
Pradeep Kumar Patel
Department of Plant Physiology, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi-221005, India
A. Hemantaranjan
Department of Plant Physiology, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi-221005, India
Asian Journal of Crop Science
Year: 2012 | Volume: 4 | Issue: 3 | Page No.: 86-102







ABSTRACT
An experiment was conducted to investigate the possible involvement of exogenously applied salicylic acid (presowing seed soaking at the rate of 1.0 and 1.5 mM salicylic acid (SA) on dry matter production and its partitioning, antioxidant (proline and sugar), antioxidant enzymes i.e., superoxide dismutase, peroxidase and yield of four chickpea (Cicer arietinum L.) genotypes, viz., Tyson, ICC 4958, JG 315 and DCP 92-3 under drought stress. Drought imposed (by withholding irrigation) at 50% flowering and 50% podding phase of development. Results indicate wide variation in tolerance to drought stress amongst chickpea genotypes at both the critical stages i.e., 50% flowering and 50% podding. However, the yield and its components were higher in ICC 4958 as compared to other genotypes. It is inferred that the significant response of applied SA at the rate of 1.5 mM under drought condition was relatively more in ICC 4958 genotype in terms of antioxidant response; substantially alleviated drought induced adverse effect and yield. It is also conclude that drought stress at 50% flowering was found to be more damaging than the 50% podding phase.
PDF Abstract XML References Citation
Received: October 26, 2011;
Accepted: December 29, 2011;
Published: February 23, 2012
How to cite this article
Pradeep Kumar Patel and A. Hemantaranjan, 2012. Salicylic Acid Induced Alteration in Dry Matter Partitioning, Antioxidant Defence System and Yield in Chickpea (Cicer arietinum L.) under Drought Stress. Asian Journal of Crop Science, 4: 86-102.
DOI: 10.3923/ajcs.2012.86.102
URL: https://scialert.net/abstract/?doi=ajcs.2012.86.102
DOI: 10.3923/ajcs.2012.86.102
URL: https://scialert.net/abstract/?doi=ajcs.2012.86.102
INTRODUCTION
Drought is a major abiotic stress limiting agricultural crop production worldwide (Farooq et al., 2009a). Plants respond to water deficit and acclimate to drought stress through various physiological and biochemical changes (Farooq et al., 2009a, b). Drought induces generation of reactive oxygen species, such as superoxide radical (O2•), hydroxy radical (•OH), hydrogen peroxide (H2O2) and alkoxy radical (RO•) in chloroplasts, mitochondria and peroxisomes (Munne-Bosch and Penuelas, 2003; Esfandiari et al., 2008). These reactive oxygen species can directly attack membrane lipids and increase the lipid peroxidation, inactivate metabolic enzymes and damage the nucleic acids-leading to cell death (Mittler, 2002).
Plants have evolved a series of enzymatic and nonenzymatic antioxidant systems to cope with drought stress (Farooq et al., 2009a, b). Without this defence mechanism, plants could not efficiently convert solar energy into chemical one. Maintaining a high level of antioxidative enzyme activities may thus contribute to drought tolerance (Sharma and Dubey, 2005). Enhanced superoxide dismutase and peroxidase activities have been associated with induced resistance of plants to drought stress (Bartels, 2001). Activities of both are involved in superoxideradical and hydrogen peroxide scavenging. Grain legumes, as compared to cereals, appear to have more sensitivity towards drought.
Previous reports have indicated adverse effects of drought on chickpea (Kumar and Abbo, 2001). Chickpea is a cool-season legume of northern region of India, which is also being cultivated in warm season environment of central and southern parts of the country. Due to changing climate, yield losses might be the result of intermittent drought during the vegetative phase or due to drought during reproductive development is expected to increase leading to reduction in its potential yield. As there is reduction in the translocation of carbohydrates during drought stress, this leads to a change in source-sink relationships. Thus, effective measures are needed to counter the negative effects of drought on this crop. One of the effective measures in this direction is the application of certain plant growth regulators for mitigating those effects.
Salicylic acid is a naturally existing phenolic compound and is considered to be a potent plant growth regulator because of its diverse regulatory role in plant metabolism. Evidences put forward that externally applied salicylic acid increased plant’s tolerance to several abiotic stresses, including osmotic stress (Wang et al., 2010), drought (Azooz and Youssef, 2010), heavy metal stress (Moussa and El-Gamal, 2010) and also influence a range of diverse processes in plants, including seed germination, stomatal closure, ion uptake and transport, membrane permeability, photosynthesis and plant growth rate (Aftab et al., 2010). Salicylic acid treatment cause decrease malondialdehyde content (MDA) while membrane stability was improved (Kazemi et al., 2011). Chickpea shows mechanisms for overcoming this condition with the help of salicylic acid.
The present investigation was carried out with a view to recognize effect of presowing seed soaking of salicylic acid for dry matter gain in shoots and its partitioning to different plant parts besides alteration in antioxidant defence system and yield variations under drought stress in four chickpea (Cicer arietinum L.) genotypes.
MATERIALS AND METHODS
The experiment was carried out during Rabi season 2009-10 in rain- protected wire- house at the Horticulture Research Farm, Institute of Agricultural Sciences, Banaras Hindu University Varanasi, India. The experimental area lies between latitudes 25.18°N, longitude 83.03°E and 123.93 m altitude. The average of climatic conditions calculated during the entire growth period is as follows: maximum/minimum temperature, Relative Humidity (RH) was 28.0/13.6°C, 71.3/36.5%, respectively and an average sunshine hour was 6.9.
