
Fatma A. Abdel Razek
National Institute of Oceanography and Fisheries, Invertebrate Laboratory, Alexandria, Egypt
LiveDNA: 20.3098
Marwa I. Farghaly
Department of Zoology, Faculty of Science, Alexandria University, Alexandria, Egypt
Jehan Sorour
Department of Zoology, Faculty of Science, Alexandria University, Alexandria, Egypt
Azza Attia
Department of Zoology, Faculty of Science, Alexandria University, Alexandria, Egypt
Asian Journal of Biological Sciences
Year: 2019 | Volume: 12 | Issue: 4 | Page No.: 626-636







ABSTRACT
Background and Objective: Despite its abundance and commercial importance, little is known about the biology of the blue swimmer crab, Portunus pelagicus (Linnaeus, 1758), of the Egyptian waters. The study aimed to characterize the maturation stages of male and female P. pelagicus through external morphological and histological examination during gonad development, with determination of gonadosomatic index (GSI), size at first sexual maturity and spawning population from Alexandria coastal waters of Egypt. Materials and Methods: The current study investigated the sexual maturity and other reproductive biology characteristics of P. pelagicus from the Egyptian Mediterranean coast off Alexandria during the period from March, 2015-February, 2016 and the data were statistically analyzed, represented in mean and standard deviation. Results: The length-weight relationships of males showed nearly an isometric pattern of growth, whereas, in females they revealed a marked departure from isometric growth. Female gonadal development stages were morphologically and histologically investigated, they were comprised of five stages, while three development stages were recorded for male. The size at which P. pelagicus reached sexual maturity was at 107.6 and 96.1 mm carapace width (CW) for males and females, respectively. Ovigerous female showed a prolonged occurrence during the warm months, from March-October. Conclusion: According to gonad maturation, GSI and the occurrence of ripe and ovigerous females, two peaks of spawning activity were detected, in April-May and in August-September. Hence, it is reasonable to say that ovigerous females may spawn more than one time throughout the year.
PDF Abstract XML References Citation
Received: December 22, 2018;
Accepted: February 04, 2019;
Published: July 29, 2019
How to cite this article
Fatma A. Abdel Razek, Marwa I. Farghaly, Jehan Sorour and Azza Attia, 2019. Population Characteristics, Maturation and Spawning of the Blue Swimmer Crab Portunus pelagicus in Eastern Mediterranean Sea, Egypt. Asian Journal of Biological Sciences, 12: 626-636.
URL: https://scialert.net/abstract/?doi=ajbs.2019.626.636
URL: https://scialert.net/abstract/?doi=ajbs.2019.626.636
INTRODUCTION
Recently, crabs are becoming one of the most popular food item, which commands high prices in the international and national markets. Portunus pelagicus (Linnaeus, 1758) is considered the major crab species supporting crab fisheries in the Egyptian Red and Mediterranean seas, as well as in the Bardawil Lagoon. Moreover, it has a considerable demand as it is an admired seafood delicacy. Egyptian crab fisheries production during the year 2016 had reached 5.17% (2790 of 53964 t) of the total fisheries from the Egyptian Mediterranean sector, this percent is represented mainly by P. pelagicus1.
Much work has been carried out on different aspects of the reproductive biology of P. pelagicus throughout the world such as in Australia2-7, India8-10 and Indonesia11-13. Few and relatively not recent data are available for P. pelagicus reproductive studies in Egyptian waters, in spite of its growing publicity as an economic and commercial seafood item14-16. The fine structure of the sperm and spermatophores of P. pelagicus were described by El-Sherief17,18. While the stages of the female P. pelagicus reproductive cycle were investigated by Bawab and El-Sherief19, depending on the spermatheca anatomical changes.
The information concerning its reproductive biology offers a valuable and unique tool for furthering our understanding of its developmental processes and to determine appropriate brood stock options for larval rearing of this species20. Additionally, fecundity, embryonic and gonad development, as well as spawning season are essential aspects of the reproductive biology of crab20 that enable us to understand the dynamics of the species population. Therefore, the aim of this study is to characterize the maturation stages of male and female P. pelagicus through external morphological and histological examination during gonad development, with determination of gonadosomatic index (GSI), size at first sexual maturity and spawning population from Alexandria coastal waters of Egypt. This fundamental information will inform the gaps which need to be addressed with regard to the reproductive biology of P. pelagicus in Egypt and could be used to support the implementation of policy for blue swimming crab’s sustainable fishery in the Egyptian waters.
