
F. Yarahmadi
Department of Plant Protection, Faculty of Agriculture, Ramin Agricultural and Natural Resources University, Ahwaz, Iran
A. Rajabpour
Department of Plant Protection, Faculty of Agriculture, Ramin Agricultural and Natural Resources University, Ahwaz, Iran
Asian Journal of Biological Sciences
Year: 2012 | Volume: 5 | Issue: 1 | Page No.: 52-56







ABSTRACT
Myzocallis coryli is one of the important pests of hazelnut in Iran. Seasonal population dynamics and spatial distribution of the aphid were studied during 2010-2011 in Lorestan, Iran. The aphid densities were monitored weekly and spatial distribution was evaluated by Taylor's power law. M. coryli was observed during April to July. During the months, average weather temperature was 12.5-31.5°C. Population peaks of the aphid were recorded in end of May and beginning of June during 2010 and 2011, respectively. The adult and nymph densities of M. coryli decreased with increasing weather temperature higher than 25°C. Spatial distribution of the aphid was aggregated on hazelnut.
PDF Abstract XML References Citation
Received: December 01, 2011;
Accepted: December 10, 2011;
Published: March 06, 2012
How to cite this article
F. Yarahmadi and A. Rajabpour, 2012. Seasonal Population Dynamics and Spatial Distribution of Myzocallis coryli Goetze on Corylus avellana in Iran. Asian Journal of Biological Sciences, 5: 52-56.
URL: https://scialert.net/abstract/?doi=ajbs.2012.52.56
URL: https://scialert.net/abstract/?doi=ajbs.2012.52.56
INTRODUCTION
The European hazelnut, Corylus spp. L. is one of the world major nut crops (Boccacci and Botta, 2008). Its geographic distribution extends from the Mediterranean coast of northward to British Isles and Scandinavian Peninsula and eastward to Ural Mountains of Russia, the Caucasus Mountain, Iran and Lebanon (Boccacci and Botta, 2008). Iran hazelnut production is about 14000 metric tons from 18000 hectares. Filbert aphid, Myzocallis coryly is one of the most important pests of Corylus avellana L. in Iran (Alikhani et al., 2010). M. coryli is cosmopolitan aphid pest that widely distributed in Europe. The aphid feed and breeds on the underside leaves of host plants and forming open colonies of scattered individuals. The aphid excretes sticky honeydew which drops down and contaminates the underlying foliage. Foliage on infested trees is not distorted. However, leaves may become contaminated by honeydew and blackened by sooty mould that reduces photosynthetic activity (Alford, 2007). M. coryli is a serious pest of commercial hazelnut in areas such as western Oregon, USA, Turkey and England (Naeem and Compton, 2000; Tuncer et al., 1997).
Decision-making in integrated pest management program is based on information about pest density and its spatial distribution (Messing and Aliniazee, 1989; Madadi et al., 2011; Pedigo, 2002). Analysis of spatial distribution pattern is recognized as a necessary procedure for insect population studies and provides basic information for designing efficient and cost-effective sampling plans for population estimation and pest management (Madadi et al., 2011). The present studies were conducted to determine seasonal population dynamics and spatial distribution of M. coryli on C. avellana in Lorestan province, West of Iran.
MATERIALS AND METHODS
Experimental design: The experiments were carried out during April 2010 to October 2011 at a commercial orchard (8 ha) in Boroujerd, Lorestan, Iran. The orchard included twenty 22-years-old hazelnut trees that distributed among other trees. No insecticide treatments were applied during trial period.
Seasonal population dynamics of Myzocallis coryli: The population densities of M. coryli were monitored weekly by leaf count method. Ten hazelnut trees were chosen and tagged to sampling. Each sampling date, three leaves from different height levels (top, middle and bottom) of the each side (North, east, west and south) of the plant canopy were randomly chosen and the adult and nymph stages of the aphid were counted in situ by a 20Χ LED lighted loupe magnifier.
Spatial distribution: The spatial distribution of the aphid on hazelnut was evaluated by using the parameters of Taylor's power law. This law describes the regression between logarithm of population variance and logarithm of population mean according to the equation as follows:
where, S2 is the population variance, ![]() is population mean, ‘a’ is the Y-intercept and ‘b’ is slope of regression line. This equation can transform as follows:
is population mean, ‘a’ is the Y-intercept and ‘b’ is slope of regression line. This equation can transform as follows:
where, ‘a’ is the antilogarithm of ‘a’ and constitutes a scaling factor depending on the sampling unit and ‘b’ is an index of organism species spatial pattern with b<1, b = 1 and b>1 indicating uniform, random and aggregated spatial pattern, respectively (Southwood, 1978; Tomanovic et al., 2008). Also, Correlation coefficient (r) was calculated to goodness of fit of Taylor's power law. Two tailed t-test at n-2 df was conducted to determine if slope and correlation coefficient values of the regression relation differ significantly from 1 and 0, respectively (Snedecor and Cochran, 1980).
