
N. Rajashekar
Department of studies in Botany, University of Mysore, Manasagangotri, Mysore-570 006, India
Prakasha
Department of studies in Botany, University of Mysore, Manasagangotri, Mysore-570 006, India
T.C. Shivashankara Murthy
Department of studies in Botany, University of Mysore, Manasagangotri, Mysore-570 006, India
Asian Journal of Crop Science
Year: 2012 | Volume: 4 | Issue: 2 | Page No.: 80-85







ABSTRACT
In the present investigation five treatments with pendimethalin concentrations namely 0.5, 1.0, 2.5, 5.0 and 10.0 ppm were given to maize cv. NAC-6002 grains to evaluate herbicidal toxicity on germination and early growth. The seedlings were maintained upto 15 days. Embryonic axis and endosperm of 4, 6, 8, 10 and 15 days old germinated soybean seedlings were used for bioassay on biochemical parameters viz., total protein, total carbohydrate, starch and reducing sugar were carried out. Germination percentage, length of radical and plumule were decreased significantly with increase in concentration of pendimethalin. The germinating seedlings of treated set showed high content of seed serves viz., total protein, total carbohydrate, starch and reducing sugar in the endosperms, whereas it was low in the shoot-root axis at all the days of observation over control. With increase in the days of germination the seed serves (protein, carbohydrate, starch and reducing sugar) showed a drastic decrease in control, whereas in treated sets the rate of seed reserves degradation was slow in the endosperm. On the contrary, in shoot-root axis a reverse trend was recorded in response to pendiemthalin treatment. The effect was more pronounced at higher herbicidal concentration. These results reveal that the mobilization of seed reserve was suppressed by pendimethalin, as result of impaired degradation of seed reserve, during germination and early growth.
PDF Abstract XML References Citation
Received: August 05, 2011;
Accepted: November 29, 2011;
Published: January 21, 2012
How to cite this article
N. Rajashekar, Prakasha and T.C. Shivashankara Murthy, 2012. Seed Germination and Physiological Behavior of Maize (cv. Nac-6002) Seedlings under Abiotic Stress (Pendimethalin) Condition. Asian Journal of Crop Science, 4: 80-85.
DOI: 10.3923/ajcs.2012.80.85
URL: https://scialert.net/abstract/?doi=ajcs.2012.80.85
DOI: 10.3923/ajcs.2012.80.85
URL: https://scialert.net/abstract/?doi=ajcs.2012.80.85
INTRODUCTION
Agricultural research till now has been primarily concerned how yield can be increased by the use of fertilizers, pesticides, irrigation etc. Pesticide form an essential part of the crop production technology that makes it possible for the farmers to feed the ever growing population. Crops differ in their responses to various biotic and abiotic stresses. Among the abiotic stresses, chemical stress is known to affect all the metabolic processes resulting in reduced crop growth and yield. The herbicide pendimethalin N-(1-ethyl proply)-2,6-dinitro 3,4 xylidine belongs to di-nitroaniline group is a highly volatizing chemical and it may cause injury or damage to the non-target plant species in addition to the weeds (Ashok et al., 1995; Madhu et al., 1996; Pahwa et al., 1988). Ashok et al. (1995) have reported in their findings that pendimethalin residue reduced the root and shoot dry weight of sorghum after 200 days of application. Madhu et al. (1996) observed adverse effect of pendimethalin on dry weight of succeeding crop viz., maize, soybean and cucumber in puddle seeded rice. The effect of herbicides on the growth and metabolism of crop plants whether beneficial or detrimental is the subject of interest to the scientist since long ago. Hence, the present investigation aims at a detail evaluation on the biochemical changes during germination and early growth of maize cv. NAC-6002, under varied concentration of pendimethalin.
MATERIALS AND METHODS
Maize (Zea mays L.) NAC-6002 variety was obtained from the University of Agricultural Science, Nagenahalli, Mysore. The herbicide Pendimethalin (TATA PANIDA) was obtained from The Rallies India Ltd., Peenya, Bangalore. The present study was taken up with commercial grade of (pendimethalin) TATA PANIDA at 0.5, 1.0, 2.5, 5.0 and 10.0 ppm along with control using maize cv. NAC-6002. Hundred healthy grains of NAC-6002 variety of maize were surface sterilized with 2% sodium hypochloride for 5 min and rinsed with distilled water for 10-12 times to remove excess chloride. Ten grains (grain sample of 100 seeds for each replicate) were placed on a 9 cms petriplates containing 8 mL of Hoagland’s nutrient media (Hoagland and Arnon, 1938) supplemented with a range of concentrations namely 0.5, 1.0, 2.5, 5.0 and 10.0 ppm of herbicide Pendimethalin along with control in dark at 28±2°C for four days. Later, on 5th day onwards, the germinated seedlings were exposed to 12 h light intensity and the seedlings were further grown up to 15-days.
The data of seed germination, length of radicle and plumule were recorded according to ISTA standard (Anonymous, 1985) and the biochemical assay were conducted separately for shoot-root axis and cotyledons from the 4, 6, 8, 10 and 15 days old seedlings. According to the standard protocol prescribed, the estimation of protein (Lowry et al., 1951), total carbohydrate (DuBois et al., 1956), reducing sugar (Miller, 1959) and total starch (Clegg, 1956) were calculated and the data obtained were subjected to statistical analysis, using SPSS package version-10.2 with Tukey’s significant test at 5%.
RESULTS AND DISCUSSION
Seed germination and associated biochemical changes decreased significantly, during germination and early growth of treated sets when compared to control. Seed germination in the control was maximum, i.e., 95.31%. The germination percentage decreased drastically in the treated sets with increasing concentrations of pendimethalin. A severe decrease of about 69% was observed at high concentration of pendimentalin-10.0 ppm when compared to control. Similarly, the length of plumule decreased by 77% and the length of radicle decreased upto 90% at high concentration of herbicide pendimethalin (10.0 ppm) (Table 1). The present investigation has shown a significant decline in seed germination and its subsequent growth by herbicide-pendimethalin indicating that some of the biochemical processes taking place during germination and early growth are impaired.
| Table 1: | Effects of pendimethalin on percent germination, length of radicle and plumule |
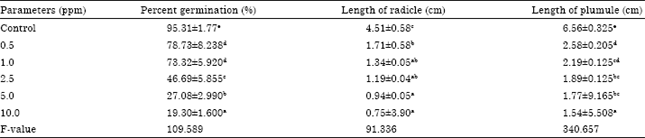 | |
| Note: Mean±SD followed by same letter are not statistically significant between the concentration, when subjected to SPSS package ver. 10.2, according to Tukey’s mean range test | |
The impaired metabolism consequently leads to reduced growth. Several reports have demonstrated the adverse effects of herbicide on seed germination. Delvin and Cunningham (1970), Dhillonand Anderson (1972), Schultz et al. (1967), Nehru et al. (1999) and Delvin and Cunningham (1970) have reported the adverse effects of alachlor and propachlor herbicides which severely reduced the germination of barley by interfering with the metabolic activities related to it. Similarly Schultz et al. (1967) and Nehru et al. (1999) observed the adverse effect of herbicide pendimethalin and trifluralin on seed germination and early growth in corn, mungbean and maize, respectively. Our results also indicate that higher concentrations of herbicide-pendimethalin inhibited the germination of maize grains at varying degree. This may be attributed to the adverse effect of the herbicide on degradation and mobilization of seed reserves.
The external morphology of development is often marked by biochemical changes of seed reserves and enzymes of the internal tissues and is considered as markers of growth and development. During seed germination and seedling growth, the seed reserve gets hydrolyzed and a change in the cellular and organellar constituents such as proteins, lipids and carbohydrates takes place. However, the rate of change varies from crop to crop and species to species (Ching, 1972; Ashton, 1976; Osborne, 1962).
