
Hussien M. Alwadie
Department of Biology, College of Sciences, King Khalid University, Abha P.O. Box 2004, Saudi Arabia
Asian Journal of Biological Sciences
Year: 2008 | Volume: 1 | Issue: 1 | Page No.: 45-50







ABSTRACT
The pollen morphology of six species of aquatic angiosperms from Saudi Arabia belonging to five genera distributed in five families was examined using both light and scanning electron microscopy. Based on apertural type, three distinct pollen types are recognized. Pollen Type-I is characterized by its non-aperturate pollen such as Elodea canadensis, Potamogeton crispus, P. pectinatus and Ruppia maritima. Pollen Type-II is easily recognized by having porate pollen such as Lemna gibba. Pollen Type-III is readily delimited by having colpate pollen such as Myriophyllum spicatum. The correlation between pollen morphology and pollination mechanism is discussed.
PDF Abstract XML References Citation
How to cite this article
Hussien M. Alwadie, 2008. Pollen Morphology of Six Aquatic Angiosperms from Saudi Arabia. Asian Journal of Biological Sciences, 1: 45-50.
URL: https://scialert.net/abstract/?doi=ajbs.2008.45.50
URL: https://scialert.net/abstract/?doi=ajbs.2008.45.50
INTRODUCTION
Aquatic angiosperms are the conspicuous plants that dominate wetlands, shallow lakes and streams. An aquatic plant can be defined as one that is normally found growing in association with standing water whose level is at or above the surface of the soil (Arber, 1963; Sculthorpe, 1967; Cook, 1974). Aquatic macrophytes have served humans well over the centuries, providing food, medicines and building materials and play a vital role in healthy ecosystems. They serve as primary producