
Masaru Ochiai
Faculty of Agriculture, Kagawa University, Ikenobe, Miki, Kagawa 761-0795, Japan
Tatsuhiro Matsuo
Faculty of Agriculture, Kagawa University, Ikenobe, Miki, Kagawa 761-0795, Japan
Asian Journal of Clinical Nutrition
Year: 2009 | Volume: 1 | Issue: 1 | Page No.: 1-11







ABSTRACT
This study investigated the effects of the combination of a voluntary resistance exercise (climbing) and the feeding of a high protein snack rich in Branched-Chain Amino Acids (BCAAs) on skeletal muscle weights in rats injected with glucocorticoid as a model of age-related sarcopenia (Experiment 1). Moreover, we examined whether BCAAs in the diet and/or BCAAs in snacks prevent sarcopenia in sedentary rats (Experiment 2). Male Wistar rats were injected with prednisolon (2 mg kg-1) every day. Rats in the exercise groups climbed voluntarily in the tower cage for 8 weeks. BCAAs were exchanged partly for casein in the experimental diets. Glucocorticoid injection decreased final body weight and muscle mass. The body weight gain did not differ among glucocorticoid-injected rats. Climbing exercise markedly prevented the loss of muscle mass, but the BCAA snack did not facilitate the effects of exercise (Experiment 1). In the sedentary rats, either BCAA in the diet or BCAA in the snack decreased mass and protein content of muscle (Experiment 2). These results suggest that resistance exercise is the most effective way to inhibit sarcopenia in rats. BCAAs did not depress muscle mass loss without resistance exercise. BCAAs should be used on the condition that all other essential amino acids are fully available in the diet.
PDF Abstract XML References Citation
How to cite this article
Masaru Ochiai and Tatsuhiro Matsuo, 2009. Dietary BCAAs Do Not Prevent Skeletal Muscle Atrophy in Rats Injected
with Glucocorticoid. Asian Journal of Clinical Nutrition, 1: 1-11.
DOI: 10.3923/ajcn.2009.1.11
URL: https://scialert.net/abstract/?doi=ajcn.2009.1.11
DOI: 10.3923/ajcn.2009.1.11
URL: https://scialert.net/abstract/?doi=ajcn.2009.1.11
INTRODUCTION
Aging is a process that all humans eventually undergo. Skeletal muscle is a good indicator of age-related decline in the physiological functions. It has been reported that the number of muscle fibers in humans 20-30 years old declines to about 39% by 80-90 years old (Lexell et al., 1988). Sarcopenia, a serious problem in elderly people, is characterized by atrophy of the muscles, which results in decreasing energy metabolism and physical activity. The mechanisms leading to sarcopenia are still unclear and result from an imbalance between the rates of protein synthesis and degradation in the skeletal muscles (Balagopal et al., 1997; Hasten et al., 2000). Reduction of energy expenditure decreases dietary protein intake. However, proteosynthesis activity in splanchnic tissues in elderly people is higher (Boirie et al., 1997) and protein metabolisms in splanchnic tissues become more and more active in elderly people (Young, 1990). Therefore, most amino acids are absorbed from the digestive tract and extracted by splanchnic tissues, especially liver (Boirie et al., 1997). This absorption could result in a lower availability of dietary amino acids in the skeletal muscles, which would reduce muscle proteosynthesis and lead to sarcopenia (Savery et al., 1998; Kang et al., 2002). Protein supplementation with high-insulinogenic carbohydrates after meals could increase the amino acid supply to peripheral tissues, especially bone and muscles (Gaudichon et al., 1999; Matsuo and Suzuki, 2005). The protein supplementation pattern was important in the effective maintenance of the amount of nitrogen in elderly women (Arnal et al., 1999), but not in young women (Arnal et al., 2000). We also previously reported that the intake of high protein snacks (60% casein) 3 h after a regular meal increased the total blood amino acid flow (Matsuo and Suzuki, 2005). On the other hand, BCAAs play a major role in the suppression of skeletal muscle proteolysis (Anthony et al., 1999; Bigard et al., 1996; MacLean et al., 1994, 1996; Mourier et al., 1997). BCAAs account for about 35% of the essential amino acids in muscle protein and about 50% of the essential amino acids in dietary protein (Harper et al., 1984). It has been believed that BCAAs are oxidized mainly in skeletal muscles, whereas other essential amino acids are catabolized mainly in the liver (Ichihara et al., 1981; Miller, 1962). Moreover, leucine stimulates muscle protein synthesis and plays an important role in the regulation of postprandial muscle protein metabolism (Dardevet et al., 2002; Arnal et al., 2002; Rieu et al., 2003; Combaret et al., 2005). Therefore, dietary protein rich in BCAAs might suppress the sarcopenia induced by aging.
