
H. Ghani Zadeh
Department of Weed Identification and Control, Faculty of Agriculture, Azad Islamic University of Shushtar, Iran
S. Lorzadeh
Department of Weed Identification and Control, Faculty of Agriculture, Azad Islamic University of Shushtar, Iran
N. Aryannia
Department of Weed Identification and Control, Faculty of Agriculture, Azad Islamic University of Shushtar, Iran
Asian Journal of Crop Science
Year: 2011 | Volume: 3 | Issue: 4 | Page No.: 179-187







ABSTRACT
The knowledge of differences between weed species in their relative capacity to capture growth-limiting resources provides information which can be used to predict competitive ability of weed. A field study was conducted to compare weeds growth indices to evaluate weeds competitive ability. The experimental design was a Randomized Complete Block (RCBD) with 3 replications. Treatments were consisted of increasing durations of weed which were established by delaying weed Removal Time (RT) to different corn growth stages. Growth analysis was performed by determining weeds growth indices. In this trial different weed species exhibited marked differences in growth indices during growing season. Compare to other weeds, Barnyard grass had the greatest leaf area index and weed growth rate from the beginning to the middle of growing season. However, redroot pigweed achieved its maximum growth later from the middle of growing season to the end of season and became the dominant weed for that time. Redroot pigweed had by far the greatest relative growth rate compare with other weeds. It reveals that barnyard grass was the dominant weed for the beginning of growing season. However, from the middle of growing season till the end redroot pigweed became the dominant weed.
PDF Abstract XML References Citation
Received: June 27, 2011;
Accepted: July 15, 2011;
Published: September 08, 2011
How to cite this article
H. Ghani Zadeh, S. Lorzadeh and N. Aryannia, 2011. Evaluating Weeds Competitive Ability in a Corn Field in Southern West of Iran. Asian Journal of Crop Science, 3: 179-187.
DOI: 10.3923/ajcs.2011.179.187
URL: https://scialert.net/abstract/?doi=ajcs.2011.179.187
DOI: 10.3923/ajcs.2011.179.187
URL: https://scialert.net/abstract/?doi=ajcs.2011.179.187
INTRODUCTION
Weeds are one of the most important factors that influence the agricultural production systems (Keramati et al., 2008; Oad et al., 2007; Yaghoobi and Siyami, 2008; Aghaalikhani and Yaghoobi, 2008). They reduce crops quantity and increase the cost of production. Using herbicides as an effective tool to control weeds causes environmental issues and increase the number of herbicide resistant weeds (Ghanizadeh et al., 2010). Considering the above mentioned factors, the Integrated Weed Management (IWM) was recommended to optimize herbicide usage in weed management (Knezevic et al., 2002). In order to manage weeds according to IWM, it is necessary to evaluate weeds competition mechanisms by investigating weeds growth Indices. These studies provide fruitful information of how weeds influence crops yields and it helps plant breeders to develop weed competitive varieties (Baghestani et al., 2006). Weeds may affect crops growth by different competitive mechanism. Leaf Area Index (LAI) is an important factor showing weeds competitiveness (Knezevic et al., 1997). It is noted that light capturing is an important factor in the competitiveness mechanism of weeds (Traore et al., 2003). It is reported that plants with high leaf area are more competitive as a result of being able to capture higher Photosynthetic Active Radiation (PAR) (Crotser and Witt, 2000). Weeds also can reduce the availability of PAR to the lower layer leaves of the crop canopy (Lindquist, 2001) and decrease the longevity and expansion rate of lower leaves due to reduce available soil nitrogen and moisture levels (Wolfe et al., 1988). Plant growth rate is also an important factor in competitive relationships (Mohammadi, 2007). Other researchers stated that plants with higher growth rate are more competitive (Bastiaans et al., 1997; Ni et al., 2000; Baghestani et al., 2006).
