresses including salinity. The effects of three different osmoprotectants; trehalose (Tre), proline (Pro), and sorbitol (Sor) on growth and physiology of rice seedlings subjected to salt stress were compared in this study. Four-day-old seedlings of rice cvs. Pokkali (PK; salt-tolerant) and Patumthani 60 (PT60; salt-sensitive) were treated with 5 mM and 10 mM Tre, Pro or Sor without and with 170 mM NaCl for 10 days. NaCl inhibited growth, caused a reduction in chlorophyll and K+ and increase Na+ and Na+: K+ ratio and the adverse effects of salt were more pronounced in PT60. Under salt stress, exogenous Tre was capable of alleviating the deleterious effects of NaCl by enhancing growth, maintaining chlorophyll content and reducing Na+: K+ ratio. In contrast, Pro exacerbated the effects of NaCl by further reducing plant height, fresh and dry weights, although, it did not have any effects on chlorophyll content and ion accumulation. Exogenous Sor neither alleviated nor exacerbated the effects of NaCl. These results indicated that Tre was the most effective osmoprotectant which offered protective roles for rice seedlings subjected to salt stress and presented a practical implication for rice cultivation in salt-affected soils.
PDF Abstract XML References Citation
Received: April 04, 2013;
Accepted: April 30, 2013;
Published: September 09, 2013
How to cite this article
P. Theerakulpisut and S. Phongngarm, 2013. Alleviation of Adverse Effects of Salt Stress on Rice Seedlings by Exogenous Trehalose. Asian Journal of Crop Science, 5: 405-415.
DOI: 10.3923/ajcs.2013.405.415
URL: https://scialert.net/abstract/?doi=ajcs.2013.405.415
DOI: 10.3923/ajcs.2013.405.415
URL: https://scialert.net/abstract/?doi=ajcs.2013.405.415
INTRODUCTION
Saline soil is one of the dominant abiotic stresses limiting crop production particularly in the arid and semi-arid regions of the world. High concentrations of salt resulting from natural processes or disarrangement in irrigated agriculture result in inhibition of plant growth and yield (Tester and Bacic, 2005). Salinity primarily imposes on plants an osmotic stress and secondarily ion toxicity stress. Salinity stress also causes nutrient imbalances, due to the competition of Na+ and Cl¯ with nutrients such as K+, Ca2+ and NO3¯ which reduces plant growth (Hu and Schmidhalter, 2005). Plants synthesize and accumulate many compatible solutes or osmoprotectants in the cytoplasm to increase their hyperosmotic tolerance against salt-induced water loss from the cells. Osmoprotectants are reported as membrane stabilizer which can protect enzymes, organelles and cell membrane that are sensitive to dehydration and ionic damage under osmotic and ionic stress (Hare et al., 1998; Sairam and Tyagi, 2004). This process is also required to balance the osmotic potential of Na+ and Cl¯ being accumulated in the vacuole (Wyn et al., 1997).
There are several types of osmoprotectants for example; betaines (fully N-methylated amino acid derivatives) and related compounds such as dimethylsulfoniopropionate (DMSP) and choline-O-sulfate; certain amino acids like proline (Pro) and ectoine; nonreducing sugars such as trehalose (Tre; 1,1 α-D-glucopyranosal-α-D-glucopyranoside) and sugar alcohol such as mannitol and sorbitol (Turkan and Demiral, 2009).
Tre is a nonreducing disaccharide consisting of two subunits of glucose bound by α, α 1,1-, linkage. Tre plays an important physiological role as a storage carbohydrate and as a protector from abiotic stress (Llorente et al., 2007). Genetic modification of tobacco transformed with ots A (from E. coli) demonstrated that photosynthetic capacity per unit leaf area and per leaf dry weight were increased. However, a large number of photosynthetic capacities did not translate into greater relative growth rate or biomass (Pellny et al., 2004). Also, exogenous application of Tre has been reported to give a good advantage to plants. Unno and Maeda (2008) reported that the survival rate of perennial ryegrass protoplasts increased under NaCl stress when Tre was introduced into the protoplasts.