Protein is a major storage reserve in many plant seeds. This protein is hydrolyzed during seed germination by proteolytic enzymes. Its metabolism has been implicated in the breaking of seed dormancy as seed storage proteins plays a pivotal role providing reducing nitrogen, carbon and amino acids to growing tissues (Staswick, 1988, 1990). In cotyledons, there was 65% decrease in total protein content from 4 to 15 days in control and it was only 25-35% in different treatments. On other hand, the total protein content (55%) increased from 4 to 15 days in both control and 0.5 ppm treatment, while it varied between 32-37% among other treatments of shoot-root axis (Table 2). The slow degradation of protein in treated cotyledons over control may be attributed to the inhibition of proteolytic activity of these enzymes (Mann et al., 1965; Moreland et al., 1969), whereas significant decrease of protein content in treated shoot-root axis over control indicates that it may be due to the inhibition of dipeptidase activity (Tsay and Ashton, 1971). The rate of protein synthesis and degradation in treated seedlings showed slowing down in the rate of protein synthesis and increase in the rate of protein break down which may be responsible for the observed rapid protein loss. These results are in line with Lorenzo et al. (1985) which demonstrates that pendimethalin might interfere with protein metabolism during germination and its early growth.
Carbohydrates are important components of storage and structural material in plants. They are the energy rich sources and are essential for germination of seeds. Germinating seeds are dependent on starch for their energy supply which is degraded into soluble forms by a-amylase enzyme. During germination the carbohydrate reserve of seed i.e., principally the starch gets degraded to provide sugars either via amylolytic or phosphorolytic pathways. They exist as free sugar and polysaccharides (Hedge and Hofreiter, 1962; Swain and Dekker, 1966). In cotyledons of soybean the reduction in total carbohydrate content from 4 to 15 days was 52% in control and 0.5 ppm treatment, while it varied between 32-46% in other treatments. On the other hand, higher content of total carbohydrate was recorded (61-67%) in the embryonic axis of the treated seedlings, when compared to control (42%) from 4 to 15 days. Similarly, the decrease in reducing sugar with time interval was 90% in control and varied between 66-80% in different treatments in cotyledons, while in shoot-root axis the increase in the reducing sugar from 4 to 15 days was 44% in control and it ranges between 54 to 66% among treatments.
| Table 2: | Effects of pendimethalin on total protien, carbohydrate, starch and reducing sugar content in Maize |
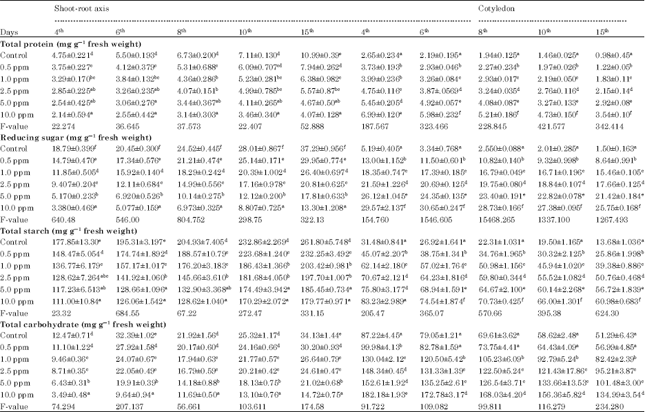 | |
| Mean±SD followed by same letter are not statistically significant between the concentration, when subjected to SPSS package ver. 10.2, according to Tukey’s mean range test | |
On the other hand, the decline in total starch content with time interval was 62% in control and it was varied from 33 to 43% in different treatments in cotyledons of soybean, while in shoot-root axis the total starch content with time interval was 37% increase in control and in different treatments it ranged from 41 to 47% (Table 2).