Another parameter in plant competitive ability is Relative Growth Rate (RGR) which describes plant performance under different conditions (Karadavut et al., 2010). RGR is varied according to growth of plants (Karadavut et al., 2009). The RGR is a fundamental measure of dry matter production and a higher RGR is an indicator of more efficient resource acquisition by the plant (Mohammadi, 2007). The knowledge of differences between weed species in their relative capacity to capture growth-limiting resources provides information which can be used to predict competitive ability of weed (Roush and Radosevich, 1985) and to design methods of predicting crop yield loss associated with weeds (Moechnig et al., 2003). Therefore, the objective of this trial was to compare weeds growth indices to evaluate weeds competitive ability. In this trial a natural population of weed was used because weed species are varied in their competitive strategies. Besides, the emergence pattern of different weeds in field condition is different from those which have been investigated in controlled conditions.
MATERIALS AND METHODS
Location: A field experiment was conducted in 2008 at the Shushtar University experiment Station in the southern west of Iran (32°3' North latitude, 48°50' East longitude, 67 m). The soil type was clay loam with 0.2% organic matter and pH 7 (Ghanizadeh, 2008).
Cultural practices: Soil preparation consisted of primary and secondary tillage as well as cultural practices was conducted according to local practices for corn production. Corn seeds, cultivar SC704, sown at a spacing of 20 cm within the row spaced 75 cm apart at a population density of 66,600 plants ha-1. Plot size was five rows each 10 m long and plots were separated by two border rows. The middle three rows of each plot were used for data collection. Fertilizers were applied at the rate of 112 kg P ha-1 at sowing and 205 kg N ha-1 with 50% applied at the time of sowing and 50% at the time of 7-8 leaf-stage of corn.
Treatments and experimental design: The experimental design was a Randomized Complete Block (RCBD) with 3 replications. Treatments were consisted of increasing durations of weed which were established by delaying weed Removal Time (RT) to corn 3 leaf stage (V3), 6 leaf stage (V6), 9 leaf stage (V9), 13 leaf stage (V13) and tasseling (VT) based on average crop growth stage as described by Ritchie et al. (1997). Natural weed populations were used in trial. The progression of crop development was monitored by recording the average growth stage of 10 consecutive corn plants every 5th day.
Sampling: The first sampling of weeds was carried out at V3 (10 days after planting (DAP)). The consecutive samplings were carried out at each removal time. Weeds were harvested from three 0.5 m2 quadrates staggered on each side of the three middles corn rows within each experimental. At each harvest, weeds were excavated from the soil surface, divided into leaf- and stem part. Leaf area of the whole plant was measured with a leaf area meter (LI-3100A, LiCor Inc., Lincoln, Nebraska, USA). Harvested materials were oven dried at 75°C for 48 h and weighed.
Growth analysis: Growth analysis was performed by determining Leaf Area Index (LAI), Relative Growth Rate (RGR) and Weed Growth Rate (WGR) according to methods outlined by Hunt (1990). The values of plants leaf area and biomass were loge-transformed to maintain homogeneity of variance among the sampling data. The Total Dry Weight (TDW) and LAI of plants were fitted to Eq. 1 (Traore et al., 2003):
| (1) |
where, ln (y) is the loge-transformed the total biomass or leaf area per m2 and t is the Days After Emergence (DAE). The coefficients a, b and c are constants. The plants RGR were fitted with Eq. 2 (Kapur and Govil, 2004):
| (2) |
where, y is the relative growth rate and the coefficients b and c are constants. WGR was calculated by multiplying TDM by RGR (Eq. 3). The curves were fitted with EXCEL software:
| (3) |
Statistical analysis: Parameters of equation were analysed using PROC NLIN in SAS. Software (SAS, 1999).
RESULTS
Weeds community: During this experiment 5 weed species were observed. Barnyard grass (Echinochloa crus-galli), Redroot Pigweed (Amaranthus retroflexus) Common Purslane (Portulaca oleracea), Common Lambsquarters (Chenopodium album L.) and Yellow Nutsedge (Cyperus esculentus) were observed. According to Table 1 there was a sharp increase in the density of weeds until corn 6-leaf stage. However, the density of weeds decreased slightly after that time. Barnyard grass, redroot pigweed and common purslane were the most common weeds in this trial and presented higher weed density. Therefore, only the growth indices of these weeds were evaluated as they were dominant weeds in our trial.
