
Fawzia Jassim Shalsh
Faculty of Agro Based Industry, University Malaysia Kelantan, Kelantan, Malaysia
Faculty of Food Science and Technology, University Putra Malaysia, 43400 UPM, Malaysia
Noor Azlina Ibrahim
Faculty of Agro Based Industry, University Malaysia Kelantan, Kelantan, Malaysia
Mohammed Arifullah
Faculty of Agro Based Industry, University Malaysia Kelantan, Kelantan, Malaysia
LiveDNA: 60.9961
Anis Shobirin Meor Hussin
Faculty of Food Science and Technology, University Putra Malaysia, 43400 UPM, Malaysia
Halal Products Research Institute, University Putra Malaysia, 43400 UPM, Malaysia
Asian Journal of Biological Sciences
Year: 2016 | Volume: 9 | Issue: 1-2 | Page No.: 10-18







ABSTRACT
Bioethanol production using lignocellulosic biomass has gained increased attention because of the abundant supply of this biomass. Saccharomyces cerevisiae is a commonly used microorganism for ethanol production. Nevertheless, S. cerevisiae can not ferment xylose, the second most abundant sugar in plant tissues. In this study, protoplast fusion with the xylose-fermenting yeast Pichia stipitis was performed to improve bioethanol production from biomass. The protoplast formation of S. cerevisiae and P. stipitis (ATCC 58785) cells was achieved using zymolase 20T. The effects of zymolase concentration, enzymatic treatment time and osmotic stabilizers were further investigated. The optimal parameter for the protoplast release of S. cerevisiae and P. stipitis included 500 μg μL–1 zymolase for 60 min and 750 μg μL–1 zymolase for 120 min, respectively. The maximum protoplast formation ratios were 98.48 and 84.42% for S. cerevisiae and P. stipitis, respectively, with 1 mol L–1 sorbitol as the osmotic stabilizer. About 4×106 mL–1 protoplasts from S. cerevisiae and P. stipitis were isolated. Protoplast fusion frequency was determined using polyethylene glycol (PEG) as fusogen. The optimized fusion conditions of S. cerevisiae protoplasts with P. stipitis required 35% (w/v) PEG 6000, 10 mM CaCl2 level and 30 min of fusion time. The protoplast fusion rate was 52.21% under the optimized fusion condition.
PDF Abstract XML References Citation
Received: October 30, 2015;
Accepted: January 01, 2016;
Published: March 15, 2016
How to cite this article
Fawzia Jassim Shalsh, Noor Azlina Ibrahim, Mohammed Arifullah and Anis Shobirin Meor Hussin, 2016. Optimization of the Protoplast Fusion Conditions of Saccharomyces cerevisiae and Pichia stipitis for Improvement of Bioethanol Production from Biomass. Asian Journal of Biological Sciences, 9: 10-18.
URL: https://scialert.net/abstract/?doi=ajbs.2016.10.18
URL: https://scialert.net/abstract/?doi=ajbs.2016.10.18
INTRODUCTION
Bioethanol production using lignocellulosic biomass has gained increased interest because of the abundant supply and relatively cheap and renewable source of this biomass (Prasad et al., 2007). Although, Saccharomyces cerevisiae is the most commonly used microorganism for the fermentation of hexose sugars in biomass and exhibits high ethanol tolerance, this organism cannot ferment xylose, which is the second major sugar component of biomass (Chandel et al., 2011). Therefore, the use of Pichia stipitis, which is native xylose-fermenting yeast, has been the focus of research; however, this species has low ethanol and sugar tolerance (Bellido et al., 2011).
Many techniques have been used to enhance the glucose and xylose fermentation ability of yeast. Protoplast fusion, mutagenesis and recombinant DNA techniques are used to modify yeast through genomic manipulation (Kumari, 2012). Protoplast fusion is a simple and widely used method to improve the preferred fermentative properties of industrial yeast strains. This method resolves the limitations of genetic alternation enforced by conventional mating systems and supports the transfer of relatively large segments of genomic DNA (Gnanam, 2013).