Penner (1968) reported that the reduction in degradation of total carbohydrate and starch content in barley seedlings by herbicides bromoxylin and amiben. He also reported that these herbicides interfere in the processes of hydrolytic enzymes synthesis, which are involved in the degradation of food reserves. And more pronounced inhibition of amylase activity by several herbicides has been reported by Jones and Foy (1971) which impair the degradation and mobilization of seed serves. Our results are in accordance with the above findings in case of total carbohydrate and starch content which showed reduced content in the shoot-root axis and a higher content in the cotyledon of all treatment at five different time interval over control. Felipe et al. (1988) reported that decline in RuDP carboxylase activity results in reduction in reducing sugar content in wheat after isoproturon treatment. Similarly, the decline in reducing sugar in shoot-root axis and a higher content in the cotyledons in all treatments and at different time intervals in our study may also be due to lower activity of RuDP carboxylase enzyme.
In short, pendimethalin bears adverse effects on the seed germination and early growth of developing maize seedlings. Each concentration affected the parameters studied at different point of observation. The herbicidal action of this degradation of reserve materials and on the control of these processes during germination may be one of the factors responsible for greater susceptibility of seedlings to the herbicide pendimethalin than that observed in mature plant. However, further studies are needed to strengthen this conclusion.
REFERENCES
- Ashton, F.M., 1976. Mobilization of storage proteins in seeds. Ann. Rev. Plant Physiol., 27: 95-117.
Direct Link - Clegg, K.M., 1956. The application of the anthrone reagent to the estimation of starch in cereals. J. Sci. Food Agric., 7: 40-44.
CrossRef - Delvin, R.M. and R.P. Cunningham, 1970. The inhibition of gibberellic acid induction of α-amylase activity in barley endosperm by certain herbicides. Weed Res., 10: 316-320.
CrossRef - Dhillon, N.S. and J.L. Anderson, 1972. Morphological, anatomical and biochemical effects of propachlor on seedling growth. Weed Res., 12: 182-189.
CrossRef - DuBois, M., K.A. Gilles, J.K. Hamilton, P.A. Rebers and F. Smith, 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem., 28: 350-356.
CrossRefDirect Link - Hoagland, D.R. and D.I. Arnon, 1938. The water culture method for growing plants without soil. Calif. Agric. Exp. Station, C347: 1-39.
Direct Link - Jones, D.W. and C.I. Foy, 1971. Herbicidal inhibition of GA-induced synthesis of α-amylase. Weed Sci., 19: 595-597.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Madhu, M., H.V. Nanjappa and H.R. Shivakumar, 1996. Residual effect of herbicides on succeeding crops in puddle seeded rice. Crop Res., 12: 241-246.
Direct Link - Mann, J.D., L.S. Jordan and B.E. Day, 1965. A survey of herbicide for their effect on protein synthesis. Plant Physiol., 40: 840-843.
PubMed - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - Moreland, D.E., S.S. Malhotra, R.D. Gruenhagen and E.H. Shokrah, 1969. Effects of herbicides on RNA and protein synthesis. Weed Sci., 17: 556-563.
Direct Link - Penner, D., 1968. . Herbicidal influence on amylase in barley and squash seedlings. Weed Sci., 16: 519-522.
Direct Link - Staswick, P.E., 1988. Soybean vegetative storage protein structure and gene expression. Plant Physiol., 87: 250-254.
Direct Link - Staswick, P.E., 1990. Novel regulation of vegetative storage protein genes. Plant Cell, 2: 1-6.
Direct Link - Swain, R.R. and E.E. Dekker, 1966. Seed germination studies. II Pathways for starch degradation in germinating pea seedlings. Biochem. Biophys. Acta, 122: 87-100.
Direct Link - Tsay, R. and F.M. Ashton, 1971. Effect of several herbicides on dipeptidase activity of squash cotyledons. Weed Sci., 19: 682-684.
Direct Link
THIRUGNANAM Reply
NAC-6002