Weeds total dry weight: In this trial different weed species exhibited marked differences in plant biomass accumulation (Fig. 1). During early growth (4 to 25 DAE), total plant biomass was greatest for barnyard grass and common purslane respectively. From 25 to approximately 50 DAE; there was a gradual shift in weeds total biomass accumulation. Total plant biomass was still greatest for barnyard grass but redroot pigweed was the second greatest weed in plant biomass accumulation. Both species exhibited greater plant biomass than common purslane. Total plant biomass peaked between 46 and 55 DAE for all species. At 65 DAE, the biomass of barnyard grass, redroot pigweed and common purslane was 720.5, 639.1 and 224.7 g m-1, respectively.
| Table 1: | The population density (plants/m2) of weed species in the experimental area |
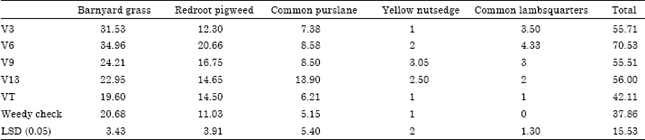 | |
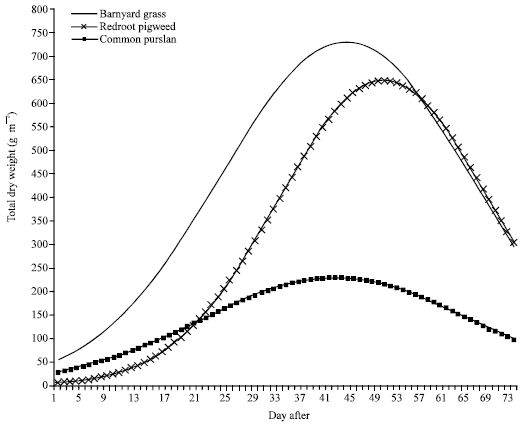 | |
| Fig. 1: | Weeds total dry biomass during growing season |
Weeds leaf area index: Leaf area index has an important role in competition on light. According to Fig. 2, the leaf area index of barnyard grass was greater than other weeds as a result of higher germination at the beginning in compare to other species in our trial. Redroot pigweed was the second greatest in the LAI. The LAI of common purslane was only higher than redroot pigweed at the beginning of growing season. However, this weed failed to maintain this priority till the end of season because of the competitive ability of this weed was not as much as other weeds like redroot pigweed or barnyard grass.
Weeds growth rate: Weed Growth Rate (WGR) is one of the factors to compare competitive ability among several weed species. According to Fig. 3, Barnyard grass had by far the greatest WGR due to the highest LAI among other weeds. The point is that the time to reach a peak of WGR among weed species was different. The maximum growth rate achieved 28, 43 and 60 DAE by Common purslane, barnyard grass and redroot pigweed, respectively.
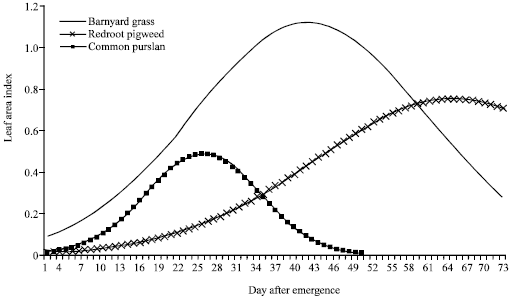 | |
| Fig. 2: | Weeds leaf area index during growing season |
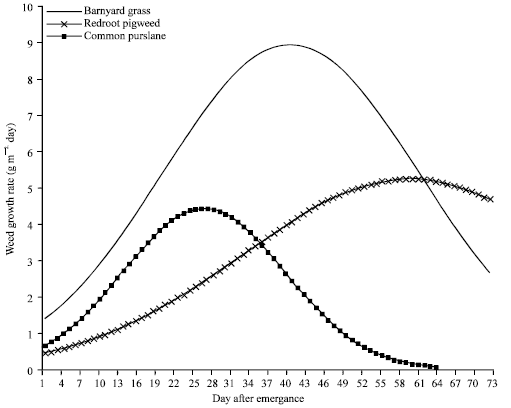 | |
| Fig. 3: | Weeds growth rate during growing season |
Weeds relative growth rate: For each species, RGR was greatest shortly after seedling emergence and declined throughout plant growth (Fig. 4). Common purslane exhibited lower RGR than either barnyard grass or redroot pigweed throughout much of its growth. RGR for redroot pigweed were greater than those for barnyard grass from 1 to 40 DAE.