The seeds of Cicer arietinum L. genotypes (Tyson, ICC 4958, JG 315 and DCP 92-3) were surface sterilized with 0.2% HgCl2 solution. Salicylic acid (Molecular Weight: 138.12 Sigma Aldrich, Chemie GmBH, Munich, Germany) was dissolved in absolute ethanol then added drop wise to water (ethanol/water: 1/1000 v/v) (Williams et al., 2003). Thereafter 10 seeds of each genotype for each treatment were soaked for 6 h in distilled water without salicylic acid (0 mM salicylic acid) taken as control (T0) and in 1.0 mM (T1) and 1.5 mM salicylic acid (T2) before sowing in pots (30 cm in diameter and 30 cm in depth) containing farm soil having 12-14% moisture at the time of sowing and were thinned to six plants per pot at the first true leaf stage. The experimental soil was sandy loam containing organic carbon 0.31%, available nitrogen 228.00 kg ha-1, available phosphorus 17.00 kg ha-1, available potassium 180.00 kg ha-1 and pH 7.3. There were 36 pots per treatment and the three replications of each experimental unit. All the pots were applied with measured dose of fertilizer for chickpea at the rate of 20, 40, 20 kg ha-1 of N, P2O5 and K2O, respectively.
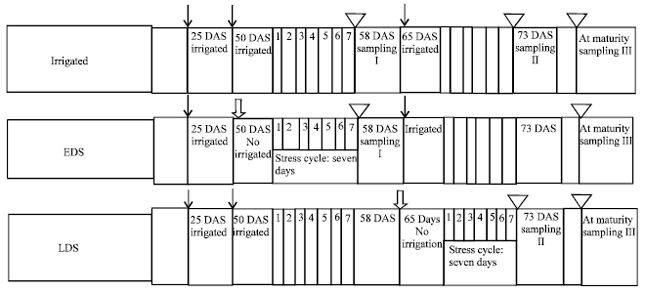 | |
| Fig. 1: | Schematic representation of experimental layout, ↓ (Solid arrow) indicate the irrigated position with respect to DAS, |
Drought stress applications: Each genotype was grouped in three sets e.g., irrigated, drought imposed at 50% flowering (i.e., early drought stress) and 50% podding (i.e., late drought stress) stage. Drought stress treatment was initiated at the early and late stage by controlling irrigation schedule and it was instigated at Days After Sowing 50 (DAS). control plants (irrigated) were given three irrigations (at 25, 50 and 65 DAS) from the date of sowing to maturity. Early Drought Stressed (EDS) received two irrigations (25 and 65 DAS) whereas Late Drought Stressed (LDS) received two irrigations at 25 and 50 DAS, stress cycle given seven days in both the conditions. Induced drought stress measured in terms of leaf Relative Water Content (RWC). RWC decreases from 1st days to 7th day of stress cycle, on the 7th day Relative water contant is varying from 67.96-71.67% in EDS and 69.0-71.17% at LDS state among chickpea (Cicer arietinum L.) genotypes (Fig. 1).
Methods: Dry matter production and partitioning expressed as g plant-1 were observed at 50% flowering and 50% podding stages. For sampling the normal and stressed plants were pulled out at each sample time and were separated into stem, leaves, flowers and pods and dried in oven at 105°C for 1 h followed by at 65°C till constant weight, to record dry weights. Total shoot dry matter and seed yield Plant-1 were recorded at maturity.
Leaf area (cm2 plant-1) was measured by leaf area meter (Model Systronics Leaf Area Meter-211). Relative Water Content Relative water contant of the leaves was estimated according to the method of Weatherly (1950). Total soluble sugar determined by anthrone method (Dubois et al., 1956). Proline content was determined by the method of Bates et al. (1973). The activity of (Super oxide dismutase) (EC 1.15.1.1) was assayed by measuring its ability to inhibit the photochemical reduction of nitroblue tetrazolium (Dhindsa et al. 1981). The assay of peroxidase (EC 1.11.1.7) activity was carried out by measuring the decrease in absorbance at 420 nm due to decomposition of H2O2 (Kar and Mishra, 1976).
Statistical analysis: Samples were collected in three replicates and each replicate/sample was assayed twice. The design of the experiment was CRD and data was analyzed for analysis of variance (ANOVA) and means were compared by the Least Significant Difference (LSD) test and those at p<0.05. Standard error of mean was also calculated (Gomez and Gomez, 1984).
RESULTS AND DISCUSSION
Phenology has a powerful effect on plant growth response and productivity under drought stress. Growth duration determines water-requirement and the probability of exposure to stress, both of which decrease in early flowering genotypes (Blum, 1996). In this experiment salicylic acid treatment is nonsignificant in respect to phenology of crop. The days to Ist flowering in chickpea genotypes ranged minimum from 45 days (ICC 4958) to maximum 53 days (Tyson). Days to 50% flowering in various chickpea genotypes ranged minimum from 59 days (Tyson) to maximum 62 days (JG 315), whereas in highest seed yielding genotype ICC 4958 it was 60 days. Days to 50% podding and maturity among the genotypes ranged minimum from 74 day and 110 days (ICC 4958) to a maximum of 85 and 125 days (DCP 92-3), respectively. The highest seed yielder ICC 4958 took 74 days for 50% podding and 110 days for maturity and thus minimizing the impact of drought stress as terminal stress predominates in the region (Fig. 2).