Resistance exercise is one of the most effective ways to prevent the sarcopenia associated with aging, because it facilitates the secretion of growth hormone (Fluckey et al., 1996; Ortoft et al., 1998). We previously reported that voluntary climbing exercise increased skeletal muscle weight (Matsuo and Suzuki, 2004, 2006; Kang et al., 2002). However, we have not yet demonstrated an interaction between resistance exercise and dietary BCAAs in preventing age-related muscle atrophy.
In the present study, we investigated the effects of voluntary resistance exercise (climbing) together with the intake of high protein snacks rich in BCAAs on skeletal muscle weights in rats injected with glucocorticoid as a model of age-related sarcopenia (Experiment 1). We further examined whether BCAAs in the diet and/or BCAAs in snacks prevented sarcopenia in sedentary rats (Experiment2).
MATERIALS AND METHODS
All procedures involving rats were approved by the Experimental Animal Care Committee of Kagawa University.
Animals and Experiment Design
Experiment 1: Effect of Voluntary Resistance Exercise Together with the Intake of a High Protein Snack Rich in BCAAs Feeding on Skeletal Muscle Weights in Glucocorticoid-Injected Rats
Fifty-five male Wistar rats (age 5 weeks) were purchased from Japan SLC (Shizuoka, Japan). All rats were housed individually at 25±1°C with light from 8:00-20:00 and free access to water. All rats were fed commercial rat chow (CE-2, Japan CLEA, Tokyo, Japan) for 3 days ad libitum, then they were fed meals twice a day (8:30-9:30, 20:30-21:30) for 4 weeks. During the 4 weeks, all rats were housed in metal cages with a wire mesh tower (![]() 20x200 cm) to adjust climbing exercise between 21:30 and 8:00 h in the same way as in the previous experiment (Matsuo et al., 2003). At the age of 9 weeks, all rats were divided into 7 groups. The control group (Ct, n = 8) was injected with 2 mL kg-1 day-1 of saline and the other groups were injected with 2 mg kg-1 day-1 prednisolon (SIGMA-Aldrich, MO, USA), which is a kind of glucocorticoid. The other groups were the glucocorticoid-injected sedentary group (GS, n = 8); the glucocorticoid-injected sedentary with snack (60% casein) group (GSc, n = 8); the glucocorticoid-injected sedentary with snack rich in BCAAs group (GSb, n = 7); the glucocorticoid-injected climbing group (GE, n = 8); the glucocorticoid-injected climbing with snack (60% casein) group (GEc, n = 8) and the glucocorticoid-injected climbing with snack rich in BCAAs group (GEb, n = 8). The experiment was carried out for 8 weeks. The composition of each diet is shown in Table 1. All groups were fed the meal twice a day (8:30-9:30, 20:30-21:30 h). The snack groups were fed the casein or BCAA snack from 12:30 to13:30 h. The ratio of BCAAs (valin, leucine and isoleucine) was 1:2:1, respectively (Table 2) (De Palo et al., 2001).
20x200 cm) to adjust climbing exercise between 21:30 and 8:00 h in the same way as in the previous experiment (Matsuo et al., 2003). At the age of 9 weeks, all rats were divided into 7 groups. The control group (Ct, n = 8) was injected with 2 mL kg-1 day-1 of saline and the other groups were injected with 2 mg kg-1 day-1 prednisolon (SIGMA-Aldrich, MO, USA), which is a kind of glucocorticoid. The other groups were the glucocorticoid-injected sedentary group (GS, n = 8); the glucocorticoid-injected sedentary with snack (60% casein) group (GSc, n = 8); the glucocorticoid-injected sedentary with snack rich in BCAAs group (GSb, n = 7); the glucocorticoid-injected climbing group (GE, n = 8); the glucocorticoid-injected climbing with snack (60% casein) group (GEc, n = 8) and the glucocorticoid-injected climbing with snack rich in BCAAs group (GEb, n = 8). The experiment was carried out for 8 weeks. The composition of each diet is shown in Table 1. All groups were fed the meal twice a day (8:30-9:30, 20:30-21:30 h). The snack groups were fed the casein or BCAA snack from 12:30 to13:30 h. The ratio of BCAAs (valin, leucine and isoleucine) was 1:2:1, respectively (Table 2) (De Palo et al., 2001).