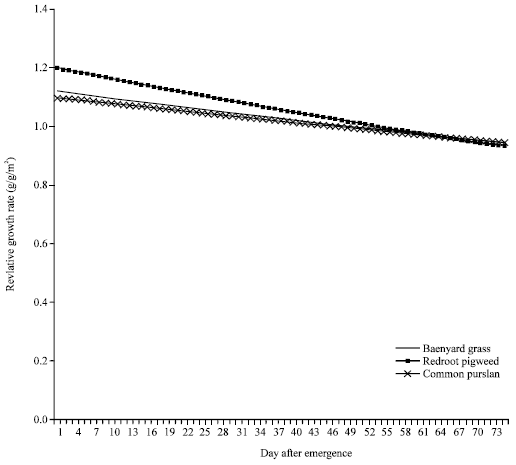 | |
| Fig. 4: | Weeds relative growth rate during growing season |
DISCUSSION
In this trial weed density of each species almost reach a peak at corn 6-leaf stage. However, weed density dropped significantly after that time due to inter- and intra-specific competition between plants community which led to remove the weak plants as a result of the self-thinning (Radosevich and Holt, 1984). The result of this experiment is in agreement with Yakubu et al. (2006). Total dry matter of each weed species showed different pattern during growing season. At the beginning the total biomass of common purslane was higher than redroot pigweed as redroot pigweed had a time lag of the germination at the beginning which caused a flashback in maximizing the density. It might be due to different level in the dormancy of redroot pigweed seed (Jha, 2008). However, at the end of growing season total biomass of redroot pigweed was higher than common purslane. This might be because of differences in weed competition mechanism and ability. For instance, it is stated that common purslane is less competitive in compare to barnyard grass or redroot pigweed (Miyanishi and Cavers, 1980). Therefore, common purslane would establish immediately after tillage and reach the flowering stage before experiencing any negative effects from more competitive weeds. It can be the mechanism of survival in the field for this species. At the end of growing season, the total biomass of all species reduced gradually. It can be explained by increasing the number of senesced leaves and decreasing the number of expanded leaves due to intra- and inter-specific competition (Sheibany et al., 2009). The leaf area index is an essential plant feature that responds to competition (Weiner et al., 2001; Baghestani et al., 2006). In this trial the time and percentage of germination showed positive correlation with Leaf area index. Compare to other weeds, barnyard grass showed by far the greatest LAI weeds due to early germination and higher density. The time lag of germination in redroot pigweed caused a delay in increasing in LAI in the beginning of growing season. However, redroot pigweed is able to shift its leaf area vertically to avoid shading effect from other plants (Rajcan and Swanton, 2001). It means reduction in light availability is associated with increases in partitioning of dry weight to stem tissue and stem elongation. It is noted that leaf area and dry matter were increasingly allocated to upper portions of the redroot pigweed under shading stress (McLachlan et al., 1993). For that reason, redroot pigweed exhibited higher LAI at the end of growing season compare to other weeds. WGR followed the same pattern that LAI did. Plant growth rate is a function of LAI (Vijayakumar et al., 2006). In fact, Whole plant productivity depends on LAI and resources allocation (Adelusi et al., 2006). For that reason, any factors which had negative effects on assimilation of leaves would influence plant growth rate negatively. For instance, it is noted that barnyard grass plants are too sensitive to shading (Maun and Barrett, 1986). For that reason, canopy closure of high plants like corn would increase the shading effects over barnyard grass and reduces the availability of Photosynthetic Active Radiation (PAR) to the lower layer leaves of the canopy. Therefore, reduction in longevity and expansion rate of leaves would reduce LAI and eventually, decrease the WGR of this weed. The data indicate that while barnyard grass exhibited lower biomass accumulation during early growth, its total dry matter production was greater due to its higher density. RGR is one of the most important determinants of competitive outcome (Wang et al., 2006). The results of this experiment indicated that redroot pigweed was more competitive than Barnyard grass and although there was a time lag of germination in this weed but redroot pigweed was able to compensate its loss by light-competition strategies. Cowan et al. (1998) stated that redroot pigweed was more competitive than barnyard grass.