Significant differences amongst genotypes in leaf area (cm2 plant-1) at 50% flowering were observed (Table 1). Leaf area gradually increased to maximum at 50% podding (Table 2) and then declined at maturity. Amongst genotypes, highest leaf area was noticed in DCP 92-3 while it was lowest in ICC 4958 and Tyson both at 50% flowering and 50% podding. The highest seed yielding genotype ICC 4958 attained leaf area of 161.65 at 50% flowering and 207.58 cm2 palnt-1 at 50% podding. Genotype DCP 92-3 attained highest leaf area but not produced highest yield. Plasticity in leaf area is an important means by which drought-stressed crop maintains control over water-use (Blum, 1996). The results suggest that crops need to adjust their transpiring surface through reducing leaf growth (as a mechanism for reducing water loss). Thus, drought tolerance in this study may be attributed to reduced transpiration and then water loss by having smaller leaf size and reducing leaf area expansion of genotypes when drought stress develops.
In grain legumes, dry matter production and its accumulation is related to the flowering time. Chickpea being a long day plant, delayed flowering that increased dry matter accumulation, whereas conditions favouring early flowering adversely affected dry matter accumulation. Tatar and Gevrek (2008) reported that onset of water stress at early phase of development greater decline of dry matter production than the late phase. Chickpea genotypes Tyson, JG 315 and DCP 92-3 exhibited slow rate of dry matter accumulation up to initiation of flowering except ICC 4958 due to lesser leaf area at this stage. Total dry matter accumulation in chickpea increased with the onset of plant growth showing maximum in between flowering and podding in all chickpea genotypes. Stem dry matter (g plant-1) recorded at 50% flowering, 50% podding and at maturity that showed significant differences among genotypes (Table 1-3). Dry matter of stem gradually increased till maturity. Further, maximum dry matter at 50% flowering was noticed in Tyson (1.52 g) followed by DCP 92-3 (1.02 g) and ICC 4958 (1.03 g), which was minimum in JG 315 (0.93 g). The percentage dry matter reduction in stressed plants was noticed to a maximum in Tyson (52.4%) over normal at 50% flowering followed by ICC 4958 (51.4%) in treatment T2 (1.5 mM SA). Subsequently, at 50% podding it was again uppermost in Tyson (2.35 g) closely followed by DCP 92-3 (2.23 g) while it was lowest in JG 315 (1.66 g). As far as percentage reduction of stem dry matter is concerned, it was maximum under stress over normal in ICC 4958 (59.3%) followed by Tyson (52.6%) at 50% podding (Table 2). Tyson (52.6%) at 50% podding (Table 2).
| Table 1: | Effect of Salicylic Acid (SA) on plant height (cm), leaf area (cm2) and dry weight (g) in chickpea (Cicer arietinum L.) genotypes under drought stress imposed at 50% flowering stage |
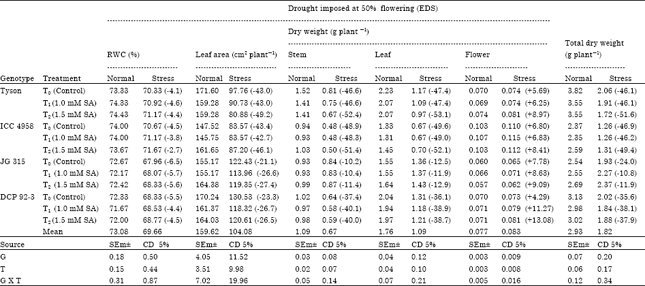 | |
| Genotypes (G), Treatments (T), Values in parenthesis indicate % increase (+), or decrease (-) under drought stress condition over normal, (EDS: Early drought stress or drought stress 50% flowering) | |
| Table 2: | Effect of salicylic acid on RWC, leaf area, dry weight (stem, leaf, flower, pod and total) in chickpea (Cicer arietinum L.) genotypes under drought stress imposed at 50% podding stage |
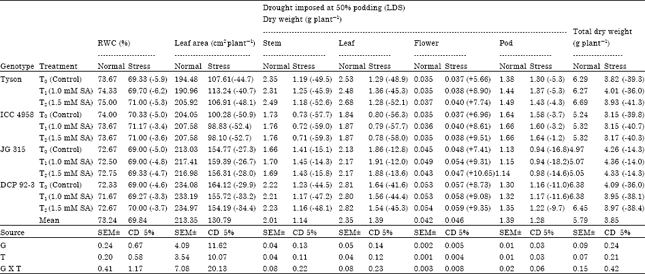 | |
| Genotypes (G), Treatments (T), Values in parenthesis indicate % increase (+), or decrease (-) under drought stress condition over normal, (LDS: Late drought stress or drought stress at 50% podding) | |
| Table 3: | Effect of salicylic acid on seed yield, 100 seed weight and biological yield in chickpea (Cicer arietinum L.) genotypes under drought stress imposed at early and late stage of development |
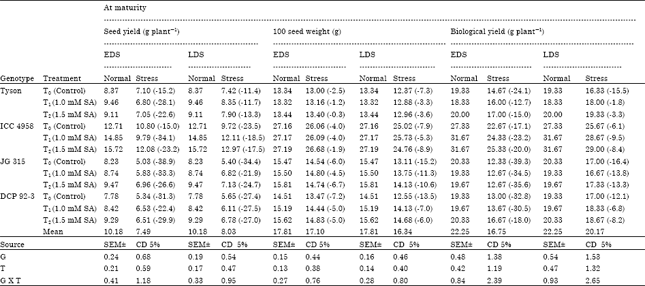 | |
| Genotypes (G), Treatments (T), Values in parenthesis indicate % increase (+), or decrease (-) under drought stress condition over normal, (EDS: Early drought stress or drought stress 50% flowering, LDS: Late drought stress or drought stress at 50% podding) | |
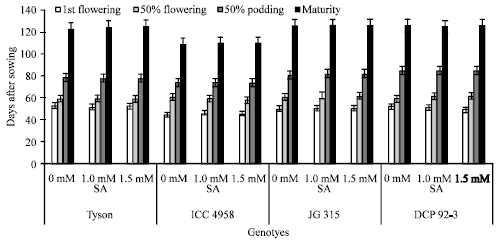 | |
| Fig. 2: | Phenological traits of chickpea (Cicer arietinum L.) genotypes. Data are presented as treatments Mean±SE |
The significant response of the threshold level of salicylic acid at the rate of 1.5 mM was relatively more in ICC 4958 and Tyson. Results also point towards allocation/partitioning of dry matter to different plant parts for growth and maintenance in relation to salicylic acid.