Pro is also considered to be involved in the protection of enzymes (Solomon et al., 1994) and cellular structures (Van Rensburg et al., 1993) and to act as a free radical scavenger. Finally, its biosynthesis could be associated with the regulation of cytosolic pH (Venekamp, 1989) or with the production of NADP+ for the stimulation of the pentose phosphate pathway (Hagedorn and Phang, 1986). Several works reported that exogenous application of Pro can show a beneficial effect to protect plants from abiotic stress for example; in mung bean (Posmyk and Janas, 2007), seeds primed with proline (5 mM) showed the highest growth during chilling period compared to those primed with water and the control (non-primed). Moreover, under oxidative stress, adding exogenous Pro to grapevine (Ozden et al., 2009) had a direct positive effect on antioxidant enzyme system and membrane phase change and a reduction in Malondialdehyde (MDA) and Electrolyte Leakage (EL).
Sor is a sugar alcohol from glucose derivative, mostly found as seed component in many crop plants and may contribute to the desiccation tolerance (Sairam and Tyagi, 2004). Exogenous Sor improved the quantity and quality of wheat callus production. Furthermore, application of Sor stimulated the green spot formation (Hassan et al., 2009). Feng et al. (2011) illustrated that not only Sor promoted plantlet regeneration frequency of rice calli but also acted as a chemical signal to directly or indirectly alter gene expression.
This study evaluated the effect of three osmoprotectants (Tre, Pro and Sor) on growth and physiology of two rice cultivars: Pokkali (PK; salt-tolerant) and Patumthani 60 (PT60; salt-sensitive) subjected to NaCl stress at 170 mM.
MATERIALS AND METHODS
Plant materials and growth condition: Seeds of rice cvs. PK and PT60 (kindly provided by Pathumthani Rice Research Institute, Thailand) were sterilized with 70% ethyl alcohol for 2 min and then incubated with 35% sodium hypochlorite for 1 h and washed extensively with three changes of distilled water. Sterilized seeds were germinated in half strength Murashige and Skoog (½ MS) medium (Murashige and Skoog, 1962) for 4 days. After 4 days, seedlings of each cultivar were transferred to fresh ½ MS medium (MS1), ½ MS medium added with 5 or 10 mM Tre (MS2 and MS3), or Pro (MS4 and MS5), or Sor (MS6 and MS7). For salt stress treatments, the 4-day-old seedlings were transferred to ½ MS medium added with 170 mM NaCl (MS8), ½ MS medium plus 170 mM NaCl supplemented with 5 or 10 mM Tre (MS9 and MS10), or Pro (MS11 and MS12), or Sor (MS13 and MS14). The plants were incubated at 25±2°C under the light 16 h days-1 (40 μmol m-2 sec-1) for 10 days. Plants were harvested and analyzed for shoot length, fresh and dry weights, chlorophyll and Na+ and K+ ion concentration.
Growth and ion measurements: Harvested leaf segments were washed three times (one min each) with distilled water, dried at 80°C for 2 days, extracted with H2NO3, H2ClO4 and HCl at 200-300°C and the Na+ and K+ content were measured with atomic absorption flame photometer (Model GBC 932 AA).
Chlorophyll determination: Chlorophyll was extracted from fresh tissue in 80% (v/v) acetone by grinding samples for 10 min, the homogenate was then filtered through filter paper. The absorbance of the filtrate was measured at 645 and 663 nm. The chlorophyll content was calculated using the following equation (Arnon, 1949):
Total chlorophyll (mg/g tissue) = [20.2 (A645)+8.02 (A663)]xV/(1000xW)
Where:
| V | = | Volume of supernatant (mL) |
| W | = | Weight of tissue (g) |
Statistical analysis: All results were presented as means±SE. The significance of differences between the mean values was determined by two-way ANOVA. The p values smaller or equal to 0.05 was considered statistically different.
RESULTS
Effect of osmoprotectants on growth of rice seedlings: Under the normal growth condition (MS1-MS7), addition of Tre tended to promote seedling growth of both cultivars whereas Pro and Sor did not. Growth of seedlings of both PK and PT60 stressed with 170 mM NaCl (MS8) was seriously inhibited. Exogenous supply of Tre (MS9 and MS10) dramatically reversed the negative effects of salt and significantly improved the overall growth of the seedlings. Sor (MS13 and MS14) exhibited a marginally positive effect whereas Pro (MS11 and MS12) did not help reverse growth inhibition of salt-stressed seedlings.