CONCLUSION
According to the results, at the beginning of the growing season Barnyard grass was the dominant weed due to higher LAI and CGR at that time. However, canopy closure of corn limited competitive ability of that weed. On the other hand, redroot pigweed was able to upward shift in leaf area distribution within the canopy to avoid shading effects of corn and became dominant from the middle of growing season to the end of growing season.
REFERENCES
- Adelusi, A.A., O.A. Akamo and A.M. Makinde, 2006. Nitrogen fertilizer and weed (Euphorbia heterophylla Linn.) effects on the growth parameters of Macrotyloma geocarpa (Harms) marechal and beudet. Int. J. Bot., 2: 74-82.
CrossRefDirect Link - Aghaalikhani, M. and S.R. Yaghoobi, 2008. Critical period of weed control in winter canola (Brassica napus L.) in a Semi-Arid region. Pak. J. Biol. Sci., 11: 773-777.
CrossRefPubMedDirect Link - Baghestani, M.A., E. Zand and S. Soufizadeh, 2006. Iranian winter wheat's (Triticum aestivum L.) interference with weeds. II. Growth analysis. Pak. J. Weed Sci. Res., 12: 131-144.
Direct Link - Bastiaans, L., M.J. Kropff, N. Kempuchetty, A. Rajan and T.R. Migo, 1997. Can simulation models help design rice cultivars that are more competitive against weeds. Field Crops Res., 51: 101-111.
Direct Link - Cowan, P., S.E. Weaver and C.J. Swanton, 1998. Interference between pigweed (Amaranthus spp.), barnyardgrass (Echinochloa crus-galli) and soybean (Glycine max). Weed Sci., 46: 533-539.
Direct Link - Crotser, M.P. and W.W. Witt, 2000. Effect of Glycine max canopy characteristics, G. max interference and weed-free period on Solanum ptycanthum growth. Weed Sci., 48: 20-26.
CrossRef - Ghanizadeh, H., S. Lorzadeh and N. Ariannia, 2010. Critical period for weed control in corn in the South-West of Iran. Asian J. Agric. Res., 4: 80-86.
CrossRefDirect Link - Karadavut, U., K. Kokten and Z. Kavurmaci, 2010. Comparison of relative growth rates in silage corn cultivars. Asian J. Anim. Vet. Adv., 5: 223-228.
CrossRefDirect Link - Knezevic, S.Z., M.J. Horak and R.L. Vanderlip, 1997. Relative time of redroot pigweed (Amaranthus retroflexus L.) emergence is critical in pigweed-sorghum (Sorghum bicolor L. Moench) competition. Weed Sci., 45: 502-508.
Direct Link - Knezevic, S.Z., S.P. Evans, E.E. Blankenship, R.C. van Acker and J.L. Lindquist, 2002. Critical period for weed control: The concept and data analysis. Weed Sci., 50: 773-786.
CrossRefDirect Link - Keramati, S., H. Pirdashti, M.A. Esmaili, A. Abbasian and M. Habibi, 2008. The critical period of weed control in soybean (Glycine max (L.) Merr.) in North of Iran conditions. Pak. J. Biol. Sci., 11: 463-467.
CrossRefPubMedDirect Link - Maun, M.A. and S.C.H. Barrett, 1986. The biology of Canadian weeds 77. Echinochloa crus-galli L. Beauv. Can. J. Plant. Sci., 66: 739-759.