Leaf dry matter (g plant-1) observed at 50% flowering, 50% podding showed significant difference amongst genotype (Table 1, 2). Leaf dry matter increased gradually to maximum at 50% podding and thereafter showed decrease till maturity. Maximum leaf dry matter amongst genotypes at 50% flowering was noticed in Tyson (2.23 g) followed by DCP 92-3 (2.04 g) while it was minimum ICC 4958 (1.31 g) here salicylic acid treatment is nonsignificant amongst genotypes. Further, maximum leaf dry matter at 50% podding was noticed in DCP 92-3 (2.82 g) followed by Tyson (2.68 g) in T2 while it was lowest in ICC 4958 (1.84 g). Flower dry weight (g plant-1) at 50% flowering had significant difference among genotypes. ICC 4958 (0.115 g) showed maximum Pod dry weight (g plant-1) at 50% podding had significant differences amongst genotypes and SA treatment (Table 2), showed maximum in the highest seed yielding genotype ICC 4958 (1.66 g) followed by Tyson (1.49 g) in T1, while in T0 it was lowest in JG 314 (1.13 g). The difference in dry matter accumulation could be related to the difference in phenology and toward the late stages it might also be related to differing pattern of assimilate partitioning between vegetative and reproductive component (Bell et al., 1991).
Adopting Levitt (1972) definition, the capability of genotypes to maintain relatively higher water content is an avoidance mechanism. When plant adjusts to lower relative water content, it is considered a tolerance mechanism. Drought stress significant reduces the relative water content at both the critical phase of development (Tatar and Gevrek, 2008; Pattanagul, 2011). In the present study, Relative water contant was low at 50% flowering compared to 50% podding in all the genotypes at drought stress conditions. Genotypes JG 315 and DCP 92-3 showed lower Relative water contant as compared to Tyson and ICC 4958 at 50% flowering stage. Tyson and ICC 4958 had maintained higher RWC at both the stages. Relative water contant (%) was in the range of 71.6-74.4 (unstressed) and 67.9-71.6 (stressed), whereas salicylic acid at the rate of 1.5 mM increased leaf Relative water contant values close to the control (unstressed plant). Results showed lowest RWC value (68%) in genotype JG 315 and the maximum RWC (71.17%) was remarkably maintained under stress in Tyson (Table 1). The higher seed yielding genotype generally maintained more than 70% leaf relative water content. Maintenance of plant water status is a fundamental phenomenon for normal growth of plants under stressful environment. Disturbances in water balance in plants lead to impaired functioning of different gas exchange attributes, ultimately resulting in reduced plant growth (Campos et al., 1999; Demirevska et al., 2010; Rapacz et al., 2010). Under such conditions, exogenous application of salicylic acid helps regulate the plant water relation. It appears that salicylic acid potentially generates a wide array of metabolic responses in plants and also effective for plant water relations (Hayat et al., 2010). Exogenous application of salicylic acid may help reduce adverse effects of drought in chickpea. Change in Relative water contant with alternate decrease and increase during the period of water stress suggests alteration in internal mechanisms for osmotic adjustments that helped to maintain Relative water contant closer to normal pre stress value and thereby preventing Relative water contant to fall below a critical level especially in Tyson genotype.
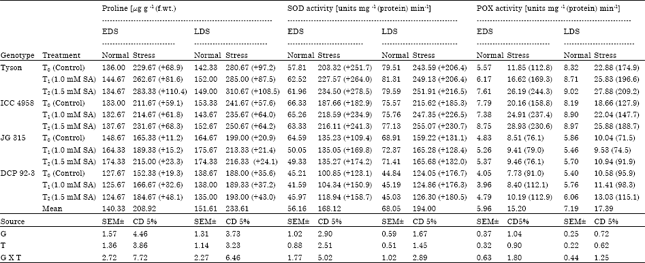
Results show that proline accumulation was increased at 50% flowering and 50% podding stages under drought with higher accumulation at 50% podding stage. As far as genotypes are concerned, Tyson and ICC 4958 accumulated more proline as compared to JG 315 and DCP 92-3 in response to salicylic acid over the control. Proline content in plants treated with salicylic acid at the rate of 1.5 mM under drought stress was maximum [310.67 μg g-1 (f.wt.)] in Tyson and minimum in DCP 92-3 [188.0 μg g-1 (f.wt.)] (Table 4). There was a strong correlation between plant water content and accumulation of compatible solutes (proline and glycinebetaine) under drought stress. Costa et al. (2008) reported that the plant submitted to stress suffered an increase in amount of proline, since the solute is an osmotic adjuster. Salicylic acid treatment obviously improved Relative water contant with simultaneous and significant increase in proline resulted in osmotic adjustment to a great extent. In this study, one of the important osmolytes, proline, was substantially increased upon exposure to drought; interestingly, salicylic acid could have induced abscisic acid mediated protective reactions of plants to water deficit mainly by increasing proline accumulation (Yoshiba et al., 1995) which along with osmolytes has improved dehydration tolerance. Azooz and Youssef (2010) reported proline content increase with application of salicylic acid under drought stress. The increased proline might have helped to maintain high tissue water contents as indicated by higher values of RWC (Table 1).