The effects of NaCl and osmoprotectants on the height of rice seedlings were shown in Fig. 1. In the absence of NaCl (MS1-MS7), the tallest plants were observed in PK and PT60 plants supplied with 5 and 10 mM Tre (MS2 and MS3). The increases in height were 9.44 and 21.75% for PK, 10.73 and 20.39% for PT60, as compared to control plants. Addition of 5 mM Pro (MS4) showed no significant changes in the height of PK plants but higher concentration of Pro (10 mM; MS5) caused a marked decrease (13.46%). Conversely, for PT60 both concentrations of Pro (MS4 and MS5) did not alter plant height. Sor (5 and 10 mM; MS6 and MS7) did not have any effects on the height of seedlings of both cultivars. With NaCl treatments (MS8-MS14), exogenous Tre (10 mM; MS10) significantly increased the height of seedlings (40.03% increase for PK and 40.97% increase for PT60). Lower concentration of Tre (5 mM; MS9) did not show a beneficial effect on PK but slightly increased seedling height of PT60 (12.61%). Application of Pro (5 and 10 mM; MS11 and MS12) significantly suppressed the height of PK plants (12.50% and 19.35% decrease, respectively) while no significant alteration was detected for PT60. In ½ MS media combined with NaCl and Sor, 5 mM Sor (MS13) no detectable change on PK and PT60 was observed but with the addition of 10 mM Sor (MS14) the height of PT60 was slightly enhanced (15.90%).
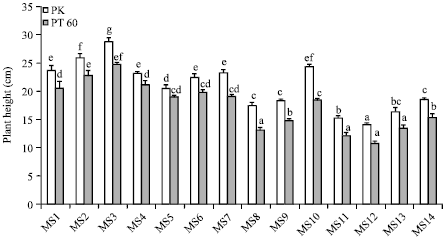 | |
| Fig. 1: | The effect of osmoprotectants on shoot length of rice cvs. PK and PT60 without (MS1-MS7) or with 170 mM NaCl (MS8-MS14). MS1-control; MS2, MS3-5 and 10 mM Tre; MS4, MS5-5 and 10 mM Pro; MS6, MS7-5 and 10 mM Sor; MS8-170 mM NaCl; MS9, MS10-NaCl+5 or 10 mM Tre; MS11, MS12-NaCl+5 or 10 mM Pro; MS13, MS14-NaCl+5 or 10 mM Sor. The values showed means±SE. Different small letters indicated that the means are significantly different (p≤0.05) |
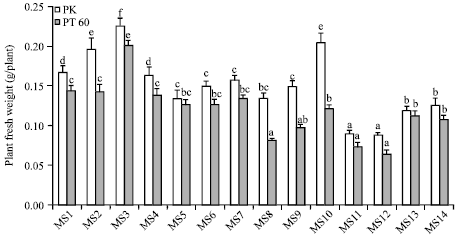 | |
| Fig. 2: | The effect of osmoprotectants on plant fresh weight of rice cvs. PK and PT60 without (MS1-MS7) or with 170 mM NaCl (MS8-MS14). See Fig. 1 for explanation of the treatments. The values showed means±SE. Different small letters indicated that the means are significantly different (p≤0.05) |
As shown in Fig. 2, under the normal growth condition, PK and PT60 plants supplemented with 10 mM Tre had 34.63 and 40.21% increased fresh weight when compared with the control plants. Supplementation with Tre (5 mM) caused a slightly increased PK fresh weight (17.61%) but had no significant effects on fresh weight of PT60. The application of 10 mM Pro significantly decreased PK and PT60 fresh weights (19.88 and 11.98%, respectively). There was a slight decrease in fresh weight of both cultivars when treated with Sor. Under salt stress, exogenous Tre promoted growth of both PK and PT60. Fresh weights of plants supplied with 10 mM Tre increased dramatically, 52.34% for PK and 50.31% for PT60, as compared to salt-stressed plants without the osmoprotectant. Tre at 5 mM also slightly increased plant fresh weight (11.45% for PK and 20.05% for PT60). In contrast, Pro induced a marked decrease in PK fresh weight i.e. 33.16 and 34.65% in 5 and 10 mM Pro, respectively. However, Pro did not affect PT60 fresh weight. Sor, on the other hand, did not have any effects on PK fresh weight but significantly enhanced PT60 fresh weight i.e., 38.37 and 32.47% in 5 and 10 mM Sor, respectively.
