
E. E. Mahdy
Department of Agronomy, Faculty of Agriculture, Assiut University, Assiut, Egypt
M. A. Ali
Department of Agronomy, Faculty of Agriculture, South Valley University, Qena, Egypt
A. M. Mahmoud
Department of Agronomy, Faculty of Agriculture, Assiut University, Assiut, Egypt
Asian Journal of Crop Science
Year: 2011 | Volume: 3 | Issue: 1 | Page No.: 1-15







ABSTRACT
This study was undertaken to estimate the general and specific combining ability of different female and male lines in F1 combinations for grain yield and some important traits and estimate heterosis as a criterion for developing superior hybrids. The twenty five hybrids and their parents were evaluated at eight environments; two planting dates at two locations in 2007 and 2008 seasons. The obtained data showed that highly significant differences among parents, F1 crosses and parents versus crosses over the two years and over all environments for traits. An evaluation of general combining ability variance components as estimated from male and/or female overall environments were larger than those of specific combining ability for days to 50% blooming, plant height and 1000-grain weight. However, opposite results were obtained for grain yield Mg ha-1. The estimates of general combining ability and specific combining ability variance components varied greatly from location to location and from early to late planting for days to 50% blooming, plant height and 1000-grain weight. Some parents having significant negative general combining ability for days to heading and significant positive for plant height, 1000-grain weight and grain yield were considered as good combiners. Significant positive heterosis in grain yield heterosis was found for more than half of the hybrids studied. Several cross combinations showed significant positive 1000-grain weight heterosis, significant negative days to heading heterosis and good performance. Since selection of grain sorghum hybrids in this study should be based on high grain yield, early maturing, taller plants and heavier grain weight. Information on general and specific combining ability and heterosis for those four traits could contribute to more efficient breeding program.
PDF Abstract XML References Citation
Received: October 11, 2010;
Accepted: December 03, 2010;
Published: March 25, 2011
How to cite this article
E. E. Mahdy, M. A. Ali and A. M. Mahmoud, 2011. The Effect of Environment on Combining Ability and Heterosis in Grain Sorghum (Sorghum bicolor L. Moench). Asian Journal of Crop Science, 3: 1-15.
DOI: 10.3923/ajcs.2011.1.15
URL: https://scialert.net/abstract/?doi=ajcs.2011.1.15
DOI: 10.3923/ajcs.2011.1.15
URL: https://scialert.net/abstract/?doi=ajcs.2011.1.15
INTRODUCTION
Grain sorghum (Sorghum bicolor L. Moench) is the fourth most important cereal crop behind wheat, rice and maize, and is grown throughout the arid and semi-arid tropics (Smith and Frederiksen, 2000). At present, grain sorghum is a minor component of livestock feeds. Local demands for cereals, including grain sorghum is progressively increasing due to population growth and total production is not sufficient to cover the internal demands (Ali, 2000). The discovery of cytoplasmic male-sterility in sorghum by Stephens and Holland (1954) and Doggett (1969) facilitates the commercial utilization of hybrid vigour. Hybrid cultivars make use of male sterility to enhance the combining abilities of the parental lines, resulting in heterosis and significant increases in phenotypic traits such as yield, plant height and days to flowering (Reddy et al., 2006). The development of hybrids in Egypt is still depending on the exotic cytoplasmic male-sterile and restorer lines from USA and ICRISAT. Such lines have to be evaluated for adaptation under several environments and the required agronomic practices before testing their combining ability and heterotic response. Based on tests of the general and specific combining abilities, good combiner lines which can contribute to hybrid vigour are identified.
In breeding programs, information on combining ability and heterosis of parents and crosses are very important. By analyzing combining ability and estimating degree of heterosis, clues about the nature of gene action, desirable parents and important yield traits will emerge, particularly in those crops which are amenable to commercial production of F1 hybrid seed using cytoplasmic male sterility; sorghum is one of such crops.
Several sorghum reports indicate that general (GCA) and specific (SCA) combining ability effects for some parental lines (male and female lines) and hybrids were positive and highly significant for grain yield (Hovny, 2000; Ali, 2000; Hovny et al., 2001; Hovny et al., 2005; Mahmoud, 2007; Essa, 2009). Can et al. (1997) pointed out that some parents were identified having high positive GCA for grain yield and negative for days to heading which were considered as good combiners. However, high positive heterosis in grain yield was found for more than half of the hybrids.