MATERIALS AND METHODS
Sample collection: Males, ovigerous and non-ovigerous females of Portunus pelagicus were monthly collected from the fishing centers of the eastern Harbor of the Egyptian Mediterranean Coast of Alexandria city. It lies between longitudes 29°53'-29°54' 40" E and latitudes 31°12'-31°13'N” during the period from March, 2015 to February, 2016.
Data analysis
Sex determination: Crab samples were brought to the invertebrate laboratory of the National Institute of Oceanography and Fisheries (NIOF), Alexandria, Egypt. Males of P. pelagicus have an inverted ‘T’ shaped abdomen. Three categories of females were defined: Triangular shaped abdomen (V-shape) for immature female crabs, globular shaped abdomen (U-shape) for adult “non-ovigerous females” and the third category is the ovigerous females (egg-bearing females) as described by Josileen21.
Morphological measurements: Morphometric measurements of P. pelagicus were assessed including: Body weight (BW), Carapace width (CW), Carapace length (CL), Gonad weight. Vernier calipers with 0.05 mm accuracy were used for the length measurements, while the weights of the crab samples and their gonads were measured using a digital balance to the nearest (0.01 g). Linear and power regression analysis were used to investigate the relationship between (CW), (CL) and (BW).
Gonad examination: Monthly samples of male and female P. pelagicus were dissected in order to morphologically determine their gonadal development stages according to the shape and the color of the gonads. Also, the maturation stages of fresh gonads were determined before fixation. Small portions of the anterior, middle and posterior gonad lobes were fixed in Davidson solution for 24 h. Then, the gonad was transferred into the 70% alcohol solution before proceeding with tissue processing. The tissue was dehydrated in ascending ethanol concentrations from 70-100%, transferred to cedar oil and embedded in paraffin. The tissue was sectioned to 5 μm using a rotary microtome, then the sections were mounted on clean glass slides, then stained with hematoxylin-eosin. Thereafter, stained sections were examined with Olympus CX31 light microscope.
Determination of size at first sexual maturity: The size at which P. pelagicus was considered sexually matured was investigated by using linear regression by setting a relationship between the ratio of matured gonad of males and non-ovigerous females for each sequential 10 mm CW, to determine the size at which 50% of the crabs would have become mature according to Hartnoll22.
Determination of gonadosomatic index (GSI): Monthly GSI was calculated according to the following equation:
![]()
Data are represented in mean and standard deviation, one-way ANOVA analysis (p<0.005) was used to test the differences of GSI between males and non-ovigerous females with time.
RESULTS
Population characteristics: A total of 445 samples of Portunus pelagicus were collected during the study period from the eastern Harbor fishery centers of the Egyptian Mediterranean Coast off Alexandria city. The crab sample was composed of 132 males, 184 non-ovigerous females and 129 ovigerous females. The weight and length characteristics of P. pelagicus are represented in Table 1.
Width/length-weight relationships: Linear and power regression analysis were used to investigate the relationship between CW, CL and BW. The values and constants of regression equations are given in Table 1.
The results of regression analysis showed that the increase in carapace width is always followed by an increase in the carapace length and body weight, so CW is positively correlated with CL and BW. In addition, the exponential values (b) of the length-weight relationships of males followed the cube law (CW-BW, b = 3.0970, CL-BW, b = 3.0300) indicating thereby nearly an isometric pattern of growth, whereas, the corresponding values of females (CW-BW, b = 2.9124, CL-BW, b = 2.8293) suggested marked departure from isometric pattern of growth.
Macroscopic and histological description of gonad development
Gonadal development stages of female P. pelagicus: The present observations showed that female gonadal development of the blue swimmer crab was classified into five stages: (I) Immature, (II) Early maturing, (III) Maturing, (IV) Ripe and (V) spent.