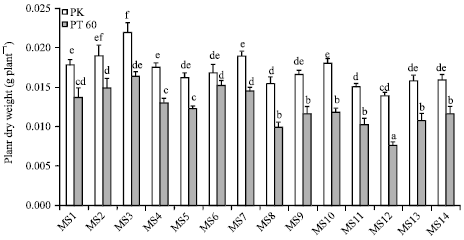 | |
| Fig. 3: | The effect of osmoprotectants on plant dry weight of rice cvs. PK and PT60 without (MS1-MS7) or with 170 mM NaCl (MS8-MS14). See Fig. 1 for explanation of the treatments. The values showed means±SE. Different small letters indicated that the means are significantly different (p≤0.05) |
In the absence of NaCl, 10 mM Tre significantly enhanced seedling growth resulting in 12.36 and 19.71% increase in dry weight for PK and PT60, respectively (Fig. 3). In contrast, either Pro or Sor did not show any beneficial effects. Under salt treatments, 10 mM Tre also led to a significant increase in PK dry weight (16.88%) as compared to salt-stressed PK plants. However, both concentrations of Tre tested in salt-stressed PT60, did not affect plant dry weight. Exogenous Pro at 5 mM did not improve dry weight of both cultivars. Moreover, 10 mM Pro markedly suppressed PT60 plants dry weight (23.23%) when compared to PT60 treated with salt alone. For plants supplied with Sor at 5 and 10 mM, no significant changes in dry weight were observed.
Effect of osmoprotectants on chlorophyll content of rice seedlings: The effects of osmoprotectants on chlorophyll content of rice seedlings are shown in Fig. 4. In the normal nutrient media, increasing concentration of Tre from 5 to 10 mM progressively increased chlorophyll content in rice seedlings of both cultivars (4.18 and 19.89% for PK, 36.85 and 48.67% for PT60). On the other hand, exogenous Pro (5 and 10 mM) caused a significant reduction in chlorophyll content of PK (11.91 and 19.34% reduction) but did not have any effects on that of PT60. In case of Sor, no significant difference was observed in PK while chlorophyll content of PT60 was significantly improved (25.85 and 32.84% in 5 and 10 mM Sor, respectively).
NaCl in the saline nutrient media significantly reduced chlorophyll content of rice seedlings. Compared to salt-stressed seedlings, the seedlings grown in saline nutrients containing 5 and 10 mM Tre enhanced the chlorophyll content (26.01 and 26.93% for PK; 29.03 and 40.20% for PT60). Exogenous Pro (5 and 10 mM) caused slightly increased chlorophyll content in PK (8.38 and 6.23%) but it did not improve chlorophyll content in PT60. Salt-treated rice seedlings supplemented with Sor at 5 and 10 mM showed a slight increase in chlorophyll content (9.08 and 18.23% for PK; 23.95 and 20.45% for PT60).
Effect of osmoprotectants on Na+ and K+ concentration of rice seedlings: The concentration of sodium ions (Na+) in rice shoots grown under various media were determined (Fig. 5a). Under normal growth condition, plants in all treatment groups had very low concentration of Na+.
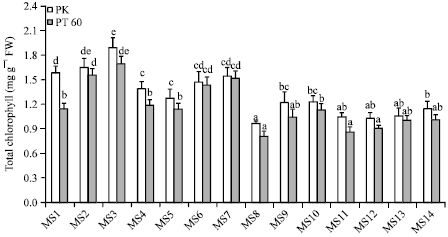 | |
| Fig. 4: | The effect of osmoprotectants on total chlorophyll of rice cvs. PK and PT60 without (MS1-MS7) or with 170 mM NaCl (MS8-MS14). See Fig. 1 for explanation of the treatments. The values showed means±SE. Different small letters indicated that the means are significantly different (p = 0.05) |
When the plants were salt-stressed, Na+ increased several folds and the sensitive PT60 accumulated markedly higher Na+ than the tolerant PK. For PK, none of the osmoprotectants had any effects on the Na+ content. However, PT60 plants grown in nutrient media containing NaCl plus Tre at 5 or 10 mM showed a significant reduction in Na+ content (12.4 and 30.4%, respectively) as compared to salt-stressed PT60 without the osmoprotectant.