Zymolyase is a commercial mixture enzyme obtained from a submerged culture of Arthrobacter luteus. This enzyme shows strong lytic activity against living yeast cell walls to produce protoplasts or spheroplasts and has been proven to be a more effective reagent for the degradation of yeast cell wall for protoplast formation compared with lyticase and glusulase (Kaneko et al., 1973). The high activity of zymolyase is ascribed to its composition, which comprises a mixture of lytic enzymes that attack various cell wall polymers. Generally, the yeast cell wall consists of three major components: an inner layer of glucans (β-1, 3 and β-1, 6-glucan), chitins (polymer of N-acetylglucosamine) and an outer layer of mannoproteins (Orlean, 2012). Zymolyase includes enzymes that act on each of these polymers. The principal enzyme is β-1, 3 glucan laminaripentaohydrolase, which degrades cell wall glucans into pentamers and the second key enzyme is β-1, 3 glucanase, which hydrolyzes glucans to glucose, thereby increasing the lytic activity of the cell wall (Cabib, 2009).
In view of the importance for development of modified yeast strain that is capable of fermenting glucose and xylose sugar for bioethanol production from biomass using protoplast fusion of yeasts S. cerevisiae and P. stipitis. This study aims to determine the optimal condition for protoplast formation of S. cerevisiae and P. stipitis. The effects of several factors, such as PEG concentration, CaCl2 level and fusion time, on protoplast fusion were studied.
MATERIALS AND METHODS
Yeast strain: Saccharomyces cerevisiae was obtained from the Industrial Biotechnology Research Laboratory, University Sains Malaysia and P. stipitis (ATCC 58785) was acquired from American Type Culture Collection.
Protoplast formation: The method described by Perez-Traves et al. (2012) was adopted. Two parent yeast cells were separately grown in 100 mL of yeast extract peptone dextrose (1% yeast extract, 2% peptone and 2% glucose) medium at 32°C until they reached the log phase. Briefly, 5 mL of the 18 h old parent cultures, with an optical density at 600 nm (OD600) of 0.5 (about 107 cells mL–1, according to cell counts under the microscope), were obtained (three sets) in centrifuge tubes and centrifuged at 6000 rpm for 5 min. The harvested cells were washed twice with 0.1 mol L–1 phosphate buffer (pH 7.5). The cells were suspended in 10 mL of protoplasting solution (1.2 M sorbitol, 0.1 M Tris, 0.02 M ethylenediamine tetraacetic acid and pH 9.8) with 50 μL of β-mercapto-ethanol and then incubated for 15 min at room temperature. The cells were then washed with 1.2 M sorbitol, suspended in 10 mL of 1.2 M sorbitol with 20 T Zymolase (from Arthrobactor luteus, 200 U mL–1, Kirin Brewery Company Chuo-ku, Tokyo, Japan) and then incubated at 30°C. Protoplast release was monitored at an interval of 30 min and the number of protoplasts formed was counted using a hemocytometer. Protoplasts were collected through centrifugation at 500 rpm for 10 min, washed repeatedly with a buffer solution (0.1 mol L–1 phosphate buffer (pH 7.5) and 0.8 mol L–1 sorbitol) and suspended in the same solution. The effects of various key parameters, such as lysing enzyme concentration, lysis duration and osmotic stabilizers (such as KCl, sorbitol and MgSO4), on protoplast formation were investigated. The optimum conditions for protoplast formation were established. The protoplasts were observed under a light microscope and the number of protoplasts formed was counted using a hemocytometer.
Protoplast yield was calculated using the Eq. 1:
| (1) |
Cell concentration was determined by a spectrophotometer under the optical density at 660 nm (Gene sys 20 spectrophotometer). Cell dry weight was obtained from the conversion of optical density value by using the calibration curve.
Protoplast fusion: The protoplasts of S. cerevisiae and P. stipitis were mixed at 1:1 ratio (about 4×l06 protoplasts of each strain) and centrifuged at 500 rpm for 5 min in a buffer solution (0.1 mol L–1 phosphate buffer, pH 7.5 and 0.8 mol L–1 sorbitol). The pellets were re-suspended in fusion buffer solution (0.6 M sorbitol; 10 mM Tris-HCl, PEG and CaCl2). The effects of PEG concentrations (molecular weight 6000), CaCl2 level and fusion time were investigated and the optimum conditions of protoplast fusion were established. The suspension was incubated at 30°C and 100 rpm. Approximately 0.25 mL of the suspension was withdrawn every 10 min. Protoplast fusion was monitored under a microscope and the number of fused protoplasts were counted using a hemocytometer. The fusion frequency was determined by Eq. 2:
| (2) |
RESULTS AND DISCUSSION
Growth curve: Saccharomyces cerevisiae and P. stipitis (ATCC 58785) were used in this study (Fig. 1). Harvest of cells in exponential or log phase is the first step in protoplast formation. Cell number continues to increase and the growth pattern is shown in Fig. 2. This phase was defined as the logarithmic phase and covered the periods of 4-18 and 8-20 h for S. cerevisiae and P. stipitis, respectively. After approximately 18 h, yeast cultures were near the end of the log phase of growth. These cultures were subjected to enzyme treatment to prepare protoplasts and selected for further study because cells in this phase are younger and easily attacked by enzymes to convert to protoplasts.