Macroscopic description:
| • | Stage I "Immature": Ovary was small, flattened and thread-like smooth structure without obvious lobulation, transparent and it was difficult to recognize from the digestive cecum with a mean GSI of 0.76±0.48 |
| • | Stage II "Early maturing": Ovary was light yellow, the anterior part slightly displaced the hepatopancreas, the middle covered the heart and the posterior lobes were convoluted with a mean GSI of 1.42±1.42 |
| • | Stage III "Maturing": Ovary was large, yellow and nodulated. Hepatopancreas was displaced by the growing of anterior lobes, the gastric and intestinal cavities were found to be covered by central and posterior lobes with a mean GSI of 3.14±1.89 |
| • | Stage IV "Ripe": Ovary was very large, dark orange with more nodulations. It occupied nearly the whole space of the carapace with a mean GSI of 7.04±3.17 |
| • | Stage V "Spent": Ovary was small, off-white have nearly the same appearance of the ovary at immature stage, but found only in crabs which have U-shaped abdomen and not in immature female that have V-shaped abdomen, with a mean GSI of 0.90±0.7 |
| Table 1: | Population measurements of male, non-ovigerous and ovigerous females of Portunus pelagicus in the eastern Harbor, Alexandria and population regression analysis of male, female of Portunus pelagicus |
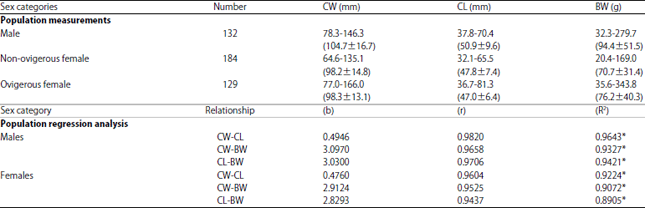 | |
Numbers in parentheses refer to the average±SD. CW: Carapace width, CL: Carapace length, BW: Body weight, b: Exponential value, r: Correlation coefficient, R2: Coefficient of determination, *: p<0.005 | |
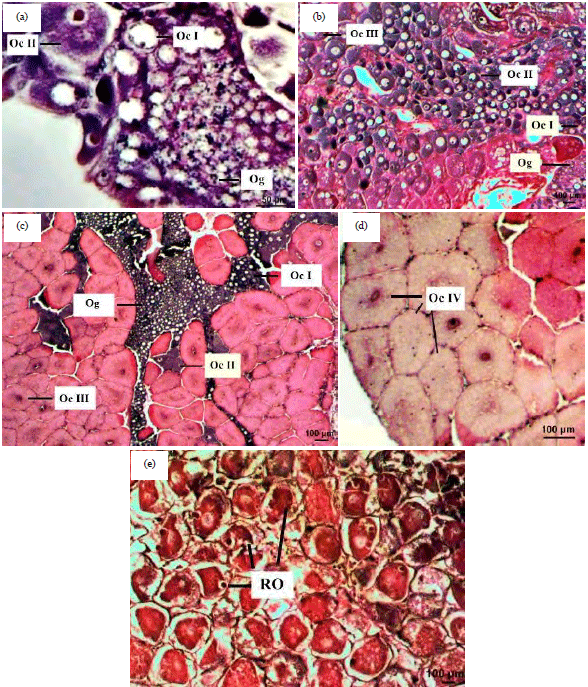 | |
| Fig. 1(a-e): | Photomicrographs of light microscope showing the ovary in Portunus pelagicus Stage (I, II, III, IV and V), (a) Immature stage, (b) Early maturing stage, (c) Maturing stage, (d) Ripe stage and (e) Spent stage. Oc I: Oocyte I, Oc II: Oocyte II, Oc III: Oocyte III, Oc IV: Oocyte IV, Og: Oogonia, RO: Resorbing oocyte |
Histological description: It is worth mentioning that the general ovarian structure and histology of P. pelagicus was similar to those of other brachyurans. During these five development stages, germ cells can be also divided into five growth development stages based on cellular diameters, shape and histological staining appearance. They were oogonia (Og) and four sequential types of oocytes (from Oc I to Oc IV). Histological description for gonadal development of female P. pelagicus is presented in Fig. 1a-e.
