
S.M. Manush
Division of Fish Nutrition and Biochemistry, Central Institute of Fisheries Education, Fisheries University Road, Versova, Mumbai-400 061, India
A.K. Pal
Division of Fish Nutrition and Biochemistry, Central Institute of Fisheries Education, Fisheries University Road, Versova, Mumbai-400 061, India
T. Das
Division of Fish Nutrition and Biochemistry, Central Institute of Fisheries Education, Fisheries University Road, Versova, Mumbai-400 061, India
N. Chatterjee
Division of Fish Nutrition and Biochemistry, Central Institute of Fisheries Education, Fisheries University Road, Versova, Mumbai-400 061, India
K. Sarma
Division of Fish Nutrition and Biochemistry, Central Institute of Fisheries Education, Fisheries University Road, Versova, Mumbai-400 061, India
S.C. Mukherjee
Division of Fish Nutrition and Biochemistry, Central Institute of Fisheries Education, Fisheries University Road, Versova, Mumbai-400 061, India
Asian Journal of Cell Biology
Year: 2007 | Volume: 2 | Issue: 1 | Page No.: 1-10







ABSTRACT
Cellular alterations of gills of freshwater prawn, Macrobrachium rosenbergii to three temperatures (25, 30 and 35°C) was assessed by means of Transmission Electron Microscopy (TEM). Acclimation of adult prawns (Macrobrachium rosenbergii) was carried out at one degree/day over and below ambient water temperature (30°C) to reach test temperatures (25, 30 and 35°C) and maintained for 30 days. Acclimated prawns were subjected to a constant rate of increase or decrease (0.3°C/min) until critical thermal maxima (CTMax) and critical thermal minima (CTMin) were reached. Dissected out gills were processed for TEM, both at the end of acclimation period and at critical thermal limits. Results indicated that at 25 and 30°C, gill tissues appeared normal. However, severe destruction of gill lamellae was observed at 35°C. At 25 and 30°C, primary lamellae showed active with euchromatin appeared dispersed in the matrix. At 35°C, severe degeneration of mitochondriae with loss of cristae, irregular nuclear membrane and margination of chromatin were evident. Gill tissues at CTMax showed severely damaged lamellae, increased number of lysosomes and karyorhexis, destruction of mitochondria and nuclear membrane, were observed. At CTMin, gill tissues exhibited little visible cellular damage as compared to CTMax. Our prima-facie report indicates M. rosenbergii demonstrates acclimatory responses to temperature. However, cellular integrity may be affected due to long-term exposure to higher temperatures (35°C).
PDF Abstract XML References Citation
How to cite this article
S.M. Manush, A.K. Pal, T. Das, N. Chatterjee, K. Sarma and S.C. Mukherjee, 2007. Ultrastructural Alterations in the Gills of Macrobrachium rosenebergii Acclimated to Three Temperatures. Asian Journal of Cell Biology, 2: 1-10.
DOI: 10.3923/ajcb.2007.1.10
URL: https://scialert.net/abstract/?doi=ajcb.2007.1.10
DOI: 10.3923/ajcb.2007.1.10
URL: https://scialert.net/abstract/?doi=ajcb.2007.1.10
INTRODUCTION
Freshwater prawn farming has become a significant and valuable sector of global aquaculture, contributing 0.3 million MT of all Macrobrachium species raised through aquaculture and is being increased with an annual expansion rate of 48% between 1999 and 2001 (New, 2003). The world production of M. rosenbergii increased from 26, 588 MT in 1991 to 118, 501 MT in 2000 (FAO, 2002). In India, the total annual production of freshwater prawns (M. rosenbergii and M. malcomsonii) increased from 7,140 MT to 30,450 MT (4.3 fold increase) from 1999-2000 and a corresponding increase in the area of prawn culture from 12,022 to 34,630 ha (2.9 fold increase) from 2002-2003 (Marine Products Export Development Authority, 2003), which clearly indicates the importance of this species in freshwater aquaculture. However, prawns encounter different stressors during culture condition, which limits their output from aquaculture (Manush et al., 2005).