Protoplast formation: Protoplasts were observed under a light microscope and the number of protoplasts formed was counted using a hemocytometer (Fig. 3a). The protoplasts are spherical in shape and can be distinguished from the oval-shaped yeast cells (Fig. 3). To investigate the optimum condition for the preparation of yeast protoplasts with high yields, we examined several factors to predict the effect of the lysis of S. cerevisiae and P. stipitis; such factors included enzyme concentration, enzymatic treatment time and osmotic stabilizer.
Effect of lysing enzyme concentration: The extent of yeast cell lysis by zymolyase varies in relation to yeast strain, growth stage of yeast and cultural condition (Aguilar-Uscanga and Francois, 2003). Sensitivity to lytic enzymes considerably varies in cells from different species and individual cells of the same species. The lysis of yeast cell wall and formation of viable protoplasts depend on the concentration of the lysing enzyme. As such, the effect of lysing enzyme on the protoplast same species. The lysis of yeast cell wall and formation of viable protoplasts depend on the concentration of the lysing enzyme. As such, the effect of lysing enzyme on the protoplast yield has been investigated at varied enzyme concentrations, ranging from 250-1000 μg μL–1.
 | |
| Fig. 1(a-b): | Yeast cells with 40×1.25 magnifications, (a) S. cerevisiae and (b) P. stipitis (ATCC 58785) |
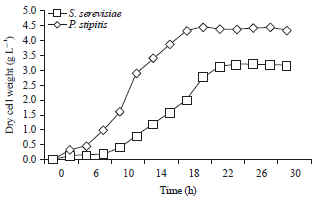 | |
| Fig. 2: | Growth curve of S. cerevisiae and P. stipitis |
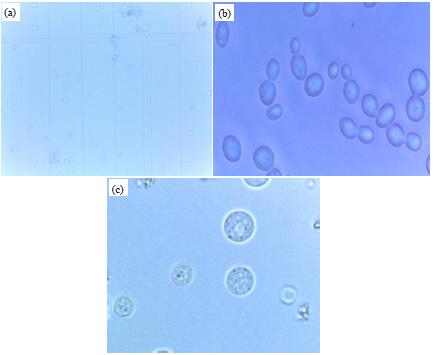 | |
| Fig. 3(a-c): | Saccharomyces cerevisiae with magnification of (a, b) 40X and (c), 100X (a) Counted using haemocytometer, (b) Intact yeast S. cerevisiae and (c) Protoplast cell |
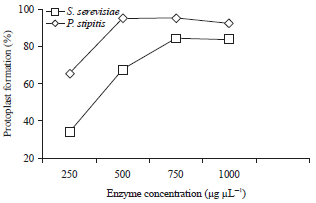 | |
| Fig. 4: | Effect of lysing enzyme concentration on protoplast formation of S. cerevisiae and P. stipitis |
As indicated in Fig. 4, the protoplast formation rate of S. cerevisiae increased as the enzyme concentration increased up to 500 μg μL–1. The rate remained almost constant at 750 μg μL–1 and then decreased thereafter with further increase in enzyme concentration. As a result of limited site access, the addition of enzyme did not affect cell wall lysis. Therefore, the rate of protoplast formation remained constant at high enzyme concentrations. A similar trend was also observed in P. stipitis yeast with a maximum yield obtained at 750 μg μL–1. The highest yield of protoplasts was observed in S. cerevisiae (98.5%), followed by P. stipitis (82%). High enzyme concentration (1000 μg μL–1) may have toxic influence, as evidenced by the lysis of protoplasts, resulting in reduced protoplast formation rate. Similar results were also reported by Balasubramanian and Lalithakumari (2008). Meanwhile, Nwachukwu et al. (2008) added 0.5 mg of Zymolase 60000 mL–1 in three S. cerevisiae strains isolated from old palm wines and incubated at 30°C for 1 h to form protoplasts.