Total soluble sugar level differed significantly at 50% flowering and podding stages among chickpea genotypes with salicylic acid treated plant under drought. In this experiment, Tyson and ICC 4958 showed more soluble sugar contents under drought stress (Fig. 3), which was further increased by salicylic acid in all four genotypes (Fig. 3). Al-Hakimi (2006) previously reported in soybean plants increase the soluble sugar content exhibited a favourable effect of the accumulation along with some ions antagonized or ameliorated the inhibitory effect of drought stress by application of salicylic acid. The oxidative stress may damage the biological membranes thus disrupting the whole assimilation process (Farooq et al., 2009c) and resulting in substantial yield decline, whereas reduction in tissue water status disrupts the activities of several enzymes involved in carboxylation and antioxidant defence system (Farooq et al., 2009a, b) thus leading to yield reduction. Cytosolic concentration of osmolytes (i.e., soluble sugar and proline) is often increased resistance to drought stress, which not only helps in maintaining the tissue but also is involved in osmoregulation (Lobato et al., 2008; Farooq et al., 2009a, b; Pattanagul, 2011; Nahar et al., 2011). Azooz and Youssef (2010) and Ismail et al. (2011) recently reported soluble carbohydrates increase with application of salicylic acid. Soluble sugar might have functioned as a typical osmoprotectant for osmotic adjustment in chickpea in order to stabilize cellular membranes for maintaining turgor pressure.
| Table 4: | Effect of salicylic acid on proline, superoxide dismutase (SOD) activity and peroxidase (POX) activity in chickpea (Cicer arietinum L.) genotypes grown under drought stress imposed at early and late stage of development |
 | |
| Genotypes (G), Treatments (T), Values in parenthesis indicate % increase (+), or decrease (-) under drought stress condition over normal, (EDS: Early drought stress or drought stress 50% flowering, LDS: Late drought stress or drought stress at 50% podding) | |
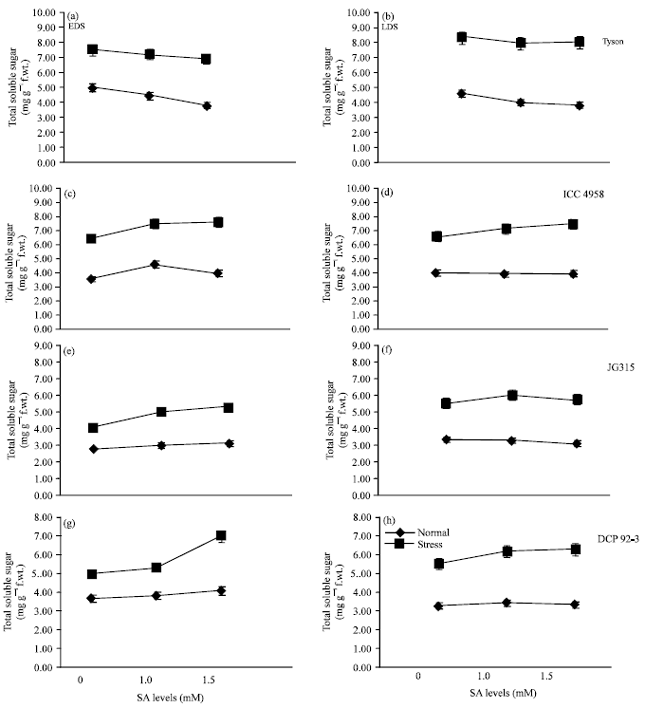 | |
| Fig. 3(a-h): | Effect of Salicylic Acid (SA) on total soluble sugar [mg g-1 (f.wt.)] in chickpea (Cicer arietinum L.) genotypes grown under drought stress imposed at early and later stage of development. (i.e., EDS: Early drought stress or drought stress 50% flowering, LDS: Late drought stress or drought stress at 50% podding). Data are presented as treatments Mean±SE |
Thus, it is evident that osmotic adjustment prevents lowering of Relative water contant below a critical level by efficient water uptake, restores photosynthesis and maintains positive carbon balance for longer stress period.