| • | Stage I"Immature": Ovary was composed of three various types of cells. Oogonial cells, oocyte I and II. The oogonial cells had each a spherical or a little oval shape, It had a mean cell diameter of 11 μm. Oocyte I (Previtellogenic oocyte, Oc I) had a spherical or a bit ovoid shape. It had a mean cell diameter of 39 μm. Its cytoplasm appeared with highly distinct blue color, while Oocytes II (Previtellogenic oocyte, Oc II) had an oval shape with a mean cell size of 57 μm. Its cytoplasm was blue with eosinophilic yolk granules (Fig. 1a) |
| • | Stage II "Early maturing": The ovaries became more enlarged. The ovaries are characterized by the appearance of oocyte III besides the other oogonial and oocyte I and II cells. Oocytes III (Early vitellogenic oocyte, Oc III) had an oval shape with a mean cell size of 92 μm. The cytoplasm was partially eosinophilic “appears pink in comparison to OcII” (Fig. 1b) |
| • | Stage III "Maturing": The ovaries at maturing stage contain Og, Oc I, Oc II and Oc III predominantly. Oc III was localized in the follicle periphery, where they develop to form Oc IV (Fig. 1c) |
| • | Stage IV "Ripe": Fully mature oocytes (Oc IV) were dominantly present. Each oocyte IV (Late vitellogenic oocyte; Oc IV) had a spherical or a little ovoid or hexagonal shape with a mean cell diameter of 156 μm, cytoplasm was extremely eosinophilic and granular because of the much presence of yolk granules (Fig. 1d) |
| • | Stage V "Spent": Blue swimmer crabs have accomplished breeding by reaching this stage, almost all the fully mature eggs had been extruded outside the ovary. Visible degeneration of the oocytes occurs and the ovaries suffer from a comprehensive breakdown of connective tissue, then after spawning, oogenesis is reinitiated immediately (Fig. 1e) |
Gonadal development stages of male P. pelagicus: The progress of the male maturation was assorted to three stages according to the external morphology of testes and vas deferens: Immature (I), maturing (II), mature (III).
Macroscopic description:
| • | Stage I "Immature": Testis and vas deferens were not clearly differentiated. Testis was small on either side of stomach; vas deferens was thin, with straight tubes and a mean GSI = 0.48±0.27 |
| • | Stage II "Maturing": Testis and vas deferens were well differentiated. Testis was large with coiled tube extending laterally and posteriorly in the stomach. Vas deferens was a white coiled mass about 0.5-1.0 mm thickness, spreading to both sides of the heart, with a mean GSI = 0.76±0.51 |
| • | Stage III "Mature": Testis showed additional enlargement. Vas deferens was much more swollen off-white mass extending to fill most body cavity and of 2-3 mm thickness with a mean GSI = 0.93±0.35 |
Histological description: It was interesting to note that testis showed almost no histological difference in the development stages. The histological differences were shown in the vas deferens structure. Descriptions of each gonadal development stage are presented in Fig. 2a-g.
| • | Stage I "Immature": Anterior vas deferens (AVD), median vas deferens (MVD) and posterior vas deferens (PVD) were devoid from spermatophores (Fig. 2a, b) |
| • | Stage II "Maturing": Spermatophores were found in AVD, but there were no spermatophores found in MVD and PVD (Fig. 2c, d) |
| • | Stage III "Maturing": Spermatophores were found in AVD and MVD. The PVD may or may not contain spermatophores (Fig. 2e-g) |
Size at 1st sexual maturity: The smallest recorded matured male and female crab during the period of study were at 88.6 and 72.5 mm (CW), respectively. On the other hand, by using the regression equation method, the calculated minimum biological sizes for mature male and female were found at 107.6 and 96.1 mm (CW), respectively (Fig. 3).
Gonadosomatic index (GSI): The mean values of monthly GSI of all males and females of P. pelagicus differ significantly (p<0.05) in all the study months (Fig. 4).
Distribution of different maturity stages and spawning activity
According to months: In the present study, for non-ovigerous females, the immature individuals were observed throughout the year with high percentages in August (56%) as shown in Fig. 5a. On the other hand, ripe stage occurred throughout the year except for the period from October -December. The high percentages of ripe individuals were in April and May, indicating that ripe females were expected to spawn with the starting of the warm months. While the highest percentage of occurrence of the spent individuals “which had accomplished breeding” was in October and November.
According to size: Ripe individuals of non-ovigerous females in the studied population started to appear from the size of 65 up to 135 mm (CW). On the other hand, the presence of immature females was represented by small size crab specimens with carapace width starting from 64.6-77.0 mm (CW) (Fig. 5b).