Direct Link - McLachlan, S.M., M. Tollenaar, C.J. Swanton and S.F. Weise, 1993. Effect of corn-induced shading on dry matter accumulation, distribution and architecture of redroot pigweed (Amaranthus retroflexus). Weed Sci., 41: 568-573.
Direct Link - Miyanishi, K. and P.B. Cavers, 1980. The biology of Canadian weeds: 40. Portulaca oleracea L. Can. J. Plant Sci., 60: 953-963.
CrossRefDirect Link - Moechnig, M.J., C.M. Boerdoom, D.E. Stoltenberg and L.K. Binning, 2003. Growth interaction in communities of common lambsquarters (Chenopodium album), Giant foxtail (Setaria faberi) and corn. Weed Sci., 51: 363-370.
CrossRef - Mohammadi, G.R., 2007. Growth parameters enhancing the competitive ability of corn (Zea mays L.) against weeds. Weed Biol. Manag., 7: 232-236.
CrossRef - Ni, H., K. Moody, R.P. Robles, E.C. Paller Jr. and J.S. Lales, 2000. Oryza sativa plant traits conferring competitive ability against weeds. Weed Sci., 48: 200-204.
CrossRef - Oad, F.C., M.H. Siddiqui and U.A. Buriro, 2007. Growth and yield losses in wheat due to different weed densities. Asian J. Plant Sci., 6: 173-176.
CrossRefDirect Link - Rajcan, I. and C.J. Swanton, 2001. Understanding maize-weed competition: Resource competition, light quality and the whole plant. Field Crop Res., 71: 139-150.
CrossRefDirect Link - Roush, M.L. and S.R. Radosevich, 1985. Relationships between growth and competitiveness of four annual weeds. J. Applied Ecol., 22: 895-905.
Direct Link - Sheibany, K., M.A.B. Meybodi and A. Atri, 2009. Competitive effects of redroot pigweed (Amaranthus retroflexus) on growth indices and yield of corn. Weed Biol. Manag., 9: 152-159.
CrossRef - Traore, S., S.C. Mason, A.R. Martin, D.A. Mortensen and J.J. Spotanski, 2003. Velvetleaf interference effects on yield and growth of grain sorghum. Agron. J., 95: 1602-1607.
Direct Link - Vijayakumar, M., S. Ramesh, N.K. Prabhakaran, P. Subbian and B. Chandrasekaran, 2006. Influence of system of rice intensification (sri) practices on growth characters, days to flowering, growth analysis and labour productivity of rice. Asian J. Plant Sci., 5: 984-989.
CrossRefDirect Link - Wang, G., M.E. McGiffen Jr. and J.D. Ehlers, 2006. Competition and growth of six cowpea (Vigna unguiculata) genotypes, sunflower (Helianthus annuus) and common purslane (Portulaca oleracea). Weed Sci., 54: 954-960.
CrossRef - Weiner, J., H.W. Griepentorg and L. Kristensen, 2001. Suppression of weed by spring wheat (Triticumaestivum) increases with crop density and spatial uniformity. J. Applied Ecol., 38: 784-790.
CrossRefDirect Link - Wolfe, D.A., D.W. Henderson, T.C. Hsiao and A. Alvino, 1988. Interactive water and nitrogen effects senescence of maize. II. Photosynthesis decline and longevity of individual leaves. Agron. J., 80: 865-870.
Direct Link - Yakubu, A.I., J. Alhassan, A. Lado and S. Sarkindiya, 2006. Comparative weed density studies in irrigated carrot (Daucus carota L.) potato (Solanum tuberosum L.) and wheat (Triticum aestivum L.) in Sokoto-Rima valley, Sokoto State, Nigeria. J. Plant Sci., 10: 14-21.
CrossRefDirect Link - Yaghoobi, S.R. and K. Siyami, 2008. Effect of different periodical weed interference on yield and yield component in winter canola (Brassica napus L.). Asian J. Plant Sci., 7: 413-416.
CrossRefDirect Link