The concentrations of potassium ion (K+) in shoots of PK and PT60 are shown in Fig. 5b. In the absence of NaCl, application of Tre (5 and 10 mM) caused a substantial increase in K+ of both PK and PT60 (118.09 and 43.33% for PK; 43.00 and 42.95% for PT60) as compared to controls. Exogenous Pro also enhanced K+ accumulation of both rice cultivars. PK plants were able to accumulate higher K+ when treated with 5 mM than 10 mM Pro whereas PT60 plants accumulated K+ better in 10 mM than in 5 mM Pro. Exogenous Sor had a positive effect on K+ accumulation of PK but not PT60. In saline nutrient media, accumulation of K+ was reduced in both cultivars in all treatments. Exogenous Tre, Pro and Sor had no effects on K+ accumulation in both rice cultivars.
NaCl stress caused a dramatic rise in Na+: K+ ratio in both cultivars (Fig. 5c). There was a marked decrease in Na+:K+ ratio in PK plants grown in MS medium containing NaCl and 5 or 10 mM Tre (20.79 and 11.88% decrease as compared to PK plants stressed with NaCl alone). Supplement of 10 mM Tre to salt-stressed PT60 also reduced Na+:K+ ratio (24.50%) compared to PT60 exposed with NaCl alone. Exogenous Pro in both concentrations did not alter Na+:K+ ratios in both PK and PT60. Exogenous Sor significantly reduced Na+: K+ ratios only in PT60 (15.23 and 12.58% reduction for 5 and 10 mM Sor, respectively).
DISCUSSION
Three different osmoprotectants; Tre, Pro and Sor influenced growth and physiology of both tolerant and sensitive rice cultivars in normal and salt-stress conditions. Without salt stress, Tre was the most effective in enhancing the seedling growth. In addition, under salt stress, Tre also gave the most beneficial effects on PK (plant height, fresh and dry weights) and PT60 (plant height and fresh weight). These observations are in good agreement with the results of exogenous Tre applied in maize. Supplementation of Tre in maize before exposure to salt treatment was able to improve growth of salt-stressed and unstressed plants (Zeid, 2009).
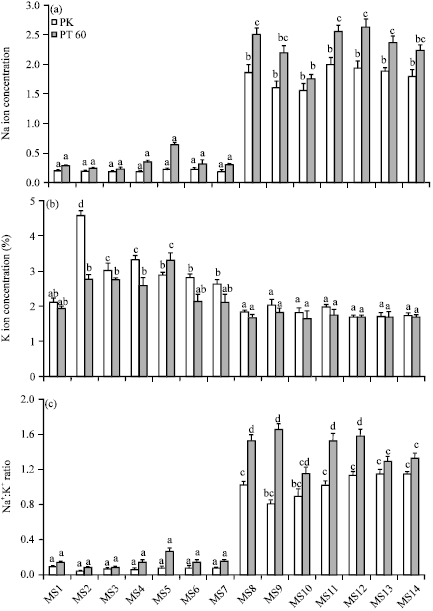 | |
| Fig. 5(a-c): | The effect of osmoprotectants on (a) Na+ concentration (b) K+ concentration and (c) Na+:K+ ratio of rice cvs. PK and PT60 without (MS1-MS7) or with 170 mM NaCl (MS8-MS14). See Fig. 1 for explanation of the treatments. The values showed means±SE. Different small letters indicated that the means are significantly different (p = 0.05) |
Among the three osmoprotectants tested, Pro was the least effective in alleviating NaCl-induced growth inhibition. Moreover, at higher concentration (10 mM) Pro suppressed growth of the salt-sensitive PT60. Similar results have been found in a desert plant (Pancratium maritimum L.) that Pro did not improve dry weight of salt-stressed seedlings (Khedr et al., 2003). Hellman et al. (2000) found that exogenous Pro caused the plants to accumulate high level of P5C, an intermediate in Pro synthesis pathway, which was toxic and growth inhibitory. Although, application of Sor did not promote plant growth in PK, it showed an ability to enhance growth of PT60. In Japanese pear it was also found that exogenous Sor was beneficial for in vitro growth by increasing shoot proliferation efficiency and fresh mass (Kadota et al., 2001).