Temperature virtually affects all levels of biological organization, membrane permeability, cellular, tissue and organ function and integration in the whole organism. Every ecothermic animal can tolerate a range of temperature, known as thermal tolerance, which is dependent on acclimation temperatures (Manush et al., 2004), species (Das et al., 2004), life stages (Chatterjee et al., 2004, Das et al., 2005). Rising temperature up to a certain limit favors aquaculture by reducing the time required to produce marketable sized animals and producing more generations per year. Beyond a threshold limit, temperature may induce stress in the organism, results in irreversible cellular alterations (Egginnton and Sidell, 1989) and becomes lethal.
The United States National Research Council proposed that the global mean temperature may increase by 1.5-4.5°C in the next half century (Beitinger et al., 2000). The potential effect of global warming makes continuous effort to define thermal adaptation of shellfishes and their consequences on health, an important area of research. The increasingly obvious problem of global warming has given greater urgency to understand biological responses to temperature, particularly when ectothermal organisms have limited independence from changes in environmental temperature. Surface water temperatures are likely to fluctuate up to 40°C based on latitude, season, altitude, time of day, depth of water and other factors (Munro and Roberts, 2001) and is more frequent in tropical shallow freshwater bodies than in sea. It is essential to know the effect of thermal extremes (maximum and minimum) on histo-architecture of vital organs in aquatic organisms. Of different organs, gills are vital structures, since they are the main sites for gaseous exchange (Hughes, 1982) and partially responsible for osmoregulation (Verbost et al., 1994), acid-base balance (Lin and Randall, 1991; Goss et al., 1992) and excretion of nitrogenous compounds (Evans and Cameron, 1986; Sayer and Davenport, 1987) in fishes and shellfishes. Therefore, gills can be used as model organ for assessing the environmental impact on the test animals. Against this background, we investigated cellular alteration of M. rosenbergii gills in response to thermal acclimation at three different temperatures (25, 30 and 35°C) and exposure to thermal extremes (critical thermal limits).
MATERIALS AND METHODS
Experimental Animals
Adult prawns (M. rosenbergii) (mean weight±SE = 38±3.1 g) were brought in aerated open containers from Bhivandi Farm, Mumbai to Wet Laboratory, Central Institute of Fisheries Education, Mumbai and were held for 15 days at ambient water temperature (30°C). During this period, prawns were fed with supplementary feed before thermal tolerance studies.
Protocol for Acclimation
Acclimation of prawns (6/aquarium) to test temperatures was carried out in a thermostatic aquarium (52 L water capacity, sensitivity±0.2°C). Acclimation was carried out at one degree/day change from ambient water temperature (30°C) to reach test temperatures (25, 30 and 35°C) and maintained for a period of 30 days prior to the critical thermal methodology (CTM) experiments. Pretrial acclimation periods and experimental acclimation temperatures suggested for conducting experiments in shellfishes remains a debatable topic among physiologists across the globe. As the thermal acclimation period of adult stages of Macrobrachium rosenbergii were unknown, other life stages were considered. M. rosenbergii post larvae were completely acclimated after four weeks of acclimation (Herrera et al., 1998). Therefore we assumed our experimental animals were completely acclimated prior to CTM tests.
Critical Thermal Methodology (CTM)
A total of 36 prawns were distributed in three groups and critical thermal maxima (CTMax) and critical thermal minima (CTMin) were determined (Beitinger et al., 2000). Animals acclimated to three temperatures were subjected to a constant rate of increase or decrease (0.3°C min-1) until Loss of Equilibrium (LOE) was reached. LOE was designated as critical thermal maxima (CTMax) and critical thermal minima (CTMin), respectively (Paladino et al., 1980; Beitinger et al., 2000). This technique has been critically evaluated by numerous workers (Hutchinson, 1976; Reynolds and Casterlin, 1979) and is well established as a powerful tool for studying the physiology of stress and adaptation in fishes (Paladino et al., 1980; Beitinger and McCauley, 1990). Dissolved oxygen concentration was maintained at 5.5±0.5 mg L-1 throughout the CTM tests by continuous aeration using a 2HP centralized air blower.