Estimates of average better parent heterosis for grain yield in grain sorghum ranged from 9.0 to 97.0%. Lower estimates were obtained with crosses of adapted parent lines (Amir, 1999; Abd-El-Mottaleb, 2004; Hovny et al., 2001; Essa, 2009), while high values were most often resulted from studies which involved exotic germplasm (Thawari et al., 2000; Abd-El-Halim, 2003; Mahmoud, 2007; Mohamed, 2007) or which were conducted under environmental stress conditions (Prabhakar, 2001; Al-Naggar et al., 2002; Hovny et al., 2005; Abo-Zaid, 2007; Abd-El-Mottaleb, 2009). Amir (1999), Ali (2000), Abd-El-Halim (2003), Abd-El-Mottaleb (2004), Hovny et al. (2005), Mahmoud (2007), Mohamed (2007) and Essa (2009) stated that the F1 hybrids showed range of heterosis with negative and positive values which indicated the potential for developing hybrids superior to their better parent for earliness, plant height, grain yield and 1000-grain weight. The objectives of this study were to (i) estimate the GCA and SCA of different female and lines in F1 combinations for grain yield and some important traits and (ii) estimate heterosis as a criterion for developing superior hybrids.
MATERIALS AND METHODS
Genetic materials: Twenty five top-cross grain sorghum hybrids were developed at Qena Agriculture Research Farm, South Valley University, in two successive summer seasons (2006 and 2007). These hybrids were produced from crossing five introduced cytoplasmic male sterile lines (A-lines) to five restorer lines (R-lines). The A-lines included A-73, A-93, A-604, A-613 and A-614 and the R-lines were R-210, R-272, R-273, R-295 and R-92010.
Field trials: The resulting 25 F1 hybrids and their 10 parents were tested in yield trials at two locations (Assiut Agric. Res. Farm, Assiut Univ. and Qena Agric. Res. Farm, South Valley Univ.). In each location, the genotypes were sown at early (15 th June) and late (15th July) planting dates in 2007 and 2008 summer seasons. The soil type at Assiut is clay (soil pH was 7.9, organic matter (%) was 1.61, total N (%) was 0.07, P (ppm) was 11.25, K (%) was 0.35 and calcium carbonate (%) was 2.5). While, the soil type at Qena is sandy loam (soil pH was 8.12, organic matter (%) was 0.35, total N (%) was 0.04, P (ppm) was 9.4, K (%) was 0.19 and calcium carbonate (%) was 13.6).
Genotypes were arranged in a randomized complete block design with three replications in each experiment. The experimental unit was single row of 6 m long, 60 cm apart and 20 cm between hills within a row. Seedling were thinned three weeks after planting to two plants per hill; 168700 plants ha-1. Data of grain yield were taken from the middle portion of each plot (3 m) and later converted to Mg ha-1 at 15% moisture. All culture practices were applied as recommended for grain sorghum production.
Statistical analysis: Combined analyses of variance were done for the data over the two years and over all environments according to Gomez and Gomez (1984) after carrying out homogeneity test. The genetic analysis was performed out using line x tester analysis according to Kempthorne (1957). Additionally, the procedures described by Singh and Chaudhry (1977) were used to estimate General Combining Ability (G.C.A.) effects for each female and male parents and Specific Combining Ability (S.C.A.) effects for hybrid combinations. Estimates of the variance components for general and specific combining ability and their interactions were computed according to Beil and Atkins (1967). Variance components were tested for significance according to Robinson et al. (1955) as follows:
where, Mi is the ith mean square in the formula, di is the degree of freedom associated with the ith mean square and C2 is the divisor of the function of mean squares.
The standard error of variance components was calculated as the square root of variance of the estimate. As the distribution of the variance components is unknown, the component of genetic variance was considered to be significant from zero if its value was more than twice of its standard error (Mode and Robinson, 1959). Heterosis percentage was determined by using the following equation:
where, MF1 and HP are means for the F1 hybrid and high parent, respectively. Test of significance were made by using LSD.