Data analyses: Analysis of variance (ANOVA) was performed to compare population densities of M. coryli in different heights and sides of hazelnut canopy. All analyses were carried out using the SPSS software version 16 (SPSS Inc., Chicago, USA).
RESULTS
Seasonal population dynamics of Myzocallis coryli: Population dynamics of M. coryli on hazelnut were shown in Fig. 1 and 2 during 2010 and 2011, respectively. The aphid observed on hazelnut during moderate temperature months of year (April to July) in Lorestan province. During the months, average weather temperature was 12.5-31.5°C.
During 2010, the first adults and nymphs were observed at 12 April. The adult densities increased slowly from 0.5 to 9.3 aphids per leaf during 12 April to 3 June. Adult densities peaked at a mean of 9.3 aphids per leaf on 3 June. The adult decreased gradually after 26 June. The nymph densities built up exponentially from 2 to 47.7 aphids per leaf during 12 April to 2 May. From 2 May to 10 June, population of the nymphs fluctuated around 35-42 aphids per leaf. Peak densities of nymph stage was 47.7 aphids per leaf at 3 June.
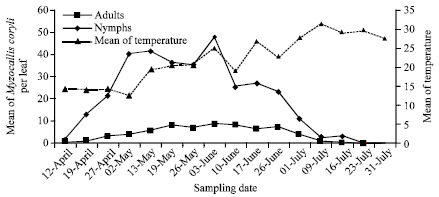 | |
| Fig. 1: | Seasonal population dynamics of Myzocallis coryli on hazelnut in 2010 |
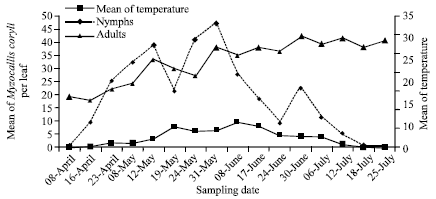 | |
| Fig. 2: | Seasonal population dynamics of Myzocallis coryli on hazelnut in 2011 |
Nymph population decreased from 3 June to end of sampling period. No adult and nymph stages of the aphid were observed after 16 July during 2010 (Fig. 1).
In 2011, adult densities of M. coryli increased from 0 to 9.3 aphids per leaf from 8 April to 8 June. Adult peak density of 46.9 aphids per leaf was recorded at 31 May. The population densities of adults decreased after 31 May as reached 0.05 aphids per leaf at 18 July.
The nymph densities were rapidly increased from 9.6 to 38.7 aphids per leaf during 8 April to 12 May. Three distinct density peaks of 38.7, 46.9 and 22.5 aphids per leaf were observed at 12 May, 31 May and 30 June, respectively. The nymph densities were dropped after 30 June. Nymph and adult stages of the aphid finally disappeared in samples in 25 July.
No significant differences were found between population densities of the aphid in different heights and sides of hazelnut canopy (df = 11, 192; F = 1.86; p>0.05) (Fig. 2).
Spatial distribution: The regression relationship between logarithm mean and logarithm variance of M. coryli is showed in Fig. 3. The correlation coefficient value for Taylor's model was significantly different from 0. Therefore, this model accurately describes the mean-variance linear relation for the M. coryli data set (p = 0.00, df = 19, t = 9.7). Slope value of the regression (b) was significantly different from 1(p = 0.00, df = 19, t = 7.8) that indicated an aggregated distribution pattern for M. corylion hazelnut leaves.
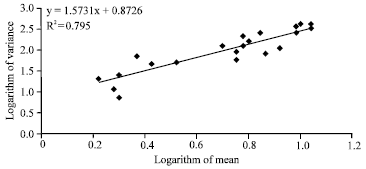 | |
| Fig. 3: | Regression relationship between logarithm mean and logarithm variance of Myzocallis coryli |
DISCUSSION
M. coryli was observed during moderate months of year (from August to July) on hazelnut. In this period, weather temperature was 12.5-31.5°C in Boroujerd during 2010-2011. The adult and nymph densities decreased with increasing weather temperature above 25°C. The negative effect of high temperature on aphids has also been reported by other authors. For example, temperature above 25°C had a harmful effect on Macrosiphum rosae L. (Mehrparvar and Hatami, 2007). Wang et al. (2002) showed that temperatures higher than 32°C reduced developmental rate of Aphis spiraecola. Similarly, study of Satar and Yokomi (2002) indicated that developmental rate of Brachycaudus scwartzi nymphs decreased at temperature above 25°C.