Effect of enzymatic treatment time: Protoplast formation was severely influenced by enzymatic treatment duration because prolonged incubation of yeast cells with degraded enzymes may destruction nascent protoplasts (Bacon et al., 1969). Therefore, the effect of enzymatic treatment time on protoplast formation was examined at various time durations of 60-150 min by incubating yeast cells with the lysing enzyme zymolase. Enzymatic treatment was conducted at 30, 60, 90, 120 and 150 min. The maximum rate of protoplast formation was obtained with S. cerevisiae (95.25%) at 60 min, followed by P. stipitis (85.48%) for 120 min (Fig. 5). The obtained rates were lower than those reported by Limtong et al. (1998); in this study, 100% of S. cerevisiae M30 protoplasts were obtained through treatment with lytic enzyme solution containing 0.075 mg mL–1 Zymolyase at 20°C for 4 h. High rates of protoplast release from S. cerevisiae cells were achieved with prolonged lytic incubation time in zymolyase for 120 h. This observation agrees with the report of Ezeronye and Okerentugba (2001); in their study, the highest number of S. cerevisiae protoplasts released was obtained from cells exposed to the undiluted snail gut enzyme and incubated for 180 min.
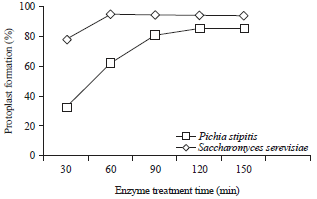 | |
| Fig. 5: | Effect of enzymatic treatment time on protoplast formation of S. cerevisiae and P. stipitis |
In the study of Zhao and Liu (2013), the condition for protoplast formation included 0.2 mg mL–1 zymolase at 30°C for 40 min. Previous studies reported that the difference in protoplast formation rates among yeast strains may be attributed to the differences in the thickness and composition of their cell wall layers (Kumari, 2012). Moreover, the variance sensitivity of yeast strains depends on enzyme degradation, different sensitivities of individual cells and resistance of older cell walls (Darling et al., 1969). Our current finding showed that after 60 and 120 min of incubation for S. cerevisiae and P. stipitis, respectively, an almost complete cell wall digestion occurred. Protoplast swelling and rounding up of cell content were monitored initially and subsequently at longer times of enzyme treatment until the protoplasts immediately burst (Hassan, 2014). Hence, 60 and 120 min were selected as the optimum enzymatic treatment duration for S. cerevisiae and P. stipitis.
Effect of osmotic stabilizers on protoplast formation: As a result of enzyme treatment, cell content would only be enclosed by the cell membrane. The protoplast should be preserved in a hypertonic medium to attain their osmotic stability and survival. The use of osmotic stabilizers (such as KCl, sorbitol, mannitol and MgSO4) is important to improve the stability of released protoplasts and inhibit lysis, leading to high yields of protoplasts. Different osmotic stabilizers, such as 0.6 M MgSO4, 1.2 M sorbitol and 0.6 M KCl, were used to determine their effects on protoplast formation. Figure 6 shows that 1 M mol–1 sorbitol exhibited the most effective protoplast formation rate for S. cerevisiae and P. stipitis. The highest protoplast formation ratios were 98.48 and 84.42% for S. cerevisiae and P. stipitis, respectively. Similar result was reported by Ezeronye and Okerentugba (2001). The highest number of protoplasts (1.59×106 protoplasts mL–1) was released in 1.0 mol L–1 sorbitol.
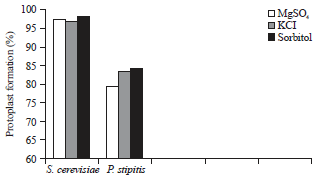 | |
| Fig. 6: | Effect of osmotic stabilizers on protoplast formation of S. cerevisiae and P. stipitis |
Approximately 1.2 M sorbitol was reported as the optimal osmotic stabilizer for protoplast formation in yeasts. Pina et al. (1986) tested different osmotic stabilizers and found that MgSO4, mannitol and sorbitol are the most suitable stabilizers. Furthermore, the obtained number of protoplasts was lower than the number of initial cells used, which may be due to the lysis of protoplasts during protoplasting. In the two cases of yeasts, S. cerevisiae and P. stipitis, the outer margin of protoplasts was irregular when the concentration of the osmotic stabilizer was changed beyond the optimum concentration. This phenomenon may be caused by the imbalance in osmotic pressure around the protoplasts.