RESULTS AND DISCUSSION
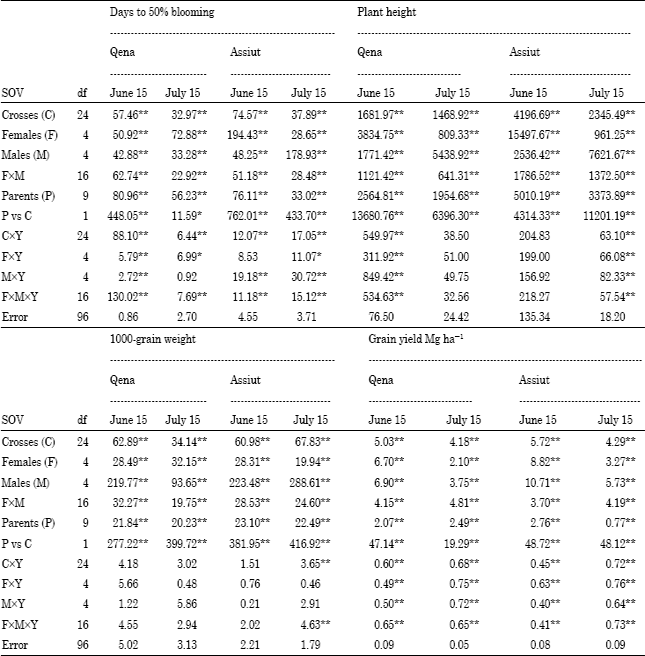
Table 1 and 2 presented the results of ANOVA for parents, F1 hybrids and their partitions; (males and females and their interaction). Significant differences (p<0.01) among parents, F1 hybrids and their partitions; (males and females and their interaction) were found for all traits, indicating wide genetic diversity among the genotypes. Mean square due to females was higher than the males for days to blooming under most environments. This indicates that the great differences among the females for this trait. While the mean square among males were larger than among females for grain weight and grain yield under early and late planting in both locations in the two years, indicating wide differences among the males.
| Table 1: | Combined analysis of variance for days to 50%blooming, plant height, 1000-grain weight and grain yield Mg ha-1 of 25 F1, s and their 10 parents over two years |
 | |
| *, ** Significant at 0.05 and 0.01 probability levels, respectively | |
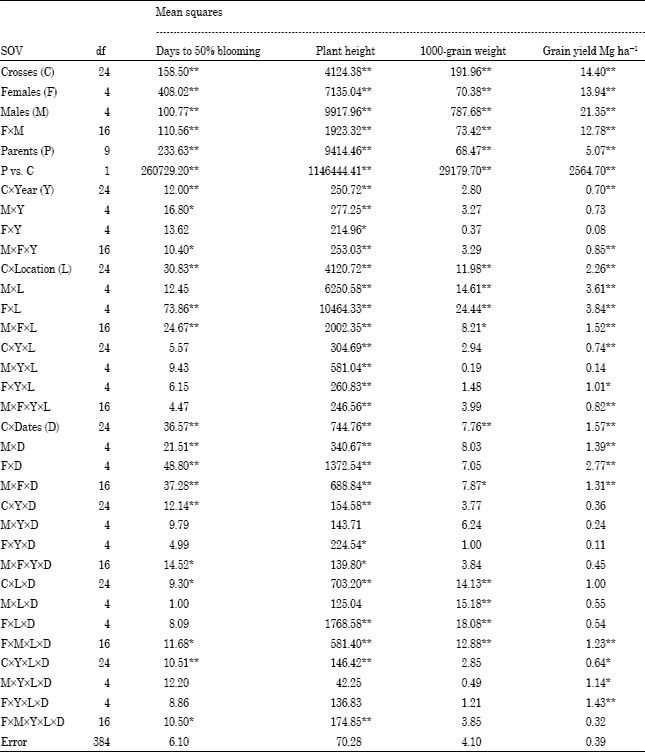
The mean square for crossesxYearxlocationsxdates was significant (p<0.01) for all traits except 1000-grain weight, emphasizing the need to evaluate such materials in several environments (Table 2). The parents vs. crosses mean squares was significant (p<0.01) for all traits, reflecting the average heterotic effect for these traits, their magnitudes were large compared with those for all sources of variation. Similar results were obtained by Amir (1999), Ali (2000), Biradar et al. (2000), Kenga et al. (2004), Bakheit et al. (2004), Hovny et al. (2005), Mohamed (2007), Mahmoud (2007), Hovny and El-Dsouky (2007) and Abd-El-Mottaleb (2009) and Essa (2009).