Chlorophyll content is widely used as an index to indicate the abiotic tolerance level in plants. Protection of chloroplast and photosynthetic machinery including the chlorophyll content is the first target of defense under stressful conditions. Exogenous Tre increased chlorophyll content in PK and PT60 plants with or without NaCl treatments. Application of Tre in salt-stressed and unstressed leaves of maize increased chlorophyll content and enhanced the Hill-reaction activity (Zeid, 2009). Ali and Ashraf (2011) reported that foliar application of Tre to maize caused significantly increased biomass and improved some key photosynthetic attributes. Under drought stress, Tre improved drought tolerance of maize plants by up-regulating photosynthetic and water relation attributes as well as antioxidant defense mechanism. The data obtained in this work showed that Pro had a small positive effect on improving chlorophyll content in PK under salt stress. Ben Ahmed et al. (2010) also found that Pro supplement ameliorated the reduction of photosynthetic pigments under salt stress in young olive tree. In Arabidopsis under drought stress, external Pro was involved in the protection of chloroplast structures by scavenging ROS and maintained PSII functions (Moustakas et al., 2012). Low concentrations of Sor (5 and 10 mM) slightly improved chlorophyll content of rice seedlings in the absence or presence of NaCl. In contrast, treatments of maize seedlings with high concentration of Sor (>200 mM) induced water deficit and reduced chlorophyll content (Jain et al., 2010).
Supplementing Tre in the media caused a slight decrease in Na+:K+ ratio under salt stress. These results were similar to that found in maize that Tre increased the ratio of K+/Na+ ions under salt stress condition (Zeid, 2009). Chen et al. (2007) reported the mitigating effect of exogenous Pro (1 and 10 mM) on NaCl-induced K+ efflux which were more pronounced in salt-sensitive than salt-tolerant barley cultivar. However, our results showed that Pro did not alleviate the negative effects of salt on Na+:K+ ratio in both rice cultivars. Recently, Umar et al. (2011) found that Na+:K+ ratio in salt-stressed Brassica campestris L. could be directly improved by the addition of K fertilizer.
While exogenous Tre obviously showed some mitigating effects, exogenous Sor did not have any beneficial or deleterious effects on salt-stressed plants. Nevertheless, Pro was found to exacerbate the effect of NaCl by further reducing plant height, fresh weight and dry weight, although, it did not have any effects on chlorophyll content and ion accumulation. Similar results were observed by Garcia et al. (1997) in two rice cultivars PK and IR 29 (salt-sensitive). Pro either had no effects or in some cases exacerbated the effects of NaCl on growth inhibition, chlorophyll loss and induction of salT gene, the highly sensitive marker for plant stress. Tre, on the other hand, reduces Na+ accumulation, salT expression and growth inhibition. Many studies have demonstrated that transgenic plants overexpressing either TPS or TPP gene, or both simultaneously, from E. coil (OstA and OstB) (Garg et al., 2002; Jang et al., 2003), Saccharomyces cerevisiae (ScTPS1 and ScTPS2) (Cortina and Macia-Culianez, 2005; Karim et al., 2007) and rice (OsTPS1 (Li et al., 2012), OsTPP1 (Ge et al., 2008)) enhanced plants tolerance to abiotic stresses.
CONCLUSION
It can be concluded from this study that exogenous Tre (10 mM) was beneficial to both rice cultivars by improving Na+:K+ status and mitigating loss of chlorophyll which led to the observed enhancement in plant growth. Exogenous application of Tre is the more appropriate target than Pro and Sor for inducing certain physiological processes leading to improvement of salt tolerance in rice seedlings.
ACKNOWLEDGMENTS
This research was supported by Khon Kaen University Research Grant awarded to the Salt tolerant Rice Research Group. The authors are grateful for the kind gift of rice seeds from Pathumthani Rice Research Institute, Thailand.
