
Monica Nainta
Laboratory of Parasitology and Immunology, Department of Biosciences, Himachal Pradesh University, 171005, Shimla, India
H.S. Banyal
Laboratory of Parasitology and Immunology, Department of Biosciences, Himachal Pradesh University, 171005, Shimla, India
Asian Journal of Biological Sciences
Year: 2013 | Volume: 6 | Issue: 5 | Page No.: 265-270







ABSTRACT
Humoral response exhibited by a fraction of Plasmodium berghei is evaluated in the present study. Immunization of mice with S fraction (supernatant of 20,000 rpm centrifugation) of Plasmodium berghei exhibited high humoral immune response. S fraction contained proteins of molecular weight ranging from 23.15-205.00 kDa. The humoral response was serodiagnostically analyzed employing ELISA, IFA and in vitro invasion inhibition assays. Antimalarial antibodies significantly inhibited invasion of Plasmodium berghei in short-term invasion assay. From the in vitro culture study it was found that S fraction exhibits a high humoral response and sera of five out of ten immunized mice inhibited more than 50% of invasion of erythrocytes by P. berghei.
PDF Abstract XML References Citation
Received: September 14, 2013;
Accepted: November 26, 2013;
Published: February 06, 2014
How to cite this article
Monica Nainta and H.S. Banyal, 2013. Antibodies Against Plasmodium berghei Fraction Inhibit Parasite Propagation
in vitro. Asian Journal of Biological Sciences, 6: 265-270.
URL: https://scialert.net/abstract/?doi=ajbs.2013.265.270
URL: https://scialert.net/abstract/?doi=ajbs.2013.265.270
INTRODUCTION
Malaria is one of the most dreadful diseases caused by a protozoan parasite, Plasmodium that has proved to be a hinderance to the cultural and socio-economic progress of man in tropical and subtropical regions of the world. Resurgence of malaria resulted due to increased resistance to conventional antimalarial agents, both chemotherapeutic and chemoprophylactic, absence of an effective malaria vaccine and weakening of malaria control programme. Due to lack of availability of funds, future of malaria control programme has become uncertain (WHO, 2011).
The antigens of parasite play an important role in malaria control. An exposure of the host with parasite antigens for longer duration activates both the humoral and cell-mediated immune responses. Although many antigens of parasite have been identified but a large number of them still remain unexplored which may impart long lasting immunity. In this study, the cytosolic constituents of P. berghei obtained after centrifugation at 20,000 rpm from asexual erythrocytic stages of parasite exhibited humoral response in mice and effectively inhibited in vitro propagation of parasite.
MATERIALS AND METHODS
Maintenance of parasite: The asexual erythrocytic stages of Plasmodium berghei (NK-65), a rodent malaria parasite, were maintained in white Swiss mice, Mus musculus, BALB/c strain with permission of Institutional Animal Ethics Committee. The parasite was maintained in mice of either sex of about 20-25 gm weight, by passing the blood from infected mice to naïve intraperitoneally (i.p.) so that each mouse gets 1x105 P. berghei infected erythrocytes in citrate saline (Banyal et al., 1991).
Isolation of cell free Plasmodium berghei: Blood was collected from P. berghei infected mice for the isolation of cell-free parasite. When the percent infection was more than 50%, the mice were anaesthetized with diethyl ether. Blood was collected by jugular vein incision in citrate saline in the ratio of 2:1. Cell-free P. berghei was isolated according to the method of Banyal and Fitch (1982).
Differential centrifugation: Differential centrifugation of the homogenized parasite was carried by centrifugation at 20,000 rpm (Sigma 3K 30) for 35 min at 4°C. The pellet was discarded and the supernatant was used for immunization.The supernatant was designated fraction S.
Protein estimation: Protein was estimated spectrophotometrically using bovine albumen as standard (Banyal and Kumar, 1994).
Immunization of mice and collection of sera: A group of 10 mice was immunized with fraction S. Each mouse was given 100 μg of protein along with 30 μg saponin as adjuvant in 0.01 M PBS, pH 7.2 intraperitoneally on day 0,14 and 28. Placebo controls received only saponin (30 μg mouse-1 dose-1) in PBS. A week after the last immunization dose all the mice were sacrificed, sera collected separately and stored at -20°C till further use.
Enzyme linked immunosorbent assay (ELISA): Enzyme linked immunosorbent assay (ELISA) was performed according to Banyal and Inselburg (1985) in 96 well polystyrene microtitre plates.
Indirect fluorescent antibody (IFA) test: Indirect Fluorescent Antibody (IFA) test was carried according to Collins and Skinner (1972) as given by Upma and Banyal (1998).
Sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE): The antigens were analysed to determine their molecular weights by SDS-PAGE according to Kumar and Banyal (1997).