| Table 2: | Combined analysis of variance for days to 50%blooming, plant height, 1000-grain weight and grain yield Mg ha-1 of 25 F1, s and their 10 parents over all environments |
 | |
| *, ** Significant at 0.05 and 0.01 probability levels, respectively | |
Combining ability
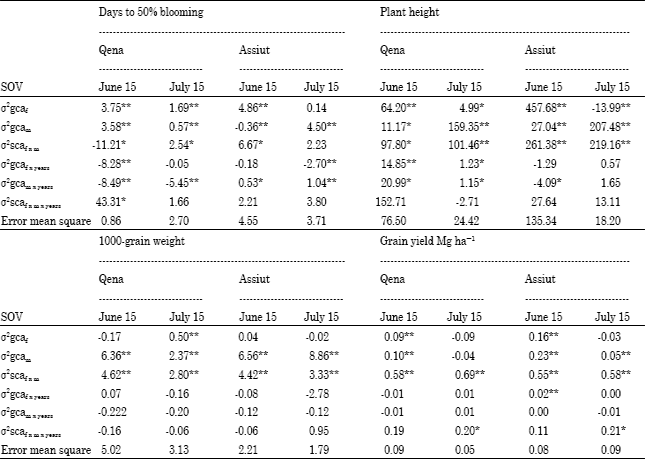
Combining ability variance: Table 3 and 4 showed the estimates of the variance components for general and specific combining ability for all traits. The results for days to blooming and 1000-grain weight, combined variance components for general effects were markedly larger than the component for specific effects (Table 3). For these traits, the interactions of general and specific effects with years gave very small and predominantly no significant variance components (Table 3), indicating that expression of these traits are controlled mostly by the additive effects of genes that are stable under early and late planting at the two locations over both years.
| Table 3: | Variance components for general and specific combining ability and their interactions with years under early and late planting at two locations |
 | |
| *, ** Significant at 0.05 and 0.01 probability levels, respectively | |
The data showed that variance components for males, females and femalesxmales were significantly positive for plant height under different environments, indicating that both additive and non-additive genetic variance were important for the inheritance of this trait. The data for grain yield indicate that the sum of the variance components for specific effects was markedly larger than the component for general effects (Table 3 and 4). This suggests that expression of this trait was controlled mostl y by the non-additive effects of genes that are stable in two dates at the two locations in both years and overall environments. Several workers have indicated that additive and non-additive components of genetic variance were important for these studied traits (Can et al., 1997; Amir, 1999; Biradar et al., 2000; Mostafa and El-Menshawi, 2001; Kenga et al., 2004; Hovny et al., 2005; Mohamed, 2007; Mahmoud, 2007; Abd-El-Mottaleb, 2009).
Combining ability effects
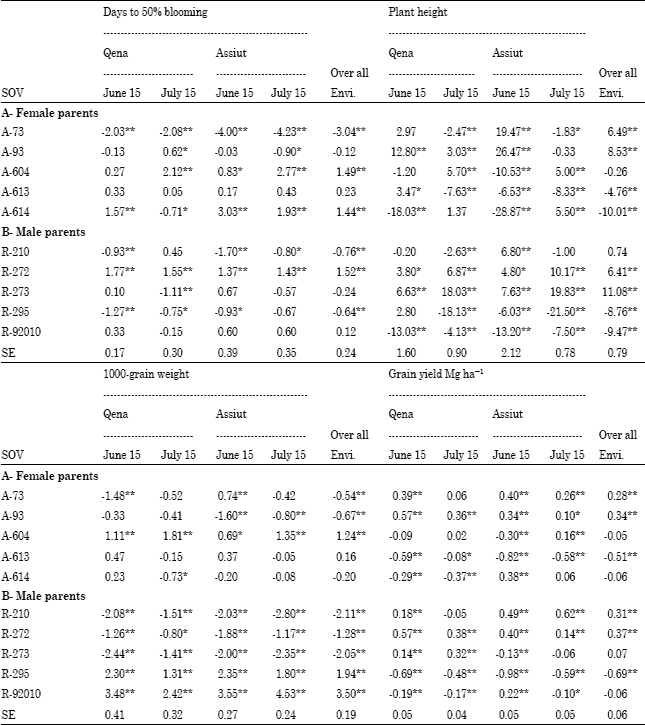
General combining ability (gca): The primary criteria for selection of desirable parents are usually based on mean values and additive gene action for traits under consideration. Genetically, gca effect is associated with additive gene action. Highly significant negative gca value for trait of days to flowering was obtained for the female line A-73 under early and late planting at two locations over the two years and overall environments, in which this female parent contributed to improving short duration to blooming in the crosses (Table 5). The male lines R-210 and R-295 showed also highly significant negative gca effect for days to flowering under most different planting dates at two locations over the two years and overall environments.