REFERENCES
- Ali, Q. and M. Ashraf, 2011. Induction of drought tolerance in maize (Zea mays L.) due to exogenous application of trehalose: Growth, photosynthesis, water relations and oxidative defence mechanism. J. Agron. Crop Sci., 197: 258-271.
CrossRefDirect Link - Ben Ahmed, C., B. Ben Rouina, S. Sensoy, M. Boukhriss and F. Ben Abdullah, 2010. Exogenous proline effects on photosynthetic performance and antioxidant defense system of young olive tree. J. Agric. Food Chem., 58: 4216-4222.
CrossRefDirect Link - Arnon, D.I., 1949. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol., 24: 1-15.
CrossRefPubMedDirect Link - Chen, Z., T.A. Cuin, M. Zhou, A. Twomey, P.N. Bodapati and S.N. Shabala, 2007. Compatible solute accumulation and stress-mitigating effects in barley genotypes contrasting in their salt tolerance. J. Exp. Bot., 58: 4245-4255.
CrossRefDirect Link - Cortina, C. and F.A. Macia-Culianez, 2005. Tomato abiotic stress enhanced tolerance by trehalose biosynthesis. Plant Sci., 169: 75-82.
Direct Link - Feng, X., P. Zhao, J. Hao, J. Hu, D. Kang and H. Wang, 2011. Effects of sorbitol on expression of genes involved in regeneration of upland rice (Oryza sativa L.). Plant Cell Tissue Organ Cult., 106: 455-463.
CrossRefDirect Link - Garcia, A.B., J.A. Engler, S. Iyer, T. Gerats, M. van Montagu and A.B. Caplan, 1997. Effects of osmoprotectants upon NaCl stress in rice. Plant Physiol., 115: 159-169.
Direct Link - Garg, A.K., J.K. Kim, T.G. Owens, A.P. Ranwala, Y.D. Choi, L.V. Kochian and R.J. Wu, 2002. Trehalose accumulation in rice plants confers high tolerance levels to different abiotic stresses. Proc. Natl. Acad. Sci. USA., 99: 15898-15903.
CrossRefDirect Link - Ge, L.F., D.Y. Chao, M. Shi, M.Z. Zhu, J.P. Gao and H.X. Lin, 2008. Overexpression of the trehalose-6-phosphate phosphatase gene OsTPP1 confers stress tolerance in rice and results in the activation of stress responsive genes. Planta, 228: 191-201.
CrossRefPubMedDirect Link - Hagedorn, C.H. and J.M. Phang, 1986. Catalytic transfer of hydride ions from NADPH to oxygen by the interconversions of proline and delta1-pyrroline-5-carboxylate. Arch. Biochem. Biophys., 248: 166-174.
CrossRefDirect Link - Hare, P.D., W.A. Cress and J. van Staden, 1998. Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ., 21: 535-553.
CrossRefDirect Link - Hassan, M., Z. Ahmed, M. Munir, S.I. Malik and K. Shahzad, 2009. Effect of sorbitol in callus induction and plant regeneration in wheat. Afr. J. Biotechnol., 8: 6529-6535.
Direct Link - Hu, Y. and U. Schmidhalter, 2005. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J. Plant Nutr. Soil Sci., 168: 541-549.
CrossRefDirect Link - Hellman, H., D. Funk, D. Rentsch and W.B. Frommer, 2000. Hypersensitivity of an Arabidopsis sugar signaling mutant toward exogenous proline application. Plant Physiol., 123: 779-790.
CrossRef - Jain, M., S. Tiwary and R. Gadre, 2010. Sorbitol-induced changes in various growth and biochemical parameters in maize. Plant Soil Environ., 56: 263-267.
Direct Link - Jang, I.C., S.J. Oh, J.S. Seo, W.B. Choi and S.I. Song et al., 2003. Expression of a bifunctional fusion of the Escherichia coli genes for trehalose-6-phosphate synthase and trehalose-6-phosphate phosphatase in transgenic rice plants increases trehalose accumulation and abiotic stress tolerance without stunting growth. Plant Physiol., 131: 516-524.