| Table 1: | Composition of experimental diets (experiment 1) |
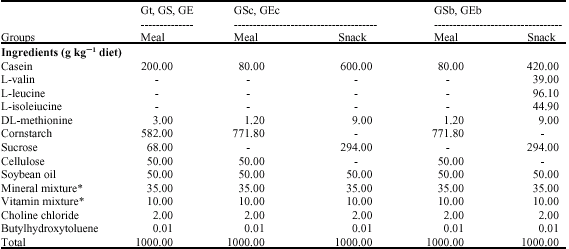 | |
| *: Based on AIN-76 mixture; Ct: Saline-injection; G: Glucocoritcold-injection; S: Sedentary; E: Climbing exercise: c: Casein; b: BCAA | |
| Table 2: | Composition of experimental diets (experiment 2) |
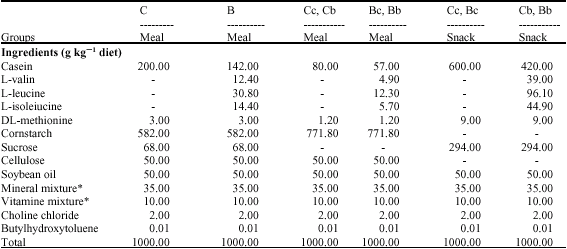 | |
| *: Based on AIN-76 mixture; C and c, Casein; B and b, BCAA | |
The additional quantities of BCAAs in the snack groups were determined by the quantities of BCAAs in casein. The GE, GEc and GEb groups exercised voluntarily in tower-climbing cages from 21:30 to 8:00 h for 8 weeks.
In the 6th or 7th week of the experimental period, in order to determine diurnal variations in the plasma urea nitrogen concentrations, blood was taken from the tail arteries of all rats using heparinized tubes at 8:00, 12:00, 16:00 and 20:00 h. The plasma was recovered from whole blood samples by centrifugation at 3000 rpm for 5 min and stored at -20°C until analysis.
At the end of the experiment, the rats were killed by decapitation at 10:00 h after overnight fasting. Blood was collected and centrifuged at 3000 rpm for 15 min in order to obtain serum. The serum was stored at -20°C. The liver, heart and skeletal muscles (soleus, plantaris, gastrocumemius, tibialis anterior and extensor digitorum longus) were quickly removed, weighed and stored at -80°C until the analysis was performed.
Experiment 2
Effects of BCAAs in Diet and/or Snacks on Skeletal Muscles in Glucocorticoid-Injected Sedentary Rats
Fifty male Wistar rats (age 8 weeks) were purchased from Japan SLC. All rats were fed with CE-2 for 3 days ad libitum and then were fed meals twice a day (8:30-9:30, 20:30-21:30 h) for a week. All rats were divided into six groups: a casein diet group (C, n = 9), a casein diet and casein snack group (Cc, n = 8), a casein diet and BCAA snack group (Cb, n = 8), a BCAA diet group (B, n = 9), a BCAA diet and casein snack group (Bc, n = 8) and a BCAA diet and BCAA snack group (Bb, n = 8). All groups were injected subcutaneously with 2 mg kg-1 day-1 of prednisolon at 9:30 h. The composition of each diet is shown in Table 2. Each group was fed each meal twice a day (8:30-9:30, 20:30-21:30 h). The Cc, Cb, Bc and Bb groups were fed with casein or BCAA snack from 12:30 to13:30. The ratio of BCAAs (valin, leucine and isoleucine) was the same as in Experiment 1. At the end of the experiment, the rats were killed in the same manner as in Experiment 1.