| Table 4: | Variance components for general and specific combining ability and their interactions overall environments |
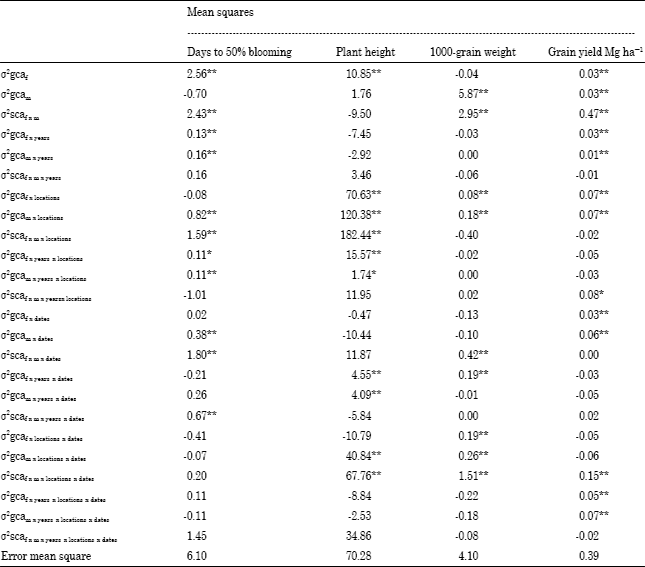 | |
| *, ** Significant at 0.05 and 0.01 probability levels, respectively | |
The male lines R-272 and R-273 showed a positive and highly significant gca effect for plant height under different planting dates at two locations over the two years and overall environments, this contributed to increase plant height in the crosses. For 1000-grain weight, the female parent A-604 and the male parents R-295 and R-92010 displayed highly significant positive gca effects under early and late planting in both locations over the two years and overall environments. These parents are considered to be good combiners for grain weight. Parental lines such as the female parent A-93 and the male parent R-272 had positive and highly significant gca effects for grain yield under early and late planting at Qena and Assiut over the two years and overall environments, indicating that they would be good parents of high grain yielding offspring. The parents selected for a particular trait were not always acceptable for other traits. For example, R-272 had high mean grain yield and highly significant positive gca effect but longer duration to flowering was not desirable in this study under different environments. Results from this study showed that the female line A-73 and the male line R-210 were considered to be good combiners for days to blooming and grain yield under different environments overall environments (Table 5). It could be expected that both parents could show better heterosis in F1 hybrids and produce earlier and high yielding genotypes in segregating generation. These results are in harmony with those obtained by Ali (2000), Biradar et al. (2000), Kenga et al. (2004), Bakheit et al. (2004), Hovny et al. (2005), Mohamed (2007) and Essa (2009).
