
Ozlem (Dalgic) Aksoy
Department of Biology, Faculty of Science and Arts, Kocaeli University, 41380, Umuttepe, Izmit, Kocaeli, Turkey
Nuran Ekici
Department of Science Education, Faculty of Education, Trakya University, 22030, Aysekadin Yerleskesi, Edirne, Turkey
Feruzan Dane
Department of Biology, Faculty of Science and Arts, Trakya University, 22030, Edirne, Turkey
Asian Journal of Cell Biology
Year: 2008 | Volume: 3 | Issue: 1 | Page No.: 34-40







ABSTRACT
In this study the genotoxic effects of Fusilade (Fluozifop-p-butyl), a selective herbicide were investigated in the mitotic cell division in lentil (Lens culinaris Medik.) root tip cells. For this aim, different doses (0.25, 5, 1 and 1.5%) of Fusilade solutions were used. Root tips after having grown to a certain length were stained according to aceto orcein squash procedure and the number of abnormal cells was counted in each phase of mitosis. The obtained results indicate that Fusilade had the ability to cause production of a large number of mitotic abnormalities. These abnormalities appeared in varying degrees depending on the dose. The types of abnormalities produced were laggards of the chromosomes, bridges, c-metaphase, stickiness, chromatin granulation and nucleus deformation. According to our findings we can say that Fusilade can produce negative effects on mitotic divisions in lentil root tip cells.
PDF Abstract XML References Citation
How to cite this article
Ozlem (Dalgic) Aksoy, Nuran Ekici and Feruzan Dane, 2008. Mitotic Changes in Root Meristems of Lens culinaris Treated with Fusilade (fluazifop-p-butyl). Asian Journal of Cell Biology, 3: 34-40.
DOI: 10.3923/ajcb.2008.34.40
URL: https://scialert.net/abstract/?doi=ajcb.2008.34.40
DOI: 10.3923/ajcb.2008.34.40
URL: https://scialert.net/abstract/?doi=ajcb.2008.34.40
INTRODUCTION
Herbicides are metabolic inhibitors and their modes of action can be classified into different groups; photosynthetic inhibitors, cell growth disruptors (mitotic inhibitors), growth regulators, lipid biosynthesis inhibitors, carotenoid biosynthesis inhibitors, branch chain amino acid inhibitors etc. (Hess, 1999). Fluazifop-p-butyl is a post-emergence phenoxy herbicide. It is absorbed rapidly through leaf surfaces and quickly hydrolyzes to fluazifop acid. The acid is transported primarily in the phloem and accumulates in the meristems where it disrupts the synthesis of lipids in susceptible species (Urano, 1982; Erlingson, 1988).
Treatment with Fusilade have been shown to be an effective method of controlling annual grasses and promoting establishment of seeded coastal sage scrub vegetation (Cione et al., 2002). In Turkey Fusilade is used especially for weed control in lentil, soy bean, cotton, sugar beet and tomatoes farms (Syngenta, 2007. http://www.syngenta.com.tr/ilaclar). Although the field use of herbicides has now become a common practice in leguminous plant cultivation, there is not much information on their cytological effects. Phytotoxic and genotoxic effects of different pesticides have been determined in different organisms (Sinha, 1989; Aktac et al., 1994; Dane and Dalgic, 2005; Tartar et al., 2006). But the studies about the effects of Fusilade are limited with the observations mostly in animal cells. Acute and chronic toxicity of Fusilade was widely studied on birds, mammals, fishes and fungus (Abdel-Mallek et al., 1996; Gorlach-Lira et al., 1997). During present research we found studies about Fusilade effectiveness, but little information is available about its cellular effects on different plants. More investigations are necessary to demonstrate precisely the effects of such chemicals and establish their in situ levels of toxicity.
The cytogenetic effects of the herbicide Avenoxan, active substance 2,4-D, were investigated in both Allium cepa L. and Allium sativum L. (Tartar et al., 2006). Different kinds of mitotic changes were observed on Allium cepa root tips treated with the herbicide flurochloridone (Yuzbasioglu et al., 2003) and with the herbicide Logran on root tip cells of Triticum aestivum L. and Hordeum vulgare L. (Kaymak and Muranli, 2006). It was found that the herbicide linuron had a marked mitodepressive action on mitosis of Helianthus annuus L. (Inceer et al., 2004).
The negative effects of Fusilade on mitotic index and α-amylase content of lentil seeds were observed by our previous study (Aksoy Dalgic et al., 2007). The present study has been carried out to investigate the influence of Fusilade in lentil (Lens culinaris Medik.) root tip cells during mitosis.