Under stress conditions, the generation of Reactive oxygen species is a common phenomenon observed in most plants. These stress-generated Reactive oxygen species damage the macromolecules such as DNA, proteins and lipid structures (Foyer and Noctor, 2003; Ashraf and Foolad, 2007; Ashraf, 2009). Plants have an internal protective enzyme catalysed clean up system to scavenge reactive oxygen species, thus ensuring normal cellular function. Superoxide dismutase constitutes the first line of defence via detoxification of super oxide radicals (Sairam et al., 2000), thereby maintaining membranes of plant tissue. Superoxide dismutase detoxifies superoxide anion free radicals by forming H2O2 it can be further eliminated by concerted action of peroxidase. In this study, a steady increase in the activities of superoxide dismutase and peroxidase was observed with time (Table 4). Super oxide dismutase showed maximum response to Salicylic acid. An increase in activity of peroxidase was observed, though not as marked as for superoxide dismutase. Superoxide dismutase and peroxidase activity increased significantly at the rate of 1.5 mM salicylic acid at drought imposed 50% podding stage (Table 4). This well elucidates that peroxidase should play a more significant role in detoxifying the produced H2O2 since the activity of peroxidase increased, in contrast to that of other antioxidant enzymes i.e., CAT. Besides, it is well known that CAT is less efficient than peroxidase in scavenging H2O2 because of its low substrate affinity (Erdal and Dumlupinar, 2010a, b). Tyson and ICC 4958 showed higher activities of superoxide dismutase and peroxidase than JG 315 and DCP 92-3 at both the stage and under all the treatments. Salicylic acid at the rate of 1.0 and 1.5 mM also significantly increased antioxidant enzymatic activities. However, activity was higher with salicylic acid at the rate of 1.5 mM under drought condition and genotypes (Tyson and ICC 4958) as compared to salicylic acid at the rate of 1.0 mM. Genotype Tyson nonsignificantly differed from ICC 4958 but it differed significantly both from JG315 and DCP 92-3. At 50% podding superoxide dismutase activity was higher at the rate of 1.5 mM SA [255.07 units mg-1 (protein) min-1] than 1.0 mM SA [247.35 units mg-1 (protein) min-1] in genotype ICC 4958 (Table 4) whereas, peroxidase activity was similar with respect to treatment but differed in genotypes. Highest values of peroxidase activity were observed at 1.5 mM SA in ICC 4958 [28.97 unit mg-1 (protein) min-1] (Table 4).
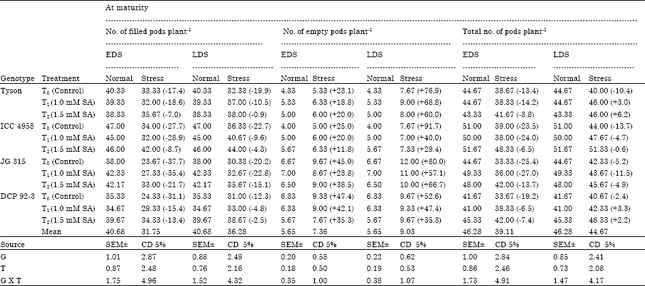
There were significant differences among the genotypes and treatments for the total number of filled pods plant-1 and seed yield plant-1 along with 100 seed weight. Maximum in numbers were observed in the genotype ICC 4958 (46.00, 15.72 and 27.19) followed by genotype JG 315 (42.17, 9.47 and 15.81), respectively in the treatment of salicylic acid at the rate of 1.5 mM. Empty pods plant-1 was significantly different among genotypes but nonsignificant with treatments. Maximum number of empty pods plant-1 was noticed in JG 315 (12.00) and minimum in ICC 4958 (7.00) under drought (Table 5).
The present investigation on the accumulation and partitioning of dry matter of chickpea crop exhibited specific trend about productivity potential that may prove helpful to achieve better criteria for yield improvement. The chickpea genotype ICC 4958 performed better than other genotype due to higher number of filled pods, seed yield plant-1, higher RWC. The higher relative water content indicated a better osmoregulation in these genotypes. The genotypes ICC 4958 also showed the characteristics of earliness by maturing in 110 days after sowing under drought.
Data supports that increases the activity of antioxidant enzymes such as superoxide dismutase and peroxidase (Singh and Usha, 2003; Hayat et al., 2008), which in turn protect plants against reactive oxygen species generation and membrane injury or may affect synthesis of other substances having a protective effect on plants under stress. Besides these, salicylic acid might have helped in regulating the stomatal functioning and continued the water status of plant under water stress, which in turn maintained various physiological processes needed for increased growth and yield.
In this investigation, salicylic acid not only alleviated the inhibitory effects of drought stress but also had a stimulatory effect in the dry matter gain in the shoots and its partitioning to different plant parts.
| Table 5: | Effect of salicylic acid on filled pods, empty pods and total pods plant-1 in chickpea (Cicer arietinum L.) genotypes under drought stress imposed at early and late stage of development |
 | |
| Genotypes (G), Treatments (T), Values in parenthesis indicate % increase (+), or decrease (-) under drought stress condition over normal, (EDS: Early drought stress or drought stress 50% flowering, LDS: Late drought stress or drought stress at 50% podding) | |
There is evidence that GA regulates the dry matter partitioning between source and sink. Noushina et al. (2011) recently reported that GA signalling is involved in adjustment of plants under limiting environmental conditions and maintains source-sink relation. This adjustment could be mediated through the GA and salicylic acid interaction. Hence, there is every reason that salicylic acid treatment under drought condition had maintained source-sink relationship in the present experiment to improve yield and yield attributes by dry matter partitioning. Genotype Tyson and ICC 4958 substantiated relatively more tolerant to drought than DCP 92-3 and JG 315. However, the response of exogenously applied salicylic acid was more pronounced in Tyson and ICC 4958.