In vitro merozoite invasion inhibition assay: The short term in vitro culture of P. berghei was carried out as given by Upma and Banyal (1998).
Culture medium: RPMI-1640 (Gibco) was used as culture medium. It was supplemented with 0.06% (w/v) N-2-hydroxyethyl piperazine-N’-2-ethane sulphonic acid (HEPES): 5% (w/v) sodium bicarbonate; antibiotics gentamycin (50 μg mL-1), penicillin (100 μL mL-1) and streptomycin (100 μg mL-1). The pH of incomplete medium was adjusted to 7.4, it was filtered through 0.22 μ millipore syringe filter under sterile conditions. Ten percent (v/v) inactivated Foetal Calf Serum (FCS) was added to incomplete medium and the complete medium was prepared.
Invasion inhibition assay: The invasion inhibition assay was carried out in 12 well culture trays (TPP, Switzerland). To each well 1 mL of complete medium having 3% haematocrit and 0.3-0.8% parasitaemia along with 50 μL of normal/immune mouse serum was added. Each sample was run in duplicate. The trays were then shaken gently to mix contents. The sample was taken, centrifuged and from resulting pellet portion 0 h smears were prepared. The culture trays were placed in a candle jar at 37°C. Culture trays were kept for 21 h in the incubator, then removed from the incubator and smears from each well were prepared after centrifugation of different samples. Smears were fixed in methanol and stained with Giemsa stain. A differential count of the parasite (rings, trophozoites and schizonts) in smears was done. The merozoite invasion inhibition percent was calculated as:
RESULTS
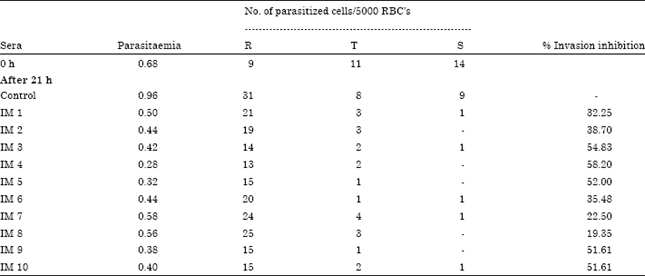
Immunization of mice with fraction S resulted in appearance of antimalarial antibodies. Indeed sera of ten mice the ELISA titre was 1:1024 in three mice, 1:2048 in three mice,1:4096 in two mice, 1:8192 and 1:16384 in one mouse each (Fig. 1). Immunoflorescence reactions were observed in immune sera as observed under UV light (Fig. 2).The short term in vitro invasion inhibition assay revealed a significant inhibition of propagation of P. berghei in vitro in the presence of immune sera. After 21 h of incubation a significant number of rings appeared in the normal sera culture (Table 1 ). In the sera of five out of ten mice immunized with fraction S of P. berghei more than 50% inhibition of propagation of parasite was observed. The number of newly formed rings in the experimental culture compared to the normal control culture was significantly less. Table 1 also revealed that there was maturation of parasite in the in vitro system.
Sds page: The S fraction was subjected to SDS PAGE analysis. There were 18 protein bands ranging from 23 to 205 kDa (Fig. 3).
In vitro invasion inhibition assay: The parasite inhibitory property of antimalarial antibodies present in sera from immunized mice were demonstrated by short term in vitro invasion inhibition assay. P. berghei infected red cells mainly consisting of trophozoite and schizonts were incubated along with normal red cells in RPMI-1640 supplemented with foetal calf serum. After 21 h of incubation there was a increase in rings as compared to 0 h in the control normal serum culture.
| Table 1: | Merozoite invasion inhibition in vitro in the presence of immune sera |
 | |
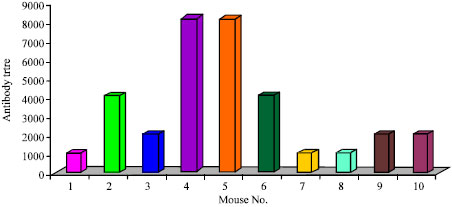 | |
| Fig. 1: | Histogram showing level of antibodies in immune sera of mice analysed by ELISA |
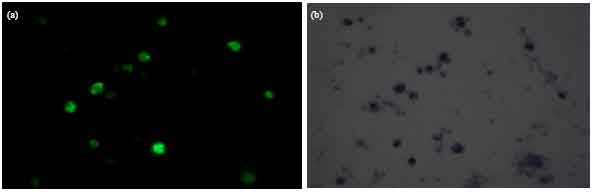 | |
| Fig. 2(a-b): | IFA reaction seen under UV light and phase contrast (χ1000), (a) Immune serum of immunized mouse (UV) and (b) Immune serum of immunized mouse (Phase contrast) |
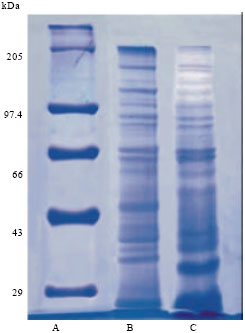 | |
| Fig. 3: | SDS page lane, (a) Molecular weight marker, (b) Total parasite homogenate and (c) Fraction S |
Effect of immune sera: The in vitro invasion inhibition assay was performed using the sera from mice immunized with S fraction of parasite. The smears prepared from culture supplemented with immunized sera after 21 h of incubation showed that the number of trophozoites and schizonts decreased as compared to 0 h and the number of rings in the experimental were less compared to the control. The percent invasion inhibition observed with serum of 10 mice as 32.25, 38.70, 54.83, 58.20, 52.00, 35.48, 22.5, 19.35, 51.61 and 51.61% (Table 1).