MATERIALS AND METHODS
This study was conducted between years 2003-2005 in cell biology laboratories of Trakya University. Seeds of Lens culinaris Medik. cv. Sultan were pretreated with 5% NaOCl for 10 min for seed surface sterilization and then placed under clean bench conditions in Petri dishes and filled with dH2O at room temperature for germination. The herbicide used was butyl 2-[4-[5- (trifluoromethyl)pyridin-2-yl]oxyphenoxy]propanoate and chemical formula of this molecule is C19H20F3NO4 (http://www.iupac.org/). The Fusilade doses used in this study was based on the doses which are used in field. The field dose (0.25%) and its higher doses were used for treatment (Ouzounidou et al., 1997). The control groups were treated only with modified Hoagland nutrient solution. The root tips which were 0.5 mm and higher were determined as germinated. All experiments were done in three repetition and 100 seeds were used for each repeat. The germination of seeds was observed in laboratory conditions in petri dishes for 5 days. The root tips of lentils were fixed in Carnoy fixative (3 alcohol:1 acetic acid) and hydrolyzed in 1 N HCl at 50°C for 5 min followed by squashing in a 2% aceto orcein stain.
Slides were kept at +4°C in a freezer and examined within a month. The evaluation of chromosomal structural changes is performed after Fusilade application. The numbers of abnormality were counted in each phase of mitosis. The abnormalities were characterized and classified in the following categories: fragments, bridges, lagging chromosome, c-metaphase, stickiness, chromatin granulation and nucleus deformation. The photographs were taken by an Olympus photomicroscope.
RESULTS
The Ratio of Abnormalities During Mitosis
The ratio of abnormalities of lentil root tip cells during mitosis is shown in Table 1 and 2. From the Table 1 and 2 it can be seen that the ratio of abnormalities increases parallel to the increase in doses of Fusilade (Fig. 1). The ratio of abnormalities was determined by counting the abnormal cells in each phase of mitosis.
| Table 1: | The effect of Fusilade doses to prophase and metaphase |
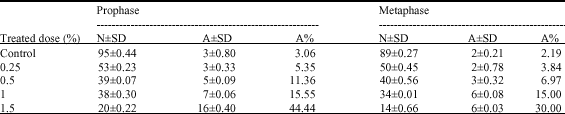 | |
| N: The number of normal cells, A: The number of abnormal cells, A%: The abnormal cell ratio, ±SD: Standard deviation | |
| Table 2: | The effect of Fusilade doses to anaphase and telophase |
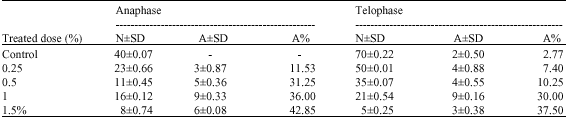 | |
| N: The number of normal cells, A: The number of abnormal cells, A%: The abnormal cell ratio, ±SD: Standard deviation | |
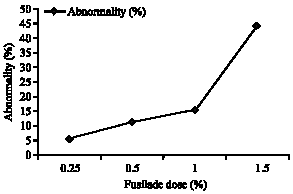 | |
| Fig. 1: | The effect of different doses of Fusilade on abnormality percentage |
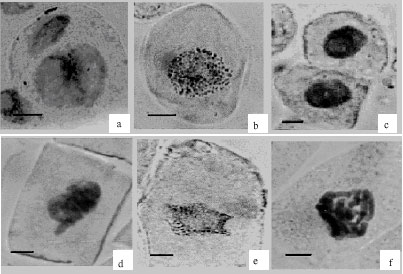 | |
| Fig. 2: | The abnormalities in interphase and prophase cells in Lens culinaris root meristematic cells (a) Two nuclei in different size, (b) chromatin granulation, (c) picnotic nucleus, (d) laggardchromosome, (e) chromatin granulation and nucleus degeneration and (f) chromosome stickiness (Scale bars = 10 μm) |
The Kind of Abnormalities Caused by Fusilade Doses
As a result of our studies a few abnormalities were observed in control group while many were observed in the groups treated with Fusilade. In interphase and prophase (1 and 1.5% Fusilade); two different sizes of nuclei was observed in one cell (Fig. 2a), instead of normal chromatin condensation chromatin granulation was observed (Fig. 2b), cell membrane and nucleus were degenerated (Fig. 2e), picnotic nuclei (Fig. 2c), laggard chromose (Fig. 2d), deformation in shape of nucleus (Fig. 2e) and chromosome stickiness were observed (Fig. 2f).