The optimal conditions for the maximum fusion of protoplasts included PEG concentrations (molecular weight 6000) within the range of 25-40% (w/v), CaCl2 levels of 0.1-100 mM and fusion time of 10-40 min. The fusion of protoplasts was monitored under a microscope and the number of fused protoplasts was counted using a hemocytometer (Fig. 7a-b).
Protoplast fusion
Effect of PEG concentration: The PEG is the only polymeric agent currently used for yeast transformation and induced protoplast fusion, as reported by Kao and Michayluk (1974). PEG molecules exhibit polarity similar to the membrane-phospholipid molecules and thus attach to membrane proteins. When the attached PEG between the two protoplasts is removed, membranes are broken down at the contact points, resulting in protoplast fusion. The optimization of PEG concentration is essential in protoplast fusion because the toxicity of fusogen to protoplasts can be restricted using high PEG concentrations; by contrast, low PEG concentrations may not generate high frequency of fusants.
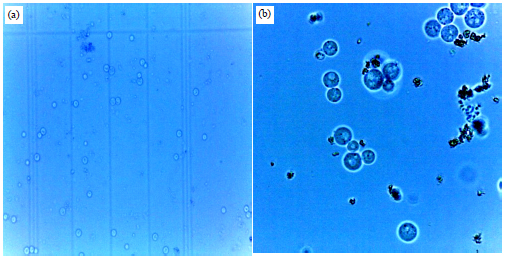 | |
| Fig. 7(a-b): | Fused protoplasts counted using haemocytometer (a) 40×1.25 magnification and (b) 100×1.25 magnification |
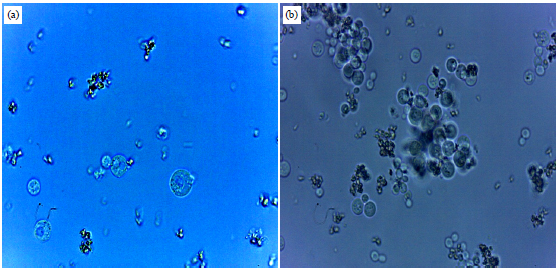 | |
| Fig. 8(a-b): | Effect of concentrations of PEG on protoplasts fusion (100×1.25 magnification), (a) low concentrations causing rupture of protoplasts and (b) higher concentrations causing aggregation and over clumping of protoplast |
For the optimized fusion of protoplasts of S. cerevisiae and P. stipitis, the concentrations of PEG (molecular weight 6000) within the range of 25-45% were investigated. Figure 8a indicates that PEG concentrations lower than 30% led to swelling, followed by rupture of protoplasts (Pina et al., 1986). Protoplast treatment with PEG concentrations higher than 35% decreased fusant formation rate. In Fig. 8b, high PEG concentrations could cause aggregation and over clumping of protoplasts. Meanwhile, the toxic effect of PEG concentrations higher than 40% destroyed yeast protoplasts. High PEG concentrations were regarded inefficient because of the difficulty in uniformly coating of viscous PEG solutions on the protoplasts. The highest number of fused protoplasts was obtained using 35% PEG (molecular weight 6000). Low PEG concentrations (<30%) did not stabilize protoplasts, thereby destructing protoplast fusion; further increase in PEG concentrations resulted in the shrinkage of protoplasts (Balasubramanian and Lalithakumari, 2008). Such as, 35% PEG was observed to be the optimal concentration.
Effect of PEG treatment time: The PEG treatment time plays important role in protoplast fusion frequency. This factor directly acts on the membrane by increasing the permeability of intact cells and enhancing their transformation efficiency and frequency (Zheng et al., 2005). The increasing treatment time of PEG causes the decline of protoplast viability because of dehydration associated with rupture (Jogdand, 2001). Based on the results, 20 min of PEG treatment at 30°C was the optimum condition to attain the highest fusion frequency (Fig. 9). A short period of PEG treatment is also necessary to induce protoplast fusion, whereas exposure of protoplasts to PEG up to 40 min decreases fusion frequency, resulting in shrinkage of fused protoplasts (Boni et al., 1981).