Measurements
Protein contents in the heart, liver, soleus muscle and tibialis anterior muscle were determined by the Kjeldahl technique using a Tecator Digestor Auto and Kjeltec 2400 (Foss Japan, Tokyo, Japan). Plasma urea nitrogen concentrations were determined using a kit (UN B-test, Wako Pure Chemical Industries, Osaka, Japan).
Statistical Analysis
In Experiment 1, the values were expressed as means±SD. Data were assessed by one-way ANOVA and Fishers PLSD test. In Experiment 2, the values were expressed as means±SD. Data were assessed by two-way ANOVA for the diet and snack. When the treatment was found to have a significant effect, the difference between the experimental groups was assessed by Fishers PLSD test. Statistical significance was set at less than 0.05.
RESULTS
Experiment 1
Body Weight and Food Intake
The final body weight of the Ct group was the greatest among all groups. Final body weight was not significantly different among the glucocorticoid-injected groups. The Ct group consumed the most food. Total intake by the GS group was significantly lower than that of the GE group. Protein intake by the GS group was the lowest of all groups except for the GEb groups. Energy intake in the control and GE groups was significantly higher than that of the GS group (Table 3).
Tissue Weights
Heart and liver weights were not influenced by climbing or snack. However, heart weight was decreased by glucocorticoid injection. The relative weights of the heart and liver in the GS group were significantly lower than those of the Ct group. The ratio (mg g-1) of heart weight to body weight was 2.25 in the Ct group and 2.56 in the GS group. The ratio (mg g-1) of liver weight to body weight in the Ct group was 25.2 while that in the GS group was 31.1. These differences were significant at the p<0.05 level (Table 4).
| Table 3: | Final body weight and food intake of each group of rats (experiment 1) |
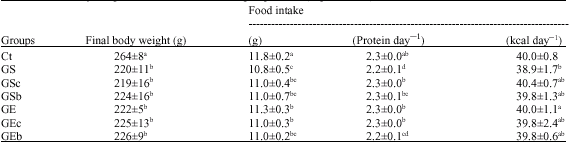 | |
| Values are means±SD for 7-8 rats; Ct: Saline-injection; G: Glucocoritcold-injection; S: Sedentary; E: Climbing exercise; c: Casein; b: BCAA snack; Different letter(s) within a column represent significant differences at p<0.05 as calculated by one-way ANOVA and Fishers PLSD tests | |
| Table 4: | Heart, liver and hindlimb muscles weights in each group of rats (experiment 1) |
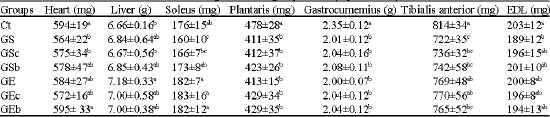 | |
| Values are means±SD for 7-8 rats Ct: Saline-injection; G: Glucocoritcold-injection; S: Sedentary; E: Climbing exercise; c: Casein; b: BCAA snack; Different letter(s) within a column represent significant differences at p<0.05 as calculated by one-way ANOVA and Fishers PLSD tests | |
| Table 5: | Diurnal variations of plasma urea nitrogen concentration at 6-8th weeks (experiment 1) |
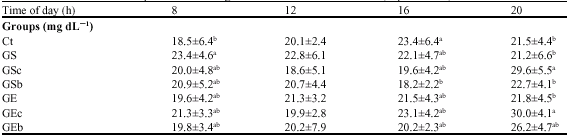 | |
| Values are means±SD for 7-8 rats; Ct, Saline-injection; G, Glucocoritcold-injection; S, Sedentary; E, Climbing exercise; c, Casein; b, BCAA snack; Different letter(s) within a column represent significant differences at p<0.05 as calculated by one-way ANOVA and Fishers PLSD tests | |
All skeletal muscle weights were significantly decreased by glucocorticoid injection. Soleus and tibialis anterior muscle weights were significantly increased by climbing, but were not influenced by snacks. Soleus muscle weight in the GSb group was significantly greater than in the GS group. However, the differences were not significant between the GS and GSc groups, or between the GSc and GSb groups. Tibialis anterior muscle weight in the GE and GEc groups was significantly higher than that in the GS group. However, plantaris, gastrocunemius and extensor digitorum longus muscle weights were not affected by climbing or snacks.