On the whole, observations it was point out that accumulation of osmolytes allows additional water to be taken up from the environment reducing the immediate effect of water shortage within the plant so as to help stabilizing protein tertiary structure and cells. For that reason, seed soaking treatment of salicylic acid is beneficial to improve antioxidant system in organizing overall physiology under drought.
CONCLUSIONS
Based on analyses of four chickpea genotypes and two salicylic acid treatments, there were substantial variations in biomass partitioning in obtaining yield under drought within chickpea genotypes. A perusal of results shows that drought tolerant genotypes ICC 4958 have higher Relative water contant, less leaf area higher, total soluble sugars, proline accumulation and enzymatic activities such as superoxide dismutase and peroxidase in comparison to drought susceptible genotypes. Dry matter partitioning is more to different plant parts for growth and maintenance under drought. The significant response of the threshold level of salicylic acid at the rate of 1.5 mM was, relatively more in ICC 4958 and Tyson. On the basis of the performance of chickpea genotypes at different levels of salicylic acid especially at 50% flowering and 50% podding stages of development, it is concluded that 50% flowering stage was most sensitive under drought stress, which could be in part mitigated by pre-soaking salicylic acid treatment for improved yield and yield attributes.
ACKNOWLEDGEMENT
We extend our sincere thanks to the University Grant Commission (UGC) for research fellowship for Ph.D. Programme and the Indian Institute of Pulse Research (IIPR- ICAR) Kanpur, India for their kind providing of chickpea genotypes through material transfer agreement.
REFERENCES
- Lobato, A.K.S., C.F.O. Neto, R.C.L. Costa, B.G.S. Filho, F.J.R. Cruz and H.D. Laughinghouse, 2008. Biochemical and physiological behavior of Vigna unguiculata (L.) Walp. Under water stress during the vegetative phase. Asian J. Plant Sci., 7: 44-49.
CrossRefDirect Link - Ismail, A.M., M.F. Abou Alhamd, H.R.M. Galal and F.A. Nasr-Eldeen, 2011. Modification of photosynthetic pigments, osmotic solutes and ions accumulation in Chlorella vulgaris and Wheat Cv. Sds-1 seedlings under the influence of NaCl with salicylic acids. Res. J. Bot., 6: 100-111.
CrossRef - Al-Hakimi, A.M.A., 2006. Counteraction of drought stress on soybean plants by seed soaking in salicylic acid. Int. J. Bot., 2: 421-426.
CrossRefDirect Link - Aftab, T., M. Masroor, A. Khan, M. Idrees, M. Naeem and Moinuddin, 2010. Salicylic acid acts as potent enhancer of growth, photosynthesis and artemisinin production in Artemisia annua L. J. Crop Sci. Biotech., 13: 183-188.
CrossRefDirect Link - Ashraf, M. and M.R. Foolad, 2007. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot., 59: 206-216.
CrossRefDirect Link - Ashraf, M., 2009. Biotechnological approach of improving plant salt tolerance using antioxidants as markers. Biotechnol. Adv., 27: 84-93.
CrossRefPubMedDirect Link - Azooz, M.M. and M.M. Youssef, 2010. Evaluation of heat shock and salicylic acid treatments as inducers of drought stress tolerance in hassawi wheat. Am. J. Plant Physiol., 5: 56-70.
CrossRefDirect Link - Bartels, D., 2001. Targeting detoxification pathways: An efficient approach to obtain plants with multiple stress tolerance. Trends Plant Sci., 6: 284-286.
CrossRef - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Blum, A., 1996. Crop responses to drought and the interpretation of adaptation. Plant Growth Regul., 20: 135-148.
CrossRefDirect Link - Campos, P.S., J.C. Ramalho, J.A. Lauriano, M.J. Silva and M. do Ceu Matos, 1999. Effects of drought on photosynthetic performance and water relations of four Vigna genotypes. Photosynthetica, 36: 79-87.
CrossRefDirect Link - Demirevska, K., L. Simova-Stoilova, I. Fedina, K. Georgieva and K. Kunert, 2010. Response of oryzacystatin I transformed tobacco plants to drought, heat and light stress. J. Agron. Crop Sci., 196: 90-99.
CrossRefDirect Link - Dhindsa, R.S., P. Plumb-Dhindsa and T.A. Thorpe, 1981. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation and decreased levels of superoxide dismutase and catalase. J. Exp. Bot., 32: 93-101.
CrossRefDirect Link - DuBois, M., K.A. Gilles, J.K. Hamilton, P.A. Rebers and F. Smith, 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem., 28: 350-356.
CrossRefDirect Link - Esfandiari, E., M.R. Shakiba, S.A. Mahboob, H. Alyari and S. Shahabivand, 2008. The effect of water stress on the antioxidant content, protective enzyme activities, proline content and lipid peroxidation in wheat seedling. Pak. J. Biol. Sci., 11: 1916-1922.
CrossRefPubMedDirect Link - Farooq, M., A. Wahid and D.J. Lee, 2009. Exogenously applied polyamines increase drought tolerance of rice by improving leaf water status, photosynthesis and membrane properties. Acta. Physiol. Plant., 31: 937-945.
CrossRefDirect Link - Farooq, M., A. Wahid, N. Kobayashi, D. Fujita and S.M.A. Basra, 2009. Plant drought stress: Effects, mechanisms and management. Agron. Sustainable Dev., 29: 185-212.
CrossRefDirect Link - Farooq, M., A. Wahid, O. Ito, D.J. Lee and K.H.M. Siddique, 2009. Advances in drought resistance of rice. Crit. Rev. Plant Sci., 28: 199-217.