| Table 5: | Estimates of general combining ability effects of parents (g.c.a.) for days to 50% blooming, plant height, 1000-grain weight and grain yield Mg ha-1 at eight and over all environments |
 | |
| *, ** Significant at 0.05 and 0.01 probability levels, respectively | |
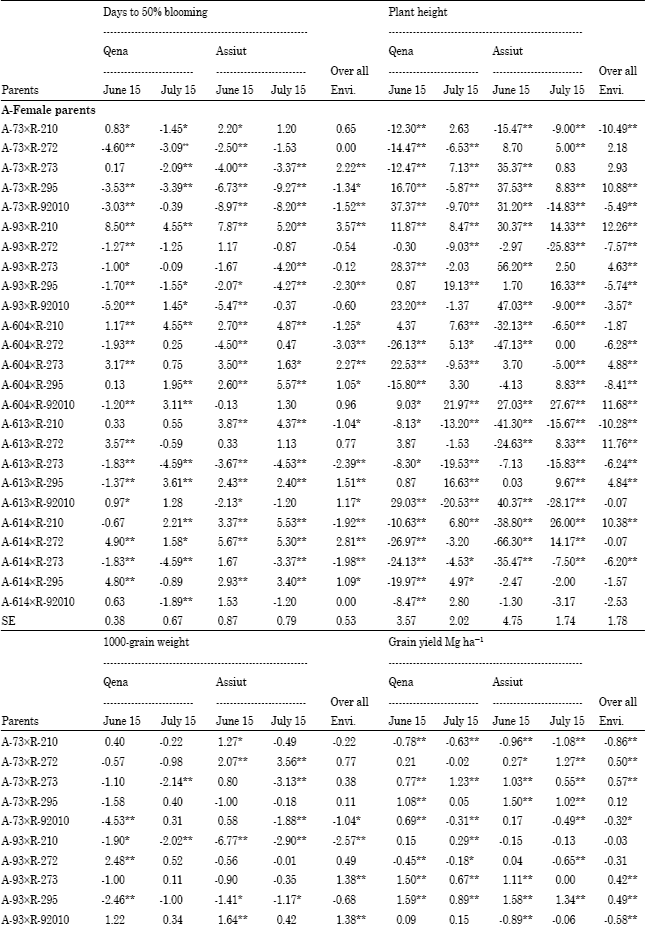
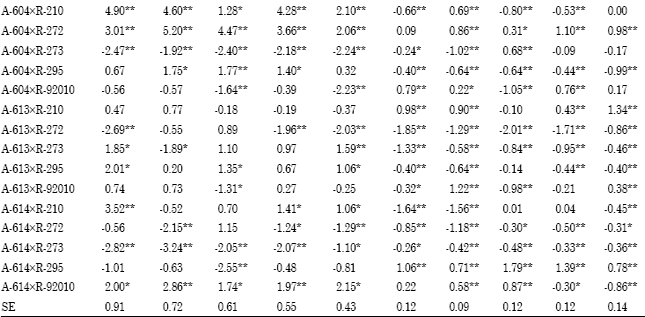
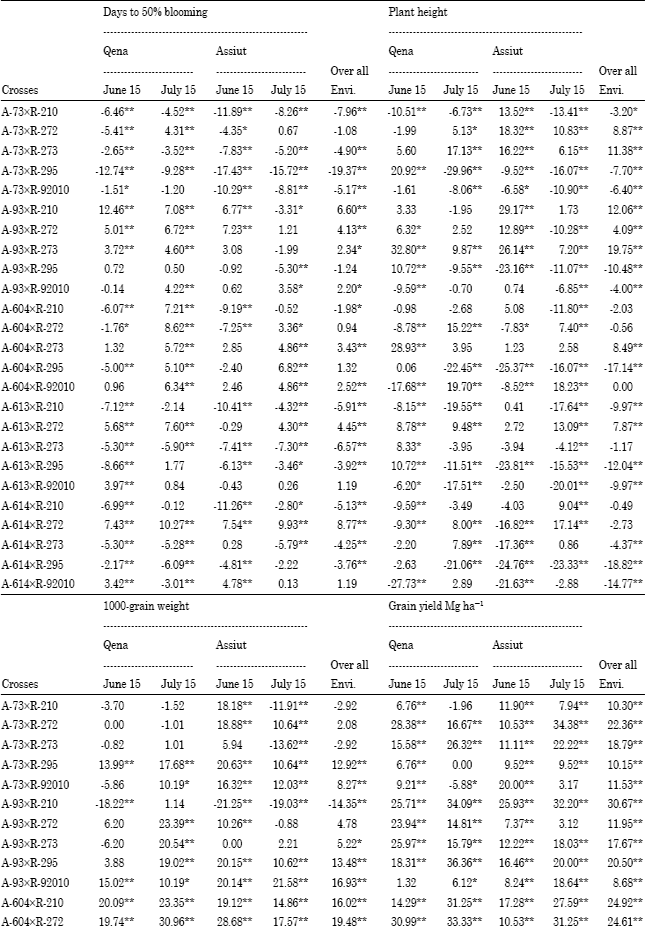
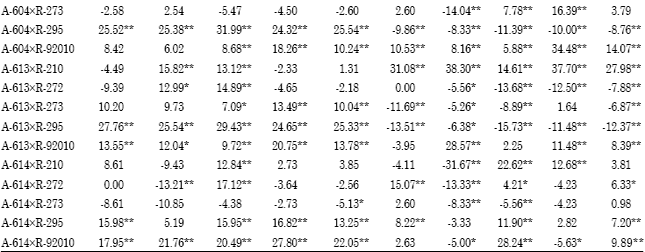
Specific combining ability (sca): Superior cross combinations were selected basedon both hybrid performance and SCA effect. Among the 25 F1 crosses, three F1 hybrids; A-73xR-295,A-93 xR-295 and A-613xR-273 showed negative and highly significant SCA effects for days to blooming under different planting dates at two locations over the two years and overall environments (Table 6).