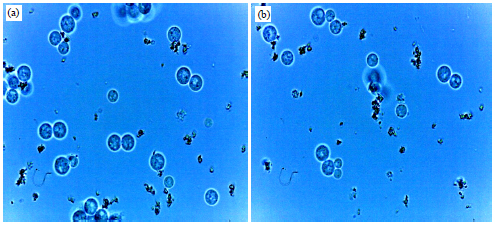 | |
| Fig. 9(a-b): | Protoplast fusion 35% PEG for 20 min exposure time at 30°C 100×1.25 magnification |
| Table 1: | Experimental optimization condition of protoplast fusion |
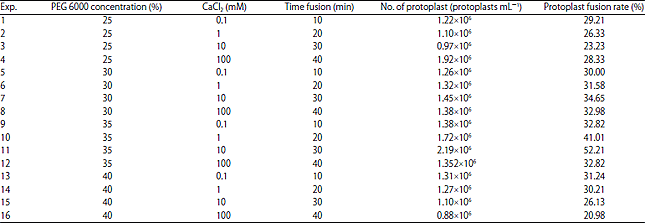 | |
| Result is the mean from three replicates of experiments | |
Yuan et al. (2004) reported that 40 min of PEG treatment at 25°C is the optimum for S. cerevisiae. The PEG treatment for 20 min favors the maximum fusion rate, whereas increasing PEG treatment time causes loss of viability of the protoplasts because of dehydration associated with rupture (Kumari, 2012).
Effect of CaCl2 (Ca2+): Physical contact of two protoplasts is essential for their fusion. However, protoplast does not easily fuse because of the net negative charge on their membrane surfaces, force of repulsion between them and difficulty to remove water from hydrophilic surface of protoplast, which create repulsive force between the two protoplasts. Positively charged ions reduce the negative charge of membrane and supply Ca2+ ions in high pH solution to reduce the repulsive force (Keller and Melchers, 1973). The high molecular weight polymer (1000-6000), PEG, acts as a molecular bridge that connects the protoplasts. Calcium ions link the negatively charged PEG and membrane surface of protoplasts, resulting in fusion. In addition, PEG causes a non-specific aggregation of protoplasts, thereby shrinking protoplasts through water withdrawal (Jogdand, 2001). For efficient protoplast fusion, PEG treatment must be accompanied by treatment with calcium ions to generate local disturbances in the membrane, leading to fusion. In the molecular mechanism of fusion, both dehydration of the membrane, which allow close contact between cellular membranes and the presence of calcium ions, which affect the membrane structure, are regarded as necessary (Papahadjopoulos et al., 1990). Zheng et al. (2005) considered PEG as the first affective factor on efficient protoplast fusion, followed by CaCl2 concentration and fusion time (Shi et al., 2014). Saccharomyces cerevisiae protoplasts were isolated from 35% PEG (molecular weight 3350), 10 mM CaCl2 and 0.8 M sorbitol for 30 min (Nwachukwu et al., 2008). By contrast, Perez-Traves et al. (2012) induced protoplast fusion of Saccharomyces for hybrid generation by using 60% PEG (molecular weight 6000) and 100 mM CaCl2 for 30 min.
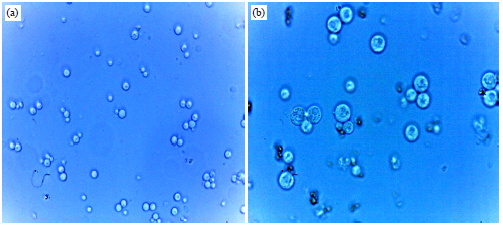 | |
| Fig. 10(a-b): | Protoplast fusions in 35% PEG with 10 mM CaCl2 after exposure for 30 min (a) 40×1.25 magnification and (b) 100×1.25 magnification |
Our current finding showed that 52.21% of S. cerevisiae protoplasts with P. stipitis were fused using 35% PEG (molecular weight 6000) and 10 mM CaCl2 at 30°C for 30 min (Table 1 and Fig. 10). The obtained fusion rate is lower than the result obtained by Kumari and Pramanik (2012), who reported that incubation in 35% PEG (molecular weight 4000) and 100 mM CaCl2 solution for 20 min could result in a fusion rate of 82.1% of S. cerevisiae protoplasts with P. stipitis, whereas 75.8% of S. cerevisiae is fused with Candida shehatae and 80.7% of S. cerevisiae is fused with Pachysolen tannophilus. Similarly, Shi et al. (2014) reported that the optimized fusion conditions of P. stipitis require 35% (w/v) PEG 6000, 10 mM CaCl2 and 30 min of fusion time to obtain a fusion rate of 0.15836%. On the fusion of the protoplasts of S. cerevisiae and Saccharomyces boulardii, Abosereh et al. (2011) used a solution of 35% PEG 4000 in 10 mM CaCl2 for 20 min and obtained a fusant percentage of 3.60%. According to Limtong et al. (1998), the protoplast fusion of S. cerevisiae M30 and Zygosaccharomyces rouxii TISTR1750 was induced by 30% PEG 6000 with 50 mM CaCl2 at 20°C for 15 min. The protoplast fusion of these two yeast genera occurred with a frequency of 5.89×10‾5.