Result showed that M. coryli observed on hazelnut during April to July in Lorestan province, Iran. This result conflicts with those observed by Naeem and Compton (2000). They showed that the aphids were presented during May to September and peaked at July in North of England. Different climatic condition of west of Iran and north of England may be due to the conflict result. Aphid populations from geographically separated areas respond to temperature differently (Olmez et al., 2003; Mehrparvar and Hatami, 2007).
Havelka and Stary (2007) reported that Myzocallis walshii Monell observed eight months (from April to November) on red oak, Quercus rubra L. in Czech Republic. Differences between aphid species, climatic condition and host plant may be due to conflict of they results with our study. Kidd (1985) showed that host plant quality affect population dynamic of aphids.
Spatial distribution of M. coryli was aggregative on hazelnut. Analysis of spatial distribution pattern is recognized as a necessary procedure for insect population studies and provides basic information to designing efficient and cost effective sampling plans for population estimation and pest management (Madadi et al., 2011).
ACKNOWLEDGMENTS
We thank S. Yarahmadi, Dr. Mohiseni, Pirhadi for their technical assistance. Research received financial support from research deputy of Ramin Agricultural and Natural Resources University which is appreciated.
REFERENCES
- Alikhani, M., A. Rezwani, E. Rakhshani and S.M.J. Madani, 2010. Survey of aphids (Hem., Aphidoidea) and their host plants in central parts of Iran. J. Entomol. Res., 2: 7-16.
Direct Link - Boccacci, P. and R. Botta, 2008. Genetic diversity of hazelnut (Corylus avellana L.) germplasm in Northeastern Spain. HortScience, 43: 667-672.
Direct Link - Kidd, N.A.C., 1985. The role of the host plant in the population dynamics of the large pine aphid, Cinara pinea. Oikos, 44: 114-122.
Direct Link - Madadi, H., F. Dashti and A. Enkegaard, 2011. Spatial distribution of Thrips tabaci and development of a fixed-precision sampling plan for greenhouse cucumber. J. Entomol., 8: 280-287.
CrossRefDirect Link - Mehrparvar, M. and B. Hatami, 2007. Effect of temperature on some biological parameters of and Iranian population of rose aphid, Macrosiphum rosae (Hemiptera: Aphididae). Eur. J. Entomol., 104: 631-634.
Direct Link - Messing, R.H. and M.T. Aliniazee, 1989. Introduction and establishment of Trioxys pallidus [Hym.: Aphidiidae] in Oregon, U.S.A. for control of filbert aphid Myzocallis coryli [Hom.: Aphididae]. BioControl, 34: 153-163.
CrossRef - Naeem, M. and S. Compton, 2000. Population dynamics of filbert aphid, Myzocallis coryli (Goetze) on hazel bushes to an agroforestry system. Pak. J. Biol. Sci., 3: 306-308.
CrossRefDirect Link - Olmez, S., E. Bayhan and M.R. Ulusoy, 2003. Effect of different temperatures on the biological parameters of Macrosiphum rosae (L.) (Homoptera: Aphididae). J. Plant Dis. Protec., 110: 203-208.
Direct Link - Satar, S. and R. Yokomi, 2002. Effect of temperature and host on development of Brachycaudus schwartzi (Homoptera: Aphididae). Ann. Entomol. Soc. Am., 95: 597-602.
Direct Link - Tomanovic, Z., N.G. Kavallieratosz and C. Athanassiou, 2008. Spatial distribution of cereal aphids (Hemiptera: Aphidoidea) in Serbia. Acta Entomol. Serbica, 13: 9-14.
Direct Link - Tuncer, C., O. Ecevit and I. Akca, 1997. Observations on biology of the filbert aphid (Myzocallis coryli, Homoptera: Aphididae) in hazelnut orchards. Acta Hortic., 445: 485-492.
Direct Link - Wang, Y.H., L. Su and J.X. Wu, 2002. Effect of temperature on the population increase of corn leaf aphid, Rhopalosiphum maidis. Entomol. Knowl., 39: 277-280.
Direct Link