Plasma Urea Concentrations
Plasma urea concentrations at 8:00, 12:00, 16:00 and 20:00 h in the 6-7th weeks are shown in Table 4. At 8:00 h, the plasma urea concentration of the Ct group was significantly lower than that of the GS group. At 16:00 h, the plasma urea concentrations of the Ct and the GEc groups were higher than that of the GSb group. Moreover, at 20:00, the plasma urea concentrations of the GSc and GEc groups were significantly higher than those of the other groups, except for the GEb group (Table 5).
Experiment 2
Body Weight and Dietary Intake
Although the differences in body weight, dietary intake and energy intake among all groups were not significant, the protein intake of the Bc group was higher than those in the C and B groups (Table 6).
Tissue Weights
In the factor analysis, heart and liver weights were not influenced by either diet or snack. The soleus muscle weight in the B group was significantly lower than that in the C group. The weights of the Bc and Bb groups were lower than those of the other groups except for the Cc group. Tibialis anterior muscle weight in the Bc group was lower than that in the C and Cb groups.
| Table 6: | Final body weight and food intake of each group of rats (experiment 2) |
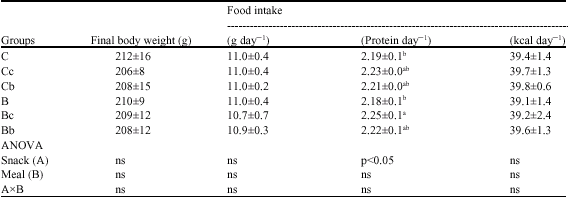 | |
| Values are means±SD for 7-8 rats; Ct: Saline-injection; G: Glucocoritcold-injection; S: Sedentary; E: Climbing exercise; c: Casein; b: BCAA snack; Different letter(s) within a column represent significant differences at p<0.05 as calculated by one-way ANOVA and Fishers PLSD tests; ns: non significant | |
| Table 7: | Heart, liver and hindlimb muscle weight in each group of rats (experiment 2) |
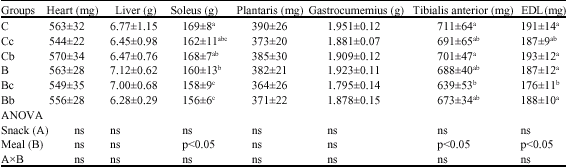 | |
| Values are means±SD for 7-8 rats; Ct: Saline-injection; G: Glucocoritcold-injection; S: Sedentary; E: Climbing exercise; c: Casein; b: BCAA snack; Different letter(s) within a column represent significant differences at p<0.05 as calculated by one-way ANOVA and Fishers PLSD tests; ns: non significant | |
| Table 8: | Protein content in heart, liver and hindlimb muscles in each group of rats (experiment 2) |
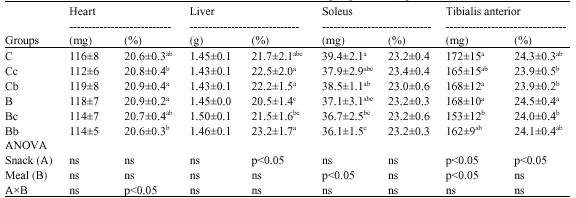 | |
| Values are means±SD for 7-8 rate; Ct: Saline-injection; G: Glucocoritcold-injection; S: Sedentary; E: Climbing exercise; c: Casein; b: BCAA snack; Different letter(s) within a column represent significant differences at p<0.05 as calculated by one-way ANOVA and Fishers PLSD tests; ns: non significant | |
EDL muscle weight in the Bc group was lower than that in the other groups except for Cc group. However, plantaris and gastrocunemius muscle weights were not affected by any of the diets or snacks (Table 7).
Tissues Protein Contents
The percentages of liver protein in the B and Bc groups were lower than those in the Cc, Cb and Bb groups. On the other hand, liver protein content did not differ among any of the groups (Table 8).