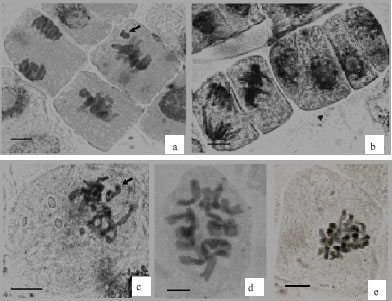 | |
| Fig. 3: | The abnormalities in metaphase cells in Lens culinaris root meristematic cells (a and c) laggard chromosome (pointed with arrow), (b) orientation fault in equatorial plate, (d) c-mitosis and (e) stickiness and swollen chromosomes (Scale bars = 10 μm) |
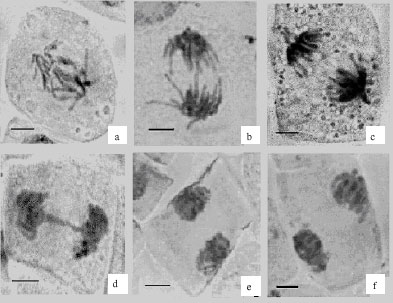 | |
| Fig. 4: | The abnormalities observed in anaphase and telophase cells of Lens culinaris root meristematic cells (a and b) bridge formation in anaphase, (c) orientation fault in equatorial plate, d. bridge formation in telophase, (e and f) chromosomes withdraw transversally in telophase (Scale bars = 10 μm) |
In metaphase (0.5, 1 and 1.5% Fusilade); laggard chromosomes (Fig. 3 a and c), orientation fault leading diagonal withdraw (Fig. 3b), chromosome stickiness and swollen chromosomes (Fig. 3e) and c-mitosis were observed (Fig. 3d).
In anaphase (1 and 1.5% Fusilade); chromosomal bridges (Fig. 4a and b), orientation fault in equatorial plate (Fig. 4c), chromosomal brigde formation (Fig. 4d) were observed and also chromosomes withdraw transversally in telophase (Fig. 4e and f). Chromosome bridges and laggard chromosomes were observed mostly in 1 and 1.5% Fusilade concentrations.
DISCUSSION
Fusilade kills annual and perennial grasses by inhibiting lipid synthesis (lipids are necessary components of cell membranes), particularly at the sites of active growth. In the environment, fluazifop-p-butyl is degraded primarily through microbial metabolism and hydrolysis (Randall and Tu, 2001). In our previous study cytological observations showed that the mitotic frequency in lentil root meristematic cells was decreased parallel to the increase in doses and all Fusilade doses applied decreased the activity of á-amylase enzyme in lentil seeds (Aksoy Dalgic et al., 2007).
In our experiments the negative side effects of Fusilade on lentil root tip cells during mitosis were investigated. While we observed normal mitotic divisions in the control group, different kinds of abnormalities were seen in all of the Fusilade doses varying to phases of mitosis. The studies about the effects of different pesticides found out some of the genotoxic effects of pesticides on plants (Sinha, 1989; Basic et al., 1991; Aktac et al., 1994; Tartar et al., 2006). In our study chromosomal bridges, lagging chromosomes, stickiness and change in the plane of the cell division were observed. Similar abnormalities were recorded with treatment conducted with the herbicide flurochloridone on Allium cepa root tips (Yuzbasioglu et al., 2003) and with the herbicide Logran on root tip cells of Triticum aestivum L. and Hordeum vulgare L. (Kaymak and Muranli, 2006). The types of chromosomal abnormalities observed after treatment with linuron in mitosis of Helianthus annuus L. included: fragments, disturbed metaphase, c-mitosis, lagging chromosome and chromatid bridge (Inceer et al., 2004).
Especially we observed the changes in the plane of cell division in all phases of mitosis in Lens culinaris root tips treated with higher doses of Fusilade. Disruption of cell division is usually caused by a primary effect on the mitotic apparatus (Hess, 1999). This apparatus organizes and separates the chromosomes during mitosis, determines the plane of cell division and orients cellular microfibril deposition in growing plant cells (Appleby and Valverde, 1989). A number of herbicides have been examined on Arabidopsis thaliana roots, with an emphasis on cortical microtubules and increased root tip swelling (Baskin et al., 2004). The herbicide aspendimethalin affect cell division and expansion processes in plant growth by inhibiting microtubule formation. This includes preprophase, spindle and phragmoplast microtubules in mitosis and cortical microtubules in controlling growth direction and cell wall formation (Nick, 2000; Wasteneys, 2004). Many commonly used synthetic herbicides, such as dinitroanilines, carbamates and dithiopyr produce similar effects on root meristems. Elmore and Bayer (1992) reported that some herbicides produce abnormal mitotic figures resulting in disrupted cell division on perennial ryegrass root meristems.
Genetic changes induced by pesticides, their metabolites and residues are expressed by various endpoints, which include; structural changes in chromosomes and chromatids, called chromosomal aberrations (breaks, deletions, inversions, gaps, translocations, rings) and other disturbances (stickiness, clumping, erosion). Disturbed phases may be due to disturbance in the function of the mechanism of chromosomes movement and the orientation of these chromosomes at the equatorial plate (Shehata, 2000).