DISCUSSION
Plasmodium berghei (NK-65) generally proved lethal to white Swiss mice (BALB/c) has been confirmed in the present investigation. The immunization of mice with P. berghei results in induction of both the humoral and cell-mediated immune responses BALB/c strain of white Swiss mice. The present work investigated the role of S (supernatant of 20,000 rpm) fraction of P. berghei in providing immunity. This fraction exhibited a strong humoral immune response. Serodiagnostic tests like IFA and ELISA were conducted. The increase in antibody level may be due to activation of memory cells which released antiparasite antibodies in blood. This shows that antigens present in S subcellular fraction of parasite triggered the humoral response. The immune sera obtained from immunized mice were further used for in vitro invasion-inhibition assay. The results of in vitro invasion-inhibition assay showed inhibition of parasite invaded to different levels.
Since this fraction contains 18 bands of molecular weight ranging from 23.15-205.00 kDa, it appears that some of these proteins have acted as antigen. Three antigens of molecular weight 30, 40, 105 kDa have been reported from asexual blood stages of P. falciparum which are inhibitory in vitro (Banyal and Inselburg, 1985). In another study, antigens of molecular weights 80 and 95 kDa have been shown to induce strong humoral response as evident by IFA and in vitro inhibition of merozoite invasion. Eighty three kDa protein could be the apical membrane antigen (AMA-1), another leading vaccine candidate which is a target for antibody dependent immune response and reported in all Plasmodium species. AMA-1 in combination with MSP-1 have also been evaluated for its efficacy against blood-stage malaria in animal models (Burns et al., 2003). Unavailability of effective antimalarial vaccine has become one of the major cause of immense suffering and despite extensive efforts malaria has not been completely eradicated. Most malaria resurgences have been linked to weakening of control programme (Cohen et al., 2012).
From the in vitro culture studies it was found that S fraction exhibits a strong humoral response and sera from mice immunized with this fraction showed in vitro inhibition against malaria invasion, this fraction can be used as candidate for effective malaria vaccine. The antigens of parasite need to be explored further so as to know their protective role against malaria or their cross reactivity with other antigens of simian or human malaria parasites.
ACKNOWLEDGMENT
Ms Monica Nainta is grateful to DST (New Delhi) for Inspire fellowship.
REFERENCES
- Banyal, H.S. and C.D. Fitch, 1982. Ferriprotoporphyrin IX binding substances and the mode of action of chloroquine against malaria. Life Sci., 31: 1141-1144.
PubMed - Banyal, H.S. and J. Inselburg, 1985. Isolation and characterization of parasite-inhibitory Plasmodium falciparum monoclonal antibodies. Am. J. Trop. Med. Hyg., 34: 1055-1064.
PubMed - Burns, J.M., Jr., P.R. Flaherty, M.M. Romero and W.P. Weidanz, 2003. Immunization against Plasmodium chabaudi malaria using combined formulations of apical membrane antigen-1 and merozoite surface protein-1. Vaccine, 21: 1843-1852.
CrossRefDirect Link - Cohen, J.M., D.L. Smith, C. Cotter, A. Ward, G. Yamey, O.J. Sabot and B. Moonen, 2012. Malaria resurgence: A systematic review and assessment of its causes. J. Malaria, Vol. 11.
CrossRefDirect Link - Collins, W.E. and J.C. Skinner, 1972. The indirect fluorescent antibody test for malaria. Am. J. Trop. Med. Hyg., 21: 690-695.
Direct Link - Kumar, S. and H.S. Banyal, 1997. Purification and characterization of the hexokinase of P. berghei, a murine malaria parasite. Acta Vet. Hung., 45: 119-126.
PubMed - Upma, J.A. and H.S. Banyal, 1998. A 24,000g sediment of Plasmodium berghei induces IL-1 response in mice and exhibits protection against malaria infection. Parasit. Hung., 31: 13-21.
Direct Link