Ultra-structural Analysis by Electron Microscopy
Transmission Electron Microscopic (TEM) study was carried out using a transmission electron microscope (Model JEM-1010, Japan) at Jaslok Hospital and Research Centre, Pedder Road, Mumbai as per the protocol described by Ghadially (1986). Prior to dissection for ultra structural studies, prawns were anesthetized using CIFECALM (200 μL L-1). CIFECALM is a herbal anaesthetic formulation containing natural alcoholic extracts of Eugenia caryophyllata and Mentha arvensis. (developed by Central Institute of Fisheries Education, Mumbai). Gill samples of prawns (after acclimation at different temperatures and at CTM endpoints) were fixed in 3% glutaraldehyde using 1 M sodium cacodylate buffer for 8 h at 4°C. The samples were then washed with sodium-cacodylate buffer (four times at 5 min intervals) and were subsequently fixed by osmication with a solution of 1% osmium tetroxide (in D/W) and 1 M sodium cacodylate in the ratio of 1:1 (v/v) for 1-2 h at 4°C. The samples were washed with 1 M sodium cacodylate buffer (10 min) followed by dehydration schedule with ethyl alcohol/acetone in the increasing order of concentration. Semi-thin sections were cut and stained with 20% toluidine blue and were observed under 40X. Suitable areas were selected and marked for utlra-thin sectioning. The ultra-thin sections were stained with lead citrate and uranyl acetate and subsequently scanned under TEM for observation and electron micrography.
RESULTS
Ultra-structural cellular architecture of gill tissues in M. rosenbergii acclimated to three temperatures (25, 30 and 35°C) and at CTM end points (CTMax and CTMin) are presented as Fig. 1-4.
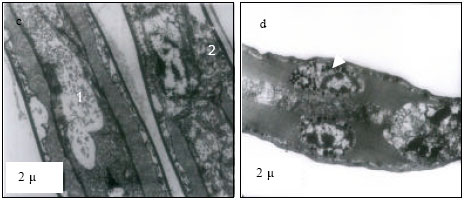
At 25 and 30°C, primary lamellae appeared to have normal cells and organelles (Fig. 1a and b). However, at 35°C, karyolysis in the cell of lamella and margination of chromatin granules in the cell were evident (Fig. 1c). At critical thermal limits, karyorrhexis was observed in the nucleus and dense lysosomes were appeared as dark circular inclusions in the cytoplasm at CTMax (Fig. 1d). At CTMin, mitotic phase of a cell appeared with dumb-bell shaped nucleus, heterochromatin appeared highly condensed moving to periphery (Fig. 1e). At 25°C, nucleus appeared normal with active aggregates of chromatin granules (Fig. 2f), with distinct envelope and euchromatin dispersed in the matrix with darkly stained nucleolus (Fig. 2g) at 30°C. Margination of chromatin granules in the nucleus (Fig. 2h) was evident at 35°C. Gills at critical thermal limits demonstrated severe damage. At CTMax, darkly stained chromatin materials in condensed form, nucleus in stage of fragmentation and cytoplasmic organelles in a cloudy form were visible (Fig. 2i) and mitochondrial swelling with loss of cristae were evident. In a CTMin cell, margination of chromatin granules, condensation of the nucleus and a total loss of mitochondria were evident (Fig. 2j).
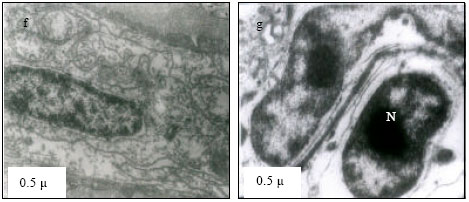
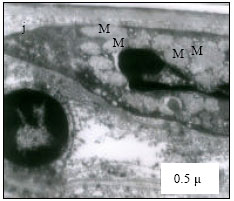
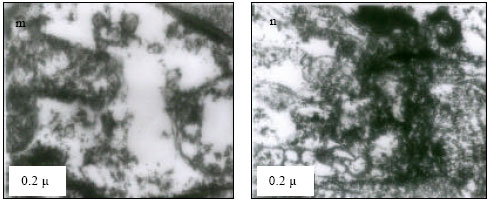
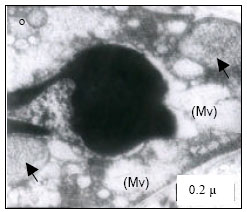
Mitochondria (M) appeared normal with few cristae (Fig. 3k) at 25°C. However, at 30°C mitochondria appeared to have dense cristae (Fig. 3l). At 35°C, mitochondrial degeneration devoid of cristae and severe destruction of the membrane were evident with vacuolated area in the cytoplasm (Fig. 3m). Severe destruction of mitochondria was visible at CTMax (Fig. 3n) and mitochondrial swelling with degenerating cristae and vacuolated areas due to marked degeneration of the mitochondriae were evident (at CTMin) (Fig. 3o).