We observed that c-metaphase where chromosomes appear scattered in the cytoplasm was found in 0.5, 1 and 1.5% Fusilade treatments. In this type of aberration, the metaphase chromosomes are shorter, thick and show no equator orientation. According to us, the reason of c-mitosis and the change in the plate of the cell division is the effect of Fusilade on the polimerization of microtubules. Such type of anomalies is an indication of the action of the herbicide on the inhibition of spindle fiber formation by their action on microtubules, which play the major role in the formation of spindle fibers (Shehata, 2000).
The most common type of aberrations in the present study was the irregular and transversally orientation of chromosomes in the equatorial plate and it was observed in almost all of the different phases of mitosis. These aberrations could also be attributed to the failure of the spindle apparatus to organize and function in a normal way rather than inhibition of these spindle fibers and this may lead to irregular orientation of chromosomes (Grant, 1978; Mansour, 1984). Induction of chromosomal bridges at anaphase and telophase was also observed after treatment with higher doses of Fusilade. These bridges may result from chromosome stickiness (Badr, 1983). Due to such stickiness the separation of the daughter chromosomes becomes incomplete even in the presence of spindle fibers and thus they remain connected by chromosome bridges (Kabarity et al., 1974), similar results were obtained by several authors (Badr, 1983; Soliman and Ghoenam, 2004).
Plants are the main recipients of pesticides, regardless of whether they themselves represent the target organism (e.g., weed) or whether the targets are pests, pathogenic fungi, etc. They are exposed to pesticides from direct application, through the uptake from soil and water and from atmospheric drift. Pesticides tend to be very reactive, mostly electrophilic, compounds that can react with various nucleophilic centers of cellular biomolecules, including DNA (Crosby, 1982), or form even more reactive electrophilic products that either modify cellular components or are metabolized to more or less stable products. It may be concluded from the present results that Fusilade can cause genotoxicity to the root meristematic cells of Lens culinaris.
REFERENCES
- Badr, A., 1983. Mitodepressive and chromotoxic activities of two herbicides in Allium cepa. Cytologia, 48: 451-457.
CrossRefDirect Link - Baskin, T.I., G.T.S. Beemster, J.E. Judy-Marsh and F. Marga, 2004. Disorganization of cortical microtubules stimulates tangential expansion and reduces the uniformity of cellulose microfibril alignment among cells in the root of Arabidopsis. Plant Physiol., 135: 2279-2290.
CrossRefPubMedDirect Link - Cione, N.K., P.E. Padgett and E.B. Allen, 2002. Restoration of a native shrubland impacted by exotic grasses, frequent fire and nitrogen deposition in southern California. Restor. Ecol., 10: 376-384.
CrossRefDirect Link - Dalgie, O.D.F., 2005. The effect of fungicide benomyl (benlate) on growth and mitosis in onion (Allium cepa L.) root apical meristem. Acta Biol. Hung., 56: 119-128.
CrossRefPubMedDirect Link - Erlingson, M., 1988. Fusilade-a strategy for long-term control of couch (Elymus repens). Weeds Weed Control, 29: 158-165.
Direct Link - Gorlach-Lira, K., O. Stefaniak, W. Slizak and I. Owedyk, 1997. The response of forest soil microflora to the herbicide formulations Fusilade and Roundup. Microbiol. Res., 152: 319-329.
CrossRefDirect Link - Kaymak, F. and F.D.G. Muranli, 2006. The genotoxic effects of Logran on Hordeum vulgare L. and Triticum aestivum L. Acta Biol. Hung., 57: 71-80.
CrossRefPubMedDirect Link - Ouzounidou, G., M. Moustakas and E.P. Eleftheriou, 1997. Physiological and ultrastructural effects of cadmium on wheat (Triticum aestivum L.) leaves. Arch. Environ. Contam. Toxicol., 32: 154-160.
CrossRef - Soliman, M.I. and G.T. Ghoneam, 2004. The mutagenic potentialities of some herbicides using Vicia faba as a biological system. Biotechnology, 3: 140-154.
CrossRefDirect Link - Tartar, G., F. Kaymak and F.D.G. Muranli, 2006. Genotoxic effects of avenoxan on Allium cepa L. and Allium sativum L. Caryologia, 59: 241-247.
CrossRefDirect Link - Wasteneys, G.O., 2004. Progress in understanding the role of microtubules in plant cells. Curr. Opin. Plant Biol., 7: 651-660.
CrossRefPubMedDirect Link - Yuzbasioglu, D., F. Unal, C. Sancak and R. Kasap, 2003. Cytological effects of the herbicide racer flurochloridone on Allium cepa. Caryologia, 56: 97-105.
Direct Link - Aksoy, O., F. Dane, F. Sanal and T. Aktac, 2007. The effects of fusilade (Fluazifop-p-butyl) on germination, mitotic frequency and α-amylase activity of lentil (Lens culinaris Medik.) seeds. Acta Physiol. Plant., 29: 115-120.
CrossRefDirect Link