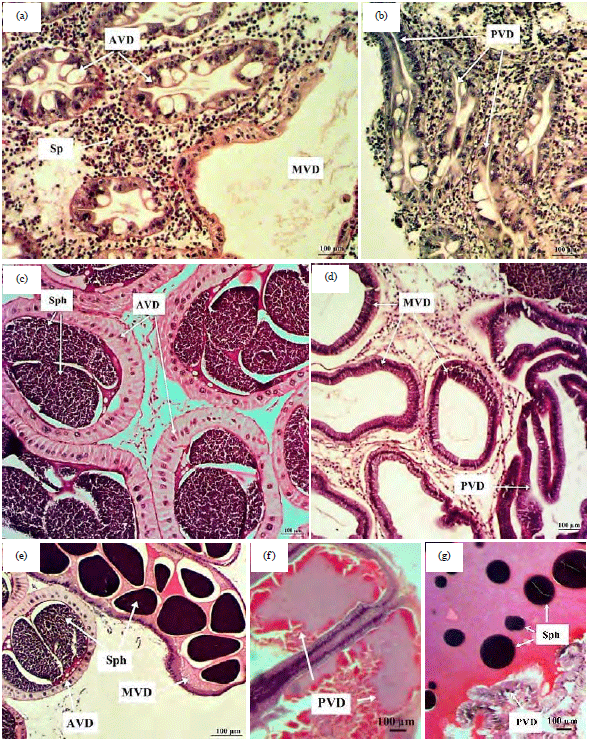 | |
| Fig. 2(a-g): | Photomicrographs of light microscope showing the vas deferens of P. pelagicus (Stage I, II and III): (a, b) Anterior vas deferens (AVD), Median vas deferens (MVD) and posterior vas deferens (PVD) were completely devoid of spermatophores in immature males, Sp: Spermatozoa, (c), d) AVD at maturing stage showing its packing with spermatophores (Sph), while MVD and PVD showing the entire lacking of spermatophores, (e) AVD and MVD at mature stage were full of spermatophores while PVD (f, g) May be empty or filled with spermatophores |
Monthly ovigerous females distribution: The ovigerous females were available from March-October, with a maximum abundance in April and August. Figure 5c described the monthly proportion of ovigerous females of P. pelagicus. In general, during the study period, the highest values of female GSI were found to be during the period of April-May and August-September. On the other hand, the occurrence of non-ovigerous females that had mature and ripe ovaries was found throughout the period from March-September with high abundance percentage from April-July.
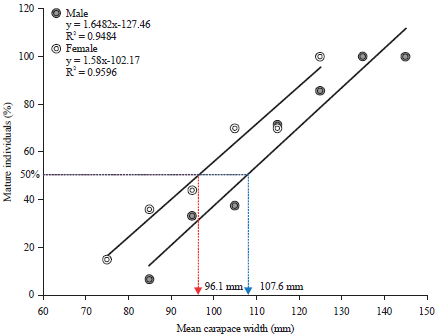 | |
| Fig. 3: | Size at 1st sexual maturity of male and female Portunus pelagicus using regression methods |
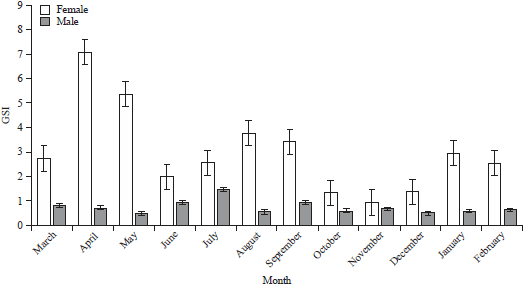 | |
| Fig. 4: | Monthly mean values of Gonadosomatic index (GSI) of males and non-ovigerous females of Portunus pelagicus |
Complementary to this, ovigerous females showed also a prolonged occurrence during the warm months from March to October as in Fig. 5c, with a maximum abundance in April and August. This tends to suggest the presence of two peaks of spawning activity, in April-May and in August-September. Hence, this indicated that ovigerous females may spawn more than one time during the spawning months.
DISCUSSION
The present study showed that the males of Portunus pelagicus are heavier than females, which is in conformity with Abdul-Sahib23 and Murni et al.12. Where Josileen8 mentioned that P. pelagicus females were marginally heavier than males up to a certain carapace size (120-125 mm) and thereafter; males became heavier than females. This could be due to the decline in the female growth during the period of gonad maturation rather than male12.
A significant positive linear relationship with high (R2) values (0.9643 and 0.9224) was observed between CW and CL for males and females, respectively. This is in agreement with Josileen8, Abdul-Sahib23 and Sukumaran and Neelakantan24, for P. pelagicus.