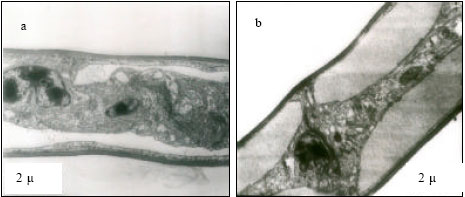 | |
| Fig. 1: | Transmission Electron micrograph of gills in M. rosenbergii acclimated at three temperatures (25, 30 and 35°C). Note the section of primary lamella with active cells and organelles (a and b) (X2.5 K, bar = 2 µ) at 25 and 30°C respectively, karyolysis in the cell of lamella (1) and margination of chromatin granules in the cell (2) (c) (X2.5 K, bar = 2 µ) at 35°C, Nucleus with karyorhexis, dense lysosomes (arrow) appear as dark circular nclusions (d) (X2.5 K, bar = 2 µ) at critical thermal maxima (CTMax), mitotic phase of a cell with dumb-bell shaped nucleus. Heterochromatin highly condensed moving to periphery (e) (X2.5 K, bar = 2 µ) at critical thermal minima (CTMin), nucleus appears normal with active aggregates of chromatin granules(f) (X10K, bar = 0.5 µ) at 25°C, nuclei appear normal with distinct envelope (arrow) euchromatin dispersed in the matrix with darkly stained nucleolus |
 | |
| Fig. 2: | Transmission Electron micrograph of gills in M. rosenbergii acclimated at three temperatures (25, 30 and 35°C) Note the section of primary lamella with active cells and organelles (g) (X10K, bar = 0.5 µ) at 30°C, margination of chromatin granules in the nucleus (h) (X10K, bar = 0.5 µ) at 35°C, darkly stained chromatin materials in condensed from, nucleus is in the stage of fragmentation and cytoplasmic organelles are in a cloudy form (i) (X10K, bar = 0.5 µ) at CTMax, Note mitochondrial swelling with loss of cristae (M). In another cell, margination of chromatin granules, condensation of the nucleus is evident with total loss of mitochondria (J) (X10K, bar = 0.5 µ) at CTMin, mitochondria (M) appeared normal with few cristae |
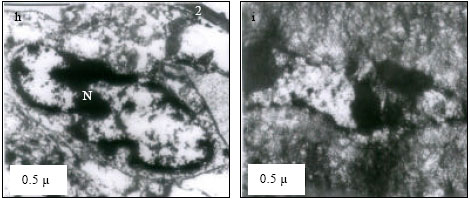
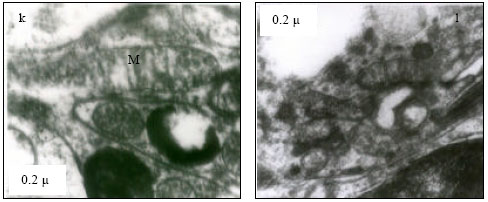 | |
| Fig. 3: | Transmission Electron micrograph of gills in M. rosenbergii acclimated at three temperatures (25, 30 and 35°C) Note the section of primary lamella with active cells and organelles (k) (X25K, bar = 0.2 µ) at 25°C, mitochondria (M) appeared normal with dense cristae (l) (X25K, bar = 0.2 µ) at 30°C, mitochondrial degeneration devoid of cristae and severe destruction of the membrane, with vacuolated area in the cytoplasm (m) (X25K, bar = 0.2 µ) at 35°C, severe destruction of mitochondriae (n) (X25K, bar = 0.2 µ) at CTMax, Mitochondrial swelling with degenerating cristae (arrow) and vacuolated area due to marked degeneration of the Mitochondrae (Mv) (o) (X25K, bar = 0.2 µ) at CTMin, intact nuclear membrane, with evenly distributed euchromatin materials |
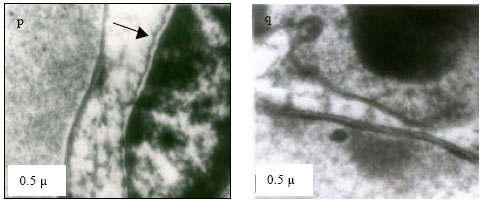 | |
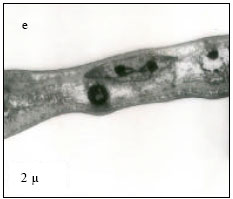
| Fig. 4: | Transmission Electron micrograph of gills in M. rosenbergii acclimated at three temperatures (25, 30 and 35°C) Note the section of primary lamella with active cells and organelles (p) (X30K, bar = 0.2 µ) at 25°C, cell membrane normal and chromatin highly condensed (q) (X30K, bar = 0.2 µ) at 30°C, Irregular nuclear membrane and margination of chromatin distinctly visible I (r) (X30K, bar = 0.2 µ) at 35°C, similar degeneration of nuclear membrane, margination of chromatin visible (s) (X30K, bar = 0.2 µ) at CTMax, Nuclear membrane intact with highly condensed chromatin (t) (X30K, bar = 0.2 µ) at CTMin |


Nuclear membrane was intact, with evenly distributed euchromatin materials (Fig. 4p) at 25°C. At 30°C, the cell membrane appeared normal. However, highly condensed chromatin was observed (Fig. 4q). Irregular nuclear membrane and margination of the chromatin was distinctly visible (Fig. 1r) at 35°C. At CTMax, similar degeneration of nuclear membrane and margination of the chromatin was visible (Fig. 4s) At CTMin, nuclear membrane was intact, however with highly condensed chromatin (Fig. 4t).