DISCUSSION
In Experiment 1, the results revealed that final body weight and skeletal muscle weight were decreased as a result of glucocorticoid injection. These results agreed with our previous findings (Matsuo and Suzuki, 2004, 2005). In the case of the plasma urea nitrogen concentration, the value at 8:00 was higher in the GS group than in the Ct group. These results support the previously reported effect of glucocorticoid. Glucocorticoid is well known to be a hormone which not only enhances skeletal muscle protein degradation but also reduces skeletal muscle protein synthesis (Rannels and Jefferson, 1980; Odedra et al., 1983; Dardevet et al., 1995, 1998; Minet-Quinard et al., 2000; Shah et al., 2000). Also, glucocorticoid is well known to stimulate gluconeogenesis in the liver (Plaschke et al., 1996). These findings indicated that not only endogenous body protein and amino acids but also exogenous body protein and amino acids could be utilized in splanchnic organs, especially in the liver, resulting in proteosynthesis. In fact, in Experiment 1, the relative weight of the liver was higher in the GS group than in the Ct group (Ct vs. GS; 25.2 vs. 31.1 mg g-1 body weight, p<0.05), indicating that the function in the liver was activated with aging.
On the other hand, the effect of resistance exercise on the suppression of the loss of skeletal muscle weight was not found with or without BCAA snack in Experiment 1 because resistance exercise alone was sufficient to inhibit sarcopenia. These results indicated that resistance exercise might be one of the strongest factors in the suppression of body protein degradation. On the other hand, soleus muscle weight in the GSb group was higher than that in the GS group. These results suggest that the increase in soleus muscle weight was caused by the BCAA snack in the sedentary condition. The effectiveness of BCAAs in elderly people as well as young people has been brought to international attention (Andrea and Nair, 2006; Satoshi and Volpi, 2006). It has been believed that sarcopenia could be inhibited in elderly people if BCAA supplements and/or food rich in BCAAs were ingested. It has been reported that BCAAs consist of about 35% of the essential amino acids in proteins and about 40% of the preformed amino acids required by mammals (Harper et al., 1984). Rennie (1996) reported that BCAAs could be oxidized mainly in skeletal muscles, whereas other essential amino acids were mainly oxidized in the liver. Thus it is believed that BCAAs are closely related with skeletal muscle protein metabolism (Rennie, 1996).
The anabolic response of whole body protein turnover was normalized in the elderly if a protein-pulse feeding pattern (80% of daily protein in 1 meal) was used instead of spread-protein feeding (daily proteins equally distributed). This finding suggested that a large amount of amino acids (especially essential amino acids) may be needed in the elderly to obtain the same anabolic effect observed in the young (Arnal et al., 1999). In our previous study, casein snack (60% protein) feeding after a meal increased serum essential amino acid and BCAA concentrations throughout the day in comparison with no snack feeding (Matsuo and Suzuki, 2005). Moreover, it was also reported that casein protein enhanced plasma BCAA concentration more than soy protein in healthy humans (Luiking et al., 2005). Therefore, it has been believed that pulse-protein feeding and casein have greater effects on muscle proteosynthesis.
Dietary and endogenous proteins were finally degraded to urea. Therefore, the diets and snacks would affect the urea concentration. Table 5 showed that the plasma urea nitrogen concentration in the GSc, GSb, GEc and GEc groups, which were casein-or BCAA-snack-intake groups, generally increased after the snack intake and the highest value in each group was seen at 20:00 h (7.5 h after the snack). The plasma urea nitrogen concentration results at 20:00 h were highest in the GSc and GEc groups and similar diurnal patterns were observed in the GSc and GEc groups. On the other hand, the plasma urea nitrogen concentration in the GSb group was lower than in the casein snack groups (GSc and GEc). These results indicated that the lowest amino acids derived from the BCAA snacks were degraded in splanchnic tissues and most amino acids were delivered to skeletal muscles. As a result, the soleus muscle weight in the GSb group might be influenced by BCAA snacks. However, the increased muscle weight observed with BCAA snacks in the GSb group (Experiment 1) was not found in Experiment 2. Therefore, it is not necessarily appropriate to suggest that BCAA snacks could lead to the suppression of sarcopenia. Several explanations for why the BCAA snacks may not lead to the suppression of sarcopenia must be considered.
First, the most crucial problem was that protein intake in the GS group was the lowest, lower than in the GSb group, which resulted in the loss in the soleus muscle weight as seen in the GS group.