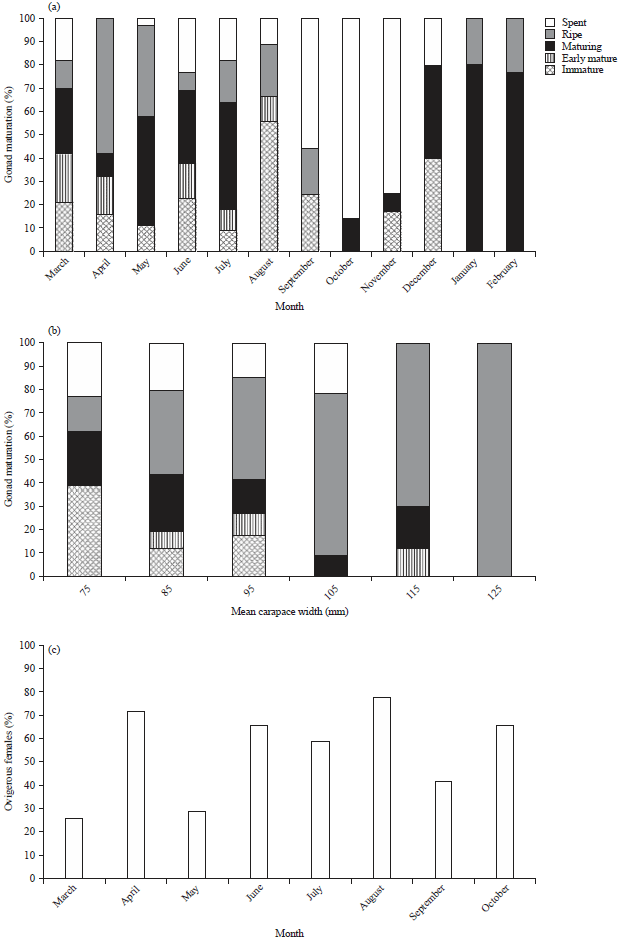 | |
| Fig. 5(a-c): | Proportion of gonad maturity stages of non-ovigerous females of P. pelagicus (a) According to months, (b) According to size (mm) and (c) Monthly occurrence of ovigerous females of P. pelagicus |
The exponential values (b) of the length-weight correlations showed that males of P. pelagicus only followed an isometric pattern of growth, which disagreed with the findings of Josileen8 and Sukumaran and Neelakantan24 for the same species as males and females showed a marked departure from isometric pattern in the Mandapam and the Karnataka coasts of India.
In the current study, the ovarian development of P. pelagicus was comprised of five stages, reflected externally and histologically, which closely agrees with Ravi et al.10. At the present study, the oocyte diameter increased as long as the ovarian maturity stage developed, leading to an increase in the size of the ovary as shown in the increased gonadosomatic index and the ovarian external observation. This agrees with the results of Islam et al.25 of the mud crab Scylla paramamosain.
Furthermore, the current results showed that (Oc I and Oc II) had basophilic cytoplasm because of its affinity for hematoxylin. Beninger et al.26 pointed out that basophilic oocytes are typically linked with previtellogenesis and devoid of yolk in most decapods. In addition, it is apparent from the current observations that Oc III in the early maturing stage had a low eosinophilic cytoplasm, while the cytoplasm of the vitellogenic oocytes (Oc III-IV) was stained evenly with eosin, taking a pink color, because of the abundant presence of yolk granules. The clear shift from basophilic to eosinophilic cytoplasm in the present study is supported by the observations of King27 in Litopenaeus setiferus and Vasudevappa28, where it was recorded only at the late vitellogenic oocyte stage of Metapenaeus monoceros.
On the other hand, for male P. pelagicus, the current findings figured out that, the histological structure of the vas deferens in immature stage and maturing stage were in agreement with the findings of De Lestang et al.3 on P. pelagicus from the west coast of Australia. It was found also in the present results, PVD may or may not contain spermatophores in mature males. De Lestang et al.3 recorded that, spermatophores were absent in PVD of mature males of P. pelagicus.
The size at 1st sexual maturity differs with latitude and locations29. Wardiatno and Fahrudin11 found that the mean size of males and females of P. pelagicus from Indonesia reached the first sexual maturity at 103mm and 98 mm (CW) respectively. Sumpton et al.30 mentioned that the smallest measured sexually mature female and male P. pelagicus were at 73.0 and 83.0 mm, respectively in the Australian waters. The differences observed between the minimum biological sizes of P. pelagicus can be explained by the effect of the difference in the site of sampling or fishing ground11.
During the present study, ovigerous females occurred in all months (except from November-February) indicating that P. pelagicus breeds almost throughout the year except in winter months. As in temperate regions, reproduction process is restricted to the warm months31. While Batoy et al.32 reported that the blue swimmer crab breeds throughout the year in tropical regions. The spawning activity of females was pronounced with maximum values of GSI during April-May. While the abundance of ovigerous females during the period of study was recorded particularly from March-October suggesting prolonged breeding in this species. This observation agrees with the findings of Sukumaran and Neelakantan33 on the blue swimmer crab from the southwest coast of India.