CONCLUSION
Bioethanol production from biomass is preferred in production of alternative energy sources. Various techniques were used to develop modified yeast strains that can ferment glucose and xylose sugar components of biomass. Protoplast fusion is an important tool for gene manipulation. As an essential step toward yeast protoplast fusion to enhance bioethanol production from biomass, several parameters that affect protoplast formation and fusion of yeasts have been investigated. The optimum conditions were also established to achieve high yields of protoplasts. Many researchers have suggested different protocols for the isolation and fusion of protoplasts in yeast. In the present study, the protoplast formation of S. cerevisiae and P. stipitis (ATCC 58785) from the log phase was achieved using Zymolase 20T (from A. luteus) and found that the enzyme offered strong lytic activity against living yeast cell walls to produce yeast protoplasts. The difference in the rate of protoplast formation between these two yeast strains may be attributed to the differences in the thickness and complexity of the cell wall layers of the strains. The use of osmotic stabilizer is important to improve the stability of released protoplasts and inhibit cell lysis, leading to high yields of protoplasts. For efficient protoplast fusion, PEG treatment must be accompanied by calcium ion treatment to generate local disturbances in the membrane, resulting in fusion. Optimization of PEG concentration is also essential in protoplast fusion because the toxicity of the fusogen to protoplast cells can be restricted using high PEG concentrations, whereas low PEG concentrations may not yield high frequency of fusants.
REFERENCES
- Abosereh, N.A., E.A.M. Soliman and A.H.A. Haggran, 2011. Effect of intrespecific hybridization between Saccharomyces cerevisiae and Saccharomyces boulardii on utilization of some carbohydrates. Aust. J. Basic Applied Sci., 5: 114-120.
Direct Link - Aguilar-Uscanga, B. and J.M. Francois, 2003. A study of the yeast cell wall composition and structure in response to growth conditions and mode of cultivation. Lett. Appl. Microbiol., 37: 268-274.
CrossRefPubMedDirect Link - Bacon, J.S.D., D. Jones and P. Ottolenghi, 1969. Cell wall residues in yeast protoplast preparations. J. Bacteriol., 99: 885-887.
PubMedDirect Link - Balasubramanian, N. and D. Lalithakumari, 2008. Characteristics of protoplast inter, intra-fusant and regeneration of antagonistic fungi Trichoderma harzianum and Trichoderma viride. Afr. J. Biotechnol., 7: 3235-3243.
Direct Link - Bellido, C., S. Bolado, M. Coca, S. Lucas, G. Gonzalez-Benito and M.T. Garcia-Cubero, 2011. Effect of inhibitors formed during wheat straw pretreatment on ethanol fermentation by Pichia stipitis. Bioresour. Technol., 102: 10868-10874.
CrossRefDirect Link - Boni, L.T., T.P. Stewart, J.L. Alderfer and S.W. Hui, 1981. Lipid-polyethylene glycol interactions: II. Formation of defects in bilayers. J. Membrane Biol., 62: 71-77.
CrossRefDirect Link - Cabib, E., 2009. Two novel techniques for determination of polysaccharide cross-links show that crh1p and Crh2p attach chitin to both β(1-6)- and β(1-3) glucan in the saccharomyces cerevisiae cell wall. Eukaryotic Cell, 8: 1626-1636.
CrossRefDirect Link - Chandel, A.K., G. Chandrasekhar, K. Radhika, R. Ravinder and P. Ravindra, 2011. Bioconversion of pentose sugars into ethanol: A review and future directions. Biotechnol. Mol. Biol. Rev., 6: 8-20.