CrossRef - Foyer, C.H. and G. Noctor, 2003. Redox sensing and signalling associated with reactive oxygen in chloroplasts, peroxisomes and mitochondria. Physiol. Plant., 119: 355-364.
CrossRefDirect Link - Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedures for Agricultural Research. 2nd Edn., John Wiley and Sons Inc., Hoboken, New Jersey, ISBN: 978-0-471-87092-0, Pages: 704.
Direct Link - Hayat, Q., S. Hayat, M. Irfan and A. Ahmad, 2010. Effect of exogenous salicylic acid under changing environment: A review. Environ. Exp. Bot., 68: 14-25.
CrossRefDirect Link - Hayat, S., S.A. Hasan, Q. Farriduddun and A. Ahmad, 2008. Growth of tomato (Lycopersicon esculentum) in response to salicylic acid under water stress. J. Plant Interact., 3: 297-304.
CrossRefDirect Link - Nahar, K., S.M. Ullah and N. Islam, 2011. Osmotic adjustment and quality response of five tomato cultivars (Lycopersicon esculentum Mill) following water deficit stress under subtropical climate. Asian J. Plant Sci., 10: 153-157.
CrossRefDirect Link - Kar, M. and D. Mishra, 1976. Catalase, peroxidase and polyphenoloxidase activities during rice leaf senescence. Plant Physiol., 57: 315-319.
CrossRefDirect Link - Kumar, J. and S. Abbo, 2001. Genetics of flowering time in chickpea and its bearing on productivity in semiarid environments. Adv. Agron., 72: 107-138.
CrossRef - Kazemi, M., E. Hadavi and J. Hekmati, 2011. Role of salicylic acid in decreases of membrane senescence in cut carnation flowers. Am. J. Plant Physiol., 6: 106-112.
CrossRefDirect Link - Mittler, R., 2002. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci., 7: 405-410.
CrossRefPubMedDirect Link - Moussa, H.R. and S.M. El-Gamal, 2010. Effect of salicylic acid pretreatment on cadmium toxicity in wheat. Biol. Plant., 54: 315-320.
CrossRefDirect Link - Munne-Bosch, S. and J. Penuelas, 2003. Photo and antioxidative protection and a role for salicylic acid during drought and recovery in field-grown Phillyrea angustifolia plants. Planta, 217: 758-766.
PubMed - Iqbal, N., R. Nazar, M.I.R. Khan, A. Masood and N.A. Khan, 2011. Role of gibberellins in regulation of source-sinkrelations under optimal and limiting environmental conditions. Cur. Sci., 100: 998-1007.
Direct Link - Tatar, O. and M.N. Gevrek, 2008. Influence of water stress on proline accumulation, lipid peroxidation and water content of wheat. Asian J. Plant Sci., 7: 409-412.
CrossRefDirect Link - Costa, R.C.L., A.K.S. Lobato, C.F.O. Neto, P.S.P. Maia, G.A.R. Alves and H.D. Laughinghouse, 2008. Biochemical and physiological responses in two Vigna uguiculata (L.) walp. Cultivars under water stress. J. Agron., 7: 98-101.
CrossRefDirect Link - Rapacz, M., J. Koscielniak, B. Jurczyk, A. Adamska and M. Wojcik, 2010. Different patterns of physiological and molecular response to drought in seedlings of malt‐ and feed‐type barleys (Hordeum vulgare). J. Agron. Crop Sci., 196: 9-19.
CrossRefDirect Link - Sairam, R.K., G.C. Srivastava and D.C. Saxena, 2000. Increased antioxidant activity under elevated temperatures: A mechanism of heat stress tolerance in wheat genotypes. Biol. Plant., 43: 245-251.
CrossRefDirect Link - Sharma, P. and R.S. Dubey, 2005. Lead toxicity in plants. Braz. J. Plant Physiol., 17: 35-52.
CrossRefDirect Link - Singh, B. and K. Usha, 2003. Salicylic acid induced physiological and biochemical changes in wheat seedlings under water stress. Plant Growth Regul., 39: 137-141.
CrossRefDirect Link - Wang, L.J., L. Fan, W. Loescher, W. Duan, G.J. Liu and J.S. Cheng, 2010. Salicylic acid alleviates decreases in photosynthesis under heat stress and accelerates recovery in grapevine leaves. BMC Plant Biol., 10: 34-34.
CrossRef - Pattanagul, W., 2011. Exogenous abscisic acid enhances sugar accumulation in rice (Oryza sativa L.) under drought stress. Asian J. Plant Sci., 10: 212-219.
CrossRefDirect Link - Williams, M., T. Senaratna, K. Dixon and K. Sivasithamparam, 2003. Benzoic acid induces tolerance to biotic stress caused by Phytophthora cinnamomi in Banksai attenuate. Plant Growth Regul., 41: 89-91.
Direct Link - Weatherley, P.E., 1950. Studies in the water relations of the cotton plant. I. The field measurement of water deficits in leaves. New Phytol., 49: 81-97.
CrossRefDirect Link
BRAJESH KUMAR MISHRA Reply
Dear,
patel this paper explains involment of sa under drought stress is really a new and innovative approch to overcome the adverse effect of drought stress in chieckpea the very important crop of rainfed agriculture. Earlier papers of sa mostly pointed that it protect plants from biotic factors but your results shows that it is responsible in drought mitigation and can be used PGR in crop cultivation really it is innovative work.