It is worth mentioning that the reproductive pattern analysis of P. pelagicus was performed by integration of the information concerning gonad maturation, GSI and the occurrence percentages of ovigerous females. Two peaks of spawning activity were detected in P. pelagicus, in April-May and in August-September. Hence, it is reasonable to say that ovigerous females may spawn more than one time during the spawning season. Wardiatno et al.11 also recorded a similar pattern of spawning frequency for P. pelagicus from Indonesia. Further research is urgently needed on fecundity, embryonic development and the redevelopment of female ovaries to estimate the reproductive output of P. pelagicus during the spawning season.
CONCLUSION
The size at which P. pelagicus reached sexual maturity was at 107.6 and 96.1 mm carapace width (CW) for males and females, respectively. The size is proposed to be the minimum legal size for the crab fishery sustainable use. According to patterns in gonad maturation and GSI as well as the occurrence of matured and ovigerous females, two peaks were detected in spawning season, in April-May and in August-September. These findings could be used to support the implementation of policy for blue swimming crab’s sustainable fishery in the Egyptian waters.
SIGNIFICANCE STATEMENT
Despite Portunus pelagicus is quite abundant and of high commercial value, relatively little is known about its biology from the Egyptian waters. This study presents the results of a study conducted to characterize the maturation stages of male and female P. pelagicus through external morphological and histological examination during gonad development, with determination of gonadosomatic index (GSI), size at first sexual maturity and spawning population from Alexandria coastal waters of Egypt. That will inform the gaps which need to be addressed with regard to crab production and aquaculture in Egypt and could be used to support the implementation of policy for blue swimming crab’s sustainable fishery in the Egyptian waters.
ACKNOWLEDGMENT
The authors are grateful to Dr. Amal Ragae Khafage, Ph.D., Lecturer in the laboratory of Invertebrate Aquaculture, the National Institute of Oceanography and Fisheries (NIOF), for her great experience and constant help to conduct all the histology work of this research.
REFERENCES
- De Lestang, S., N.G. Hall and I.C. Potter, 2003. Reproductive biology of the blue swimmer crab (Portunus pelagicus, Decapoda: Portunidae) in five bodies of water on the West coast of Australia. Fish. Bull., 101: 745-757.
Direct Link - Kumar, M.S., Y. Xiao, S. Venema and G. Hooper, 2003. Reproductive cycle of the blue swimmer crab, Portunus pelagicus, off Southern Australia. J. Mar. Biol. Assoc. UK., 83: 983-994.
CrossRefDirect Link - Xiao, Y. and M. Kumar, 2004. Sex ratio and probability of sexual maturity of females at size, of the blue swimmer crab, Portunus pelagicus Linneaus, off Southern Australia. Fish. Res., 68: 271-282.
CrossRefDirect Link - Johnson, D.D., C.A. Gray and W.G. Macbeth, 2010. Reproductive biology of Portunus pelagicus in a South-East Australian estuary. J. Crustacean Biol., 30: 200-205.
CrossRefDirect Link - Stewart, M.J., P. Stewart, N. Soonklang, V. Linthong, P.J. Hanna, W. Duan and P. Sobhon, 2010. Spermatogenesis in the blue swimming crab, Portunus pelagicus and evidence for histones in mature sperm nuclei. Tiss. Cell, 42: 137-150.
CrossRefDirect Link - Josileen, J., 2011. Morphometrics and length-weight relationship in the blue swimmer crab, Portunus pelagicus (Linnaeus, 1758) (Decapoda, Brachyura) from the Mandapam coast, India. Crustaceana, 84: 1665-1681.
Direct Link - Sahoo, D., S. Panda and B.C. Guru, 2011. Studies on reproductive biology and ecology of blue swimming crab Portunus pelagicus from Chilika Lagoon, Orissa, India. J. Mar. Biol. Assoc. UK., 91: 257-264.
CrossRefDirect Link - Ravi, R., M.K. Manisseri and N.K. Sanil, 2013. Ovarian maturation and oogenesis in the blue swimmer crab, Portunus pelagicus (Decapoda: Portunidae). Acta Zool., 94: 291-299.
CrossRefDirect Link - Wardiatno, Y. and A. Fahrudin, 2015. Sexual maturity, reproductive pattern and spawning female population of the blue swimming crab, Portunus pelagicus (Brachyura: Portunidae) in East Lampung coastal waters, Indonesia. Indian J. Sci. Technol., 8: 596-607.