| Table 6: | Estimates of specific combining ability effects of 25 F1crosses (SCA) for days to 50% blooming, plant height, 1000-grain weight and grain yield Mg h-1 at two locations over years and overall environments |
 | |
 | |
| *, ** Significant at 0.05 and 0.01 probability levels, respectively | |
These crosses were considered the best combinations for earliness. Two crosses; A-93xR-210 and A-604x R-92010 exhibited highly significant positive sca effects for plant height in both planting dates at two locations over the two years and overall environments. The crosses A-604xR-210, A-604xR-272 and A-614xR-92010 had significantly positive sca effects for 1000-grain weight in early and late planting in both locations over the two years and overall environments. Four crosses; A-73xR-273, A-93xR-273, A-93xR-295 and A-614xR-295 showed highly significant positive sca effects for grain yield and also gave highest grain yield in their performance under early and late planting at Qena and Assiut over the two years and overall environments (Table 6). Considering two main traits, A-93xR-295 followed by A-73xR-273 were selected as a good hybrid combinations with highly significant positive sca effect for grain yield and highly significant negative sca effect for days to blooming under early and late planting at Qena and Assiut over the two years and overall environments and suggested that this yield could be exploited commercially. These results are in agreement with those obtained by Ali (2000), Biradar et al. (2000), Kenga et al. (2004), Hovny et al. (2005), Mahmoud (2007), Hovny and El-Dsouky (2007), Abd-El-Mottaleb (2009) and Essa (2009).
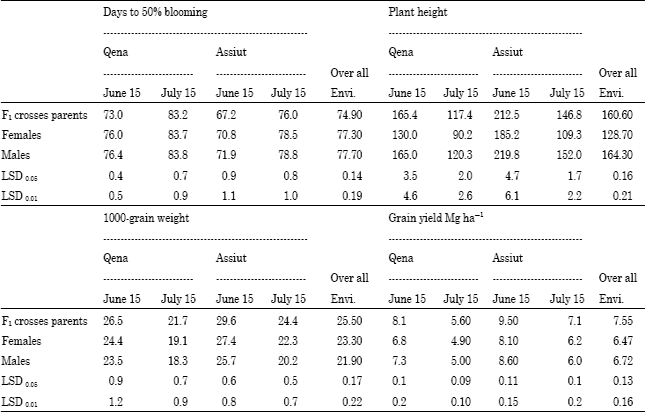
Heterosis: Average (p<0.01) than their parents; females and males under different environments except late planting at Qena (Table 7). Desirable (negative and significant) high-parent heterosis was observed for days to blooming in several hybrids under early and late planting at two locations over the two years (Table 8), showing earlier blooming than the earlier parent a locations over the two years. Over all environments, high-parent heterosis ranged from -19.37 to 8.77%. A-73xR-295 expressed desirable (i.e., negative and highly significant).
| Table 7: | Mean values for days to 50% blooming, plant height, 1000-grain weight and grain yield Mg h-1 of F1 hybrids and their parents in each planting date at two locations over years and overall environments |
 | |
earliest high-parent heterosis for days to blooming in early and late planting at two locations over the two years and over all environments as it could be expected from sca effects.