DISCUSSION
Biologists have been studying thermal tolerance of poikilotherms for more than a hundred years, considering its multifaceted significance (Heath, 1884; Hezel and Prosser, 1974; Monica et al., 1996; Beitinger et al., 2000; Fernando et al., 2000, 2002; Monica and Fernando, 2002). The increasingly obvious problem of global warming has given greater urgency to the understanding of biological responses to temperature, particularly in the ectothermal organisms that have limited independence from changes in environmental temperature. Our earlier investigations revealed that M. rosenbergii demonstrates thermal acclimation dependent adaptation (Manush et al., 2004). In the present study, we present the prima-facie report on cellular change, in response to thermal acclimation and due to thermal extremes.
Prawn gills were taken into consideration for present investigation as they were in direct contact with environment and therefore, gills could be good indicators of water quality. Conduction of heat between the environment and prawns is mediated through the gills; hence, gills are regarded as the heat exchanger. Therefore, gills are considered to be the most appropriate organ for indicating thermal pollution (Alazemi et al., 1996) and consequently in all the pollution studies in gill tissues of prawns should be invariably become an organ of paramount importance to assess the magnitude of damage. Prawns exposed to high temperatures may suffer respiratory stress and nervous disorders. Increasing water temperature exerts a dual effect on their oxygen demand-supply ratio. Warmer waters carry less dissolved oxygen, yet the oxygen uptake by aquatic organisms is increased due to their higher metabolic rate. One of our earlier investigations indicated that metabolic activity increased with increasing acclimation temperatures (Manush et al., 2004). Cellular structures observed from the present ultrastructural study indicated that 25 and 30°C are not detrimental to M. rosenbergii. At 35°C, destruction of gill lamellae was evident, as it may be due to osmotic imbalance. It is reported that higher water temperature affects the ability of fishes to maintain the osmotic balance by altering the lipids of gill cells, resulting in leakage of cells and reducing the efficiency of salt excretion and balance (Munro and Roberts, 2001).
A relation could be established with mitochondrial morphology and metabolic activity. Denser mitochondrial cristae was evident at 30°C over the cells acclimated at 25°C. This may be due to the increase in metabolic activity. Similar correlation of mitochondrial cristae with metabolic status is evident in high energy transforming flight muscle of the dragon fly (Ghadially, 1982). However, acclimation at 35°C imparted tissue damage in the present study. Irregularity of nuclear membrane is considered as an indication of increased metabolic activity, as nuclear irregularity increases the contact between nucleus and cytoplasm and nucleocytoplasmic exchanges during high metabolic activity, which was evident in prawns acclimated at 35°C. Degeneration of mitochondria was visible in the cells at 35°C. Margination of chromatin granules, pyknosis and karyolysis of cell are considered as different stages of cell death, which was evident in the present study at 35°C. Similar findings were reported in mouse liver incubated in aseptic conditions (Trump et al., 1963). At CTMax, margination of chromatin granules, karyorhexis of nucleus and dense lysosomes of cytoplasm were observed and are clear evidence of cell death. At CTMin, lesser tissue damage was evident in gill tissues of M. rosenbergii as compared to CTMax. Mitochondrial hypertrophy and hyperplasia must be a compensatory mechanism to derive energy for combating cold stress. Similar findings were reported in Labeo rohita in response to cold acclimation (Das, 2004).