Direct Link - Darling, S., J. Theilade and A. Birch-Andersen, 1969. Kinetic and morphological observations on Saccharomyces cerevisiae during spheroplast formation. J. Bacteriol., 98: 797-810.
Direct Link - Ezeronye, O.U. and P.O. Okerentugba, 2001. Optimum conditions for yeast protoplast release and regeneration in Saccharomyces cerevisiae and Candida tropicalis using gut enzymes of the giant African snail Achatina achatina. Lett. Applied Microbiol., 32: 190-193.
CrossRefDirect Link - Hassan, M.M., 2014. Influence of protoplast fusion between two Trichoderma spp. on extracellular enzymes production and antagonistic activity. Biotechnol. Biotechnol. Equipment, 28: 1014-1023.
CrossRefDirect Link - Kaneko, T., K. Kitamura and Y. Yamamoto, 1973. Susceptibilities of yeasts to yeast cell wall lytic enzyme of Arthrobacter luteus. Agric. Biol. Chem., 37: 2295-2302.
Direct Link - Kao, K.N. and M.R. Michayluk, 1974. A method for high-frequency intergeneric fusion of plant protoplasts. Planta, 115: 355-367.
CrossRefDirect Link - Keller, W.A. and G. Melchers, 1973. The effect of high pH and calcium on tobacco leaf protoplast fusion. Zeitschrift Fur Naturforschung C, 28: 737-741.
Direct Link - Kumari, R. and K. Pramanik, 2012. Improved bioethanol production using fusants of Saccharomyces cerevisiae and xylose-fermenting yeasts. Applied Biochem. Biotechnol., 167: 873-884.
CrossRefDirect Link - Limtong, S., S. Deejing, W. Yongmanitchai and W. Santisopasri, 1998. Construction of high ethanol fermenting halotolerant hybrid by intergeneric protoplast fusion of Saccharomyces cerevisiae and Zygosaccharomyces rouxii. Kasetsart J., 32: 213-223.
Direct Link - Nwachukwu, I.N., V.I. Ibekwe, R.N. Nwabueze, B.N. Anyanwu, U. Ezeji, I. Kalu and E. Chinakwe, 2008. Production of high-ethanol-yielding Saccharomyces cerevisiae of palm wine origin by protoplast fusion. Life Sci. J., 5: 64-68.
Direct Link - Orlean, P., 2012. Architecture and biosynthesis of the Saccharomyces cerevisiae cell wall. Genetics, 192: 775-818.
CrossRefDirect Link - Papahadjopoulos, D., S. Nir and N. Duzgunes, 1990. Molecular mechanisms of calcium-induced membrane fusion. J. Bioenerget. Biomemb., 22: 157-179.
CrossRefDirect Link - Perez-Traves, L., C.A. Lopes, E. Barrio and A. Querol, 2012. Evaluation of different genetic procedures for the generation of artificial hybrids in Saccharomyces genus for winemaking. Int. J. Food Microbiol., 156: 102-111.
CrossRefDirect Link - Pina, A., I.L. Calderon and T. Benitez, 1986. Intergeneric hybrids of Saccharomyces cerevisiae and Zygosaccharomyces fermentati obtained by protoplast fusion. Applied Environ. Microbiol., 51: 995-1003.
Direct Link - Prasad, S., A. Singh and H.C. Joshi, 2007. Ethanol as an alternative fuel from agricultural, industrial and urban residues. Resour. Conserv. Recycl., 50: 1-39.
CrossRefDirect Link - Shi, J., M. Zhang, L. Zhang, P. Wang, L. Jiang and H. Deng, 2014. Xylose-fermenting Pichia stipitis by genome shuffling for improved ethanol production. Microb. Biotechnol., 7: 90-99.
CrossRefDirect Link - Yuan, Y., X. Guo, X. He, B. Zhang and S. Liu, 2004. Construction of a high-biomass, iron-enriched yeast strain and study on distribution of iron in the cells of Saccharomyces cerevisiae. Biotechnol. Lett., 26: 311-315.
CrossRefDirect Link - Zheng, H.Z., H.H. Liu, S.X. Chen, Z.X. Lu and Z.L. Zhang et al., 2005. Yeast transformation process studied by fluorescence labeling technique. Bioconjugate Chem., 16: 250-254.
CrossRefDirect Link