F1 hybrids were significantly taller (p<0.01) than their female parents, while they were significantly shorter (p<0.01) than their male parents under different environments except early planting at Qena (Table 7). Positive and significant high-parent heterosis was observed for some hybrids for plant hight in both dates at Qena and Assiut over the two years (Table 8). High-parent heterosis was tallest for hybrid (A-93xR-273) under early and late planting at two locations over the two years and over all environments. Over all environments, the range of high-parent heterosis for plant height was wide; between -18.82 and 19.75%.
F1 hybrids had significantly heavier average 1000-grain weight than their parents (females and males) in early and late planting at Qena and Assiut over the two years and over all environments (Table 7). Desirable (positive and significant) high-parent heterosis was observed for 1000-grain weight in several hybrids (Table 8). Over all environments, high-parent heterosis ranged from-14.35 to 25.54%. Two crosses; (A-604xR-295) and (A-613xR-295) gave positive and heaviest grain weight and high-parent heterotic values under all environments.
F1 hybrids produced higher average grain yield than their parents; females and males (Table 7). The difference was significant (p<0.01) in both dates at the two locations over the two years. The degree of high-parent heterosis differed among hybrids. Several hybrids grown in early and late planting at two locations over the two years showed advantageous high-parent heterosis (Table 8), indicating superior performance of grain yield compared to the best parent. Over all environments, the range of high-parent heterosis was wide, between-12.37 and 30.67%. The maximum high parent heterosis for grain yield of 30.67 and 27.98% were recorded for (A-93xR-210) and (A-613xR-210), respectively, between the three lowest yielding lines (A-93, A-613 and R-210).
| Table 8: | Percentage heterosis of 25 F1 crosses for days to 50% blooming, plant height, 1000-grain weight and grain yield Mg h−1 at two locations over years and overall environments |
 | |
 | |
| *, ** Significant at 0.05 and 0.01 probability levels, respectively | |
These results are in harmony with those otained by Ali (2000), Thawari et al. (2000), Prabhakar (2001), Kenga et al. (2004), Hovny et al. (2005), Abo-Zaid (2007), Hovny and El-Dsouky (2007), Abd-El-Mottaleb (2009) and Essa (2009).
CONCLUSION
Results from this study indicated that among 25 F1 hybrids and their parents under investigation, the variance components gca for both days to blooming and 1000-grain weight werelarger than those of sca under different environments over the two years and overall environments, while the variances for sca for grain yield were larger than those of gca under early and late planting at Qena and Assiut in both years. A comparable component of gca and sca were obtained for plant height. Several hybrids had significantly negative days to flowering heterosis. Grain yield yield, early maturing, taller plants and heavier grain weight, information of gca, sca and heterosis for those four traits could contribute to more efficient breeding program.
REFERENCES
- Beil, G.M. and R.E. Atkins, 1967. Estimation of general and specific combining ability in F1 hybrids for grain yield and its components in grain sorghum, Sorghum vulgare Pers 1. Crop Sci., 7: 225-228.
Direct Link - Can, N.D., S. Nakamura and T. Yoshida, 1997. Combining ability and genotype x environmental interaction in early maturing grain sorghum for summer seeding. Jpn. J. Crop Sci., 4: 698-705.
Direct Link - Hovny, M.R.A., K.M. Mahmoud, M.A. Ali and H.I. Ali, 2005. The effect of environment on performance, heterosis and combining ability in grain sorghum (Sorghum bicolor (L.) Moench). Proceedings of the 11th Conference of Agronomy, Nov. 15-16, Agronomy Dept., Faculty Agriculture, Assiut University, 689-699.
- Mode, G.J. and H.E. Robinson, 1959. Pleiotropisim and the genetic variance and co variance. Biometrics, 15: 518-537.
Direct Link - Prabhakar, 2001. Heterosis in rabi sorghum [Sorghum bicolor (L.) Moench]. Indian J. Genet. Plant Breed., 61: 364-365.
Direct Link - Kenga, R., S.O. Alabi and S.C. Gupta, 2004. Combining ability studies in tropical sorghum (Sorghum bicolor (L.) Moench). Field Crops Res., 88: 251-260.
CrossRef