Overall results indicate that cellular integrity is altered in the gills, in response to thermal acclimation and exposure to thermal extremes. From our results, it is evident that long-term exposure to increasing temperatures (35°C and above) due to global warming and climatic changes may cause respiratory stress and breakdown of compensatory general adaptive syndrome (GAS) and ultimately endanger M. rosenbergii in open fresh water bodies.
ACKNOWLEDGMENTS
The first author is thankful to Indian Council of Agricultural Research for providing financial assistance in the form of institutional fellowship during the research period.
REFERENCES
- Alazemi, B.M., J.W. Lewis and E.B. Andrews, 1996. Gill damage in the freshwater fish Gnathonemus petersii (family: Mormyridae) exposed to selected pollutants: An ultrastructural study. Environ. Technol., 17: 225-238.
CrossRef - Beitinger, T.L. and R.W. McCauley, 1990. Whole anmal physiological processes of the assessment of stress in fishes. J. Great Lakes Res., 16: 542-575.
Direct Link - Beitinger, T.L., W.A. Bennett and R.W. McCauley, 2000. Temperature tolerances of North American freshwater fishes exposed to dynamic changes in temperature. Environ. Biol. Fish., 58: 237-275.
Direct Link - Manush, S.M., A.K. Pal, N. Chatterjee, T. Das and S.C. Mukherjee, 2004. Thermal tolerance and oxygen consumption of Macrobrachium rosenbergii acclimated to three temperatures. J. Thermal Biol., 29: 15-19.
Direct Link - Das, T., A.K. Pal, S.K. Chakraborty, S.M. Manush, N. Chatterjee and S.C. Mukherjee, 2004. Thermal tolerance and oxygen consumption of Indian Major. J. Thermal Biol., 23: 157-163.
CrossRef - Das, T., A.K. Pal, S.K. Chakraborty, S.M. Manush, N.P. Sahu and S.C. Mukherjee, 2005. Thermal tolerance, growth and oxygen consumption of Labeo rohita fry (Hamilton, 1822) acclimated to four temperatures. J. Thermal Biol., 30: 378-383.
Direct Link - Egginton, S. and B.D. Sidell, 1989. Thermal acclimation induces adaptive changes in subcellular structure of fish skeletal muscle. Am. J. Physiol., 256: R1-R9.
CrossRefDirect Link - Fernando, D., A. Miguel, S. Elizabeth, A. Marisela and R. Denisse, 2000. Preferred temperature and critical thermal maxima of red abalone Haliotis rufescens. J. Thermal Biol., 25: 257-261.
CrossRef - Fernando, D., S. Elizabeth, Ana R. Denisse and R. Leticia, 2002. Behavioural thermoregulation and critical thermal limits of Macrobrachium acanthurus (Weigman), J. Thermal Biol., 27: 423-428.
CrossRef - Lin, H. and D. Randall, 1991. Evidence for the presence of an electrogenic proton pump on the trout gill epithelium. J. Exp. Biol., 161: 119-134.
Direct Link - Manush, S.M., A.K. Pal, N. Chatterjee, T. Das and S.C. Mukherjee, 2004. Thermal tolerance and oxygen consumption of Macrobrachium rosenbergii acclimated to three temperatures. J. Thermal Biol., 29: 15-19.
Direct Link - Manush, S.M., A.K. Pal, T. Das and S.C. Mukherjee, 2005. Dietary high protein and vitamin C mitigate stress due to chelate claw ablation in Macrobrachium rosenbergii males. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol., 142: 10-18.
CrossRefDirect Link - Monica, H.R., B.R. Fernando and D.H. Fernando, 1996. Critical thermal maximum of Macrobrachium tenellum. J. Thermal Biol., 21: 139-143.
Direct Link - Monica, H.R. and B.R. Fernando, 2002. Temperature tolerance polygon of Poecilia sphenops (Pisces: Poeciliidae). J. Thermal Biol., 27: 1-5.
CrossRef - Sayer, M.D.J. and J. Davenport, 1987. The relative importance of the gills to ammonia and urea excretion in five seawater and one freshwater teleost species. J. Fish. Biol., 31: 561-570.
CrossRefDirect Link