Second, the positive effect of the BCAA snack was seen only in soleus muscle, but not in glycolytic and oxidative glycolytic muscles (plantaris, gastrocnemius, tibialis anterior, EDL). Taking these results into consideration, the effect of the BCAA snack was not surprisingly high and the BCAA snack could not always lead to suppression of the sarcopenia induced by aging.
On the other hand, BCAAs had negative effects on muscle weight and protein content in the soleus and tibia anterior muscles in the BCAA-diet-ingestion groups, especially the Bc and Bb groups in Experiment 2. These results conflict with previous reports that BCAAs are metabolized mainly in skeletal muscles (Ichihara et al., 1981; Miller, 1962) and that BCAA supplementation suppresses skeletal muscles proteolysis (Anthony et al., 1999; Bigard et al., 1996; MacLean et al., 1994, 1996; Mourier et al., 1997). It is possible that these results were due to a worse balance of amino acids in the BCAA diet. It was expected that the essential amino acid intake except for BCAAs would be lowest in the GBb group and that the total amino acid balance in the GBb group might not have sufficiently met the amino acid requirement for muscle proteosynthesis in Experiment 1. However, in the present study, we thought it was more important that the free amino acid concentration increased and that more free amino acids arrived in the skeletal muscles than with the dietary amino acid balance. Another reason for the negative result may be that the amount of protein per meal in the snack feeding groups was rather low (15%). This protein ratio might not sufficiently meet the amino acid requirement for the proteosynthesis in splanchnic tissues. However, a previous study showed that even a 15% protein (casein) ratio in a meal met the amino acid requirement for proteosynthesis in skeletal muscle (Matsuo and Suzuki, 2006).
In addition, it should be considered that glucocorticoid can stimulate gluconeogenesis in the liver (Plaschke et al., 1996). It was reported that glucocorticoid stimulated the degradation of BCAAs by activating branched-chain á-keto acid dehydrogenase complex, a limited enzyme of BCAA degradation (Huang et al., 1999). Therefore, more BCAAs in the BCAA snack might be catabolized by glucocorticoid injection, which indicates that a lower supply of other essential amino acids to peripheral tissue might have occurred and suppressed muscle protein synthesis in Experiment 2.
In conclusion, routine resistance exercise and adequate dietary protein were the most effective ways to inhibit sarcopenia related with aging. On the other hand, a higher BCAA content in the diet and in snacks did not prevent skeletal muscle atrophy and sarcopenia in the sedentary condition. BCAA should be used on the condition that all other essential amino acids are fully available.
REFERENCES
- Andrea, T. and K.S. Nair, 2006. Assessment of branched-chain amino acid status and potential for biomarkers. J. Nutr., 136: 324S-330S.
PubMedDirect Link - Anthony, J.C., T.G. Anthony and D.K. Layman, 1999. Leucine supplementations enhance skeletal muscle recovery in rats following exercise. J. Nutr., 129: 1102-1106.
PubMedDirect Link - Arnal, M.A., L. Mosoni, Y. Boirie, M.L. Houlier, L. Morin, E. Verdier and P. Ritz et al., 1999. Protein pulse feeding improves protein retention in elderly women. Am. J. Clin. Nutr., 69: 1202-1208.
PubMedDirect Link - Arnal, M.A., L. Mosoni, Y. Boirie, M.L. Houlier, L. Morin, E. Verdier, P. Ritz, J.M. Antoine, J. Prugnaud, B. Beaufrere and P.P. Mirand, 2000. Protein feeding pattern does not affect protein retention in young women. J. Nutr., 130: 1700-1704.
PubMedDirect Link - Arnal, M.A., L. Mosoni, D. Dardevet, M.C. Ribeyre, G. Bayle, J. Prugnaud and P.P. Mirand, 2002. Pulse protein feeding pattern restores stimulation of muscle protein synthesis during the feeding period in old rats. J. Nutr., 132: 1002-1008.
Direct Link - Balagopal, P., O.E. Rooyackers, P.A. Adey and K.S. Nair, 1997. Effects of aging on in vivo synthesis of skeletal muscle myosin heavy-chain and sarcoplasmic protein in humans. Am. J. Physiol., 273: 790-800.