CrossRefDirect Link - Murni, L., D. Arfiati, A. Kurniawan, Y. Kilawati and H.U. Subarijanti et al., 2018. Biological analysis of blue swimmer crab (Portunus pelagicus) from collectors in Lamongan, Tuban, Pasuruan and Rembang, Java, Indonesia. J. Exp. Life Sci., 8: 90-96.
CrossRefDirect Link - Hamid, A. and Y. Wardiatno, 2015. Population dynamics of the blue swimming crab (Portunus pelagicus Linnaeus, 1758) in Lasongko bay, Central Buton, Indonesia. Aquacult. Aquar. Conserv. Legisl., 8: 729-739.
Direct Link - Abdel Razek, F.A., S.M. Taha and A.A. Ameran, 2006. Population biology of the edible crab Portunus pelagicus (Linnaeus) from Bardawil lagoon, Northern Sinai, Egypt. Egyp. J. Aquat. Res., 32: 401-418.
Direct Link - El-Sherief, S.S., 1991. Fine structure of the sperm and spermatophores of Portunus pelagicus (L.) (Decapoda, Brachyura). Crustaceana, 61: 271-279.
Direct Link - El-Sherief, S.S., 1993. Histochemical characteristics of the egg membranes of Portunus pelagicus (L.). Acta Biol. Hung., 44: 269-280.
PubMedDirect Link - Bawab, F.M. and S.S. El-Sherief, 1988. Stages of the reproductive cycle of the female crab Portunus pelagicus (Linnaeus, 1758) based on the anatomical changes of the spermatheca (Decapoda Brachyura, Portunidae). Crustaceana, 54: 139-148.
CrossRefDirect Link - Ikhwanuddin, M., M.N. Azra, H. Siti-Aimuni and A.B. Abol-Munafi, 2012. Fecundity, embryonic and ovarian development of blue swimming crab, Portunus pelagicus (Linnaeus, 1758) in Coastal Water of Johor, Malaysia. Pak. J. Biol. Sci., 15: 720-728.
CrossRefDirect Link - Hartnoll, R.G., 1974. Variation in growth pattern between some secondary characters in crabs (Decapoda Brachyura). Crustaceana, 27: 131-136.
Direct Link - Abdul-Sahib, I.M., 2012. Some biological aspects of the swimming crab Portunus pelagicus (Linnaeus, 1766) (Decapoda: Portunidae) in NW Arabian Gulf. Mesopotamian J. Mar. Sci., 27: 78-87.
Direct Link - Sukumaran, K.K. and B. Neelakantan, 1997. Length-weight relationship in two marine portunid crabs Portunus pelagicus (Linnaeous) from the Kamataka cost. Indian J. Mar. Sci., 26: 39-42.
Direct Link - Islam, M.S., K. Kodama and H. Kurokura, 2010. Ovarian development of the mud crab Scylla paramamosain in a tropical mangrove swamps, Thailand. J. Sci. Res., 2: 380-389.
Direct Link - Beninger, P.G., C. Lanteigne and R.W. Elner, 1993. Reproductive processes revealed by spermatophore dehiscence experiments and by histology, ultrastructure and histochemistry of the female reproductive system in the snow crab Chionoecetes opilio (O. Fabricius). J. Crustacean Biol., 13: 1-16.
CrossRefDirect Link - King, J.E., 1984. A study of the reproductive organs of the common marine shrimp, Penaeus setiferus (Linnaeus). Biol. Bull., 94: 244-262.
PubMedDirect Link - Sumpton, W.D., G.S. Smith and M.A. Potter, 1989. Notes on the biology of the portunid crab, Portunus sanguinolentus (Herbst), in subtropical Queensland waters. Aust. J. Mar. Freshwater Res., 40: 711-717.
CrossRefDirect Link - Batoy, C.B., J.F. Sarmago and B.C. Pilapil, 1987. Breeding season, sexual maturity and fecundity of the blue crab, Portunus pelagicus (L.) in selected coastal waters in leyte and vicinity, Philippines. Ann. Trop. Res., 9: 157-177.
Direct Link - Sukumaran, K. K. and B. Neelakantan, 1996. Relative growth and sexual maturity in the marine crabs, Portunus (Portunus) sanguinolentus (Herbst) and Portunus (Portunus) pelagicus (Linnaeus) along the Southwest coast of India. Indian J. Fish., 43: 215-224.
Direct Link