CrossRefDirect Link - Kadota, M., K. Imizu and T. Hirano, 2001. Double-phase in vitro culture using sorbitol increases shoot proliferation and reduces hyperhydricity in Japanese pear. Sci. Horticult., 89: 207-215.
CrossRefDirect Link - Karim, S., H. Aronsson, H. Fricson, M. Pirthonen and B. Leyman et al., 2007. Improved drought tolerance without undesired side effects in transgenic plants producing trehalose. Plant Mol. Biol., 64: 371-386.
Direct Link - Khedr, A.H.A., M.A. Abbas, A.A. Abdel Wahid, W.P. Quick and G.M. Abogadallah, 2003. Proline induces the expression of salt-stress-responsive proteins and may improve the adaptation of Pancratium maritimum L. to salt-stress. J. Exp. Bot., 54: 2553-2562.
CrossRefDirect Link - Li, H.W., B.S. Zang, Z.W. Deng and X.P. Wang, 2012. Overexpression of the trehalose-6-phosphate synthase gene OsTPS1 enhances abiotic stress tolerance in rice. Planta, 234: 1007-1018.
CrossRefDirect Link - Llorente, B.E., L.M. Juarez and N.M. Apostolo, 2007. Exogenous trehalose affects morphogenesis In vitro of jojoba. Plant Cell Tissue Organ Cult., 89: 193-201.
CrossRefDirect Link - Moustakas, M., I. Sperdouli, T. Kouna, C.I. Antonopoulou and I. Therios, 2012. Exogenous proline induces soluble sugar accumulation and alleviates drought stress effects on photosystem II functioning of Arabidopsis thaliana leaves. Plant Growth Regul., 65: 315-325.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Ozden, M., U. Demirel and A. Kahraman, 2009. Effects of proline on antioxidant system in leaves of grapevine (Vitis vinifera L.) exposed to oxidative stress by H2O2. Sci. Hort., 119: 163-168.
CrossRef - Posmyk, M.M. and K.M. Janas, 2007. Effects of seed hydropriming in presence of exogenous proline on chilling injury limitation in Vigna radiata L. seedlings. Acta Physiol. Plant, 29: 509-517.
CrossRefDirect Link - Pellny, T.K., O. Ghannoum, J.P. Conroy, H. Schluepmann and S. Smeekens et al., 2004. Genetic modification of photosynthesis with E. coli genes for trehalose synthesis. Plant Biotechnol. J., 2: 71-82.
CrossRefDirect Link - Sairam, R.K. and A. Tyagi, 2004. Physiology and molecular biology of salinity stress tolerance in plants. Curr. Sci., 86: 407-421.
Direct Link - Solomon, A., S. Beer, Y. Waisel, G.P. Jones and L.G. Paleg, 1994. Effects of NaCl on the carboxylating activity of Rubisco from Tamarix jordanis in the presence and absence of proline-related compatible solutes. Physiol. Plant., 90: 198-204.
Direct Link - Turkan, I. and T. Demiral, 2009. Recent developments in understanding salinity tolerance. Environ. Exp. Bot., 67: 2-9.
CrossRefDirect Link - Unno, H. and Y. Maeda, 2008. Effect of exogenous application of sugars on the salt tolerance of perennial ryegrass protoplasts. Biologia, 63: 204-206.
CrossRefDirect Link - Umar, S., I. Diva, N.A. Anjum, M. Iqbal, I. Ahmad and E. Pereira, 2011. Potassium-induced alleviation of salinity stress in Brassica campestris L. Central Eur. J. Biol., 6: 1054-1063.
CrossRefDirect Link - Van Rensburg, L., G.H.J. Kruger and H. Kruger, 1993. Proline accumulation the drought-tolerance selection: Its relationship to membrane integrity and chloroplast ultra structure in Nicotiana tabacum L. J. Plant Physiol., 141: 188-194.
CrossRefDirect Link - Venekamp, J.H., 1989. Regulation of cytosol acidity in plants under conditions of drought. Physiologia Plantarum, 76: 112-117.
CrossRefDirect Link - Zeid, I.M., 2009. Trehalose as osmoprotectant for maize under salinity-induced stress. Res. J. Agric. Biol. Sci., 5: 613-622.
Direct Link