PubMedDirect Link - Bigard, A.X., P. Lavier, L. Ullmann, H. Legrand, P. Douce and C.Y. Guezennec, 1996. Branched chain amino acid supplementation during repeated prolong skiing exercise at altitude. Int. J. Sports Nutr., 6: 295-306.
PubMed - Combaret, L., D. Dardevet, I. Rieu, M.N. Pouch, D. Bechet, D. Taillandier, J. Grizard and D. Attaix, 2005. A leucine-supplemented diet restores the defective postprandial inhibition of proteasome-dependent proteolysis in aged rat skeletal muscle. J. Physiol., 569: 489-499.
CrossRefPubMedDirect Link - Dardevet, D., C. Sornet, D. Taillandier, I. Savary, D. Attaix and J. Grizard, 1995. Sensitivity and protein turnover response to glucocorticoids are different in skeletal muscle from adult and old rats. Lack of regulation of the ubiquitin-proteasome proteolytic pathway in aging. J. Clin. Invest., 96: 2113-2119.
CrossRefPubMed - Dardevet, D., C. Sornet, G. Bayle, J. Prugnaud, C. Pouyet and J. Grizard, 2002. Postprandial stimulation of muscle protein synthesis in old rat can be restored by a leucine-supplemented meal. J. Nutr., 132: 95-100.
PubMedDirect Link - De Palo, E.F., R. Gatti, E. Cappellin, C. Schiraldi, C.B. De Palo and P. Spinella, 2001. Plasma lactate, GH and GH-binding protein levels in exercise following BCAA supplementation in athletes. Amino Acids, 20: 1-11.
CrossRefPubMedDirect Link - Fluckey, J.D., T.C. Vary, L.S. Jefferson, W.J. Evans and P.A. Farrell, 1996. Insulin stimulation of protein synthesis in rat skeletal muscle following resistance exercise is maintained with advancing age. J. Gerontol. A. Bio. Sci. Med. Sci., 51: 323-330.
PubMed - Gaudichon, C., S. Mahe, R. Benamouzig, C. Luengo and H. Fouillet et al., 1999. Net postprandial utilization of [15N]-labeled milk protein nitrogen is influenced by diet composition in humans. J. Nutr., 129: 890-895.
PubMedDirect Link - Harper, A.E., R.H. Miller and K.P. Block, 1984. Branched-chain amino acid metabolism. Annu. Rev. Nutr., 4: 409-454.
CrossRefPubMedDirect Link - Kang, H., T. Matsuo, R. Choue and M. Suzuki, 2002. Effect of voluntary resistance exercise on heme biosynthesis in rats given glucocorticoid-injections. Biosci. Biotechnol. Biochem., 66: 2710-2714.
CrossRefPubMedDirect Link - Luiking, C.Y., N. Deutz, M. Jaekel and P. Soeters, 2005. Casein and soy protein meals differentially affect whole-body and splanchnic protein metabolism in healthy humans. J. Nutr., 135: 1080-1087.
PubMedDirect Link - Matsuo, T., T. Nozaki, K. Okamura, K. Matsumoto, T. Doi, S. Gohtani and M. Suzuki, 2003. Effect of voluntary resistance exercise and high-protein snack on bone mass, composition and strength in rats given glucocorticoid injections. Biosci. Biotechnol. Biochem., 67: 2518-2523.
CrossRefPubMedDirect Link - Mourier, A., A.X. Bigard, E. De Kerviler, B. Roger H. Legrand and C.Y. Guezennec, 1997. Combined effects of caloric restriction and branched-chain amino acid supplementation on body composition and exercise performance in elite wrestlers. Int. J. Sports Med., 18: 47-55.
CrossRefPubMedDirect Link - Rieu, I., C. Sornet, G. Bayle, J. Prugnaud, C. Pouyet, M. Balage, I. Papet, J. Grizard and D. Dardevet, 2003. Leucine-supplemented meal feeding for ten days beneficially affects postprandial muscle protein synthesis in old rats. J. Nutr., 133: 1198-1205.
PubMedDirect Link - Satoshi, F. and E. Volpi, 2006. Amino acids and muscle loss with aging. J. Nutr., 136: 277S-280S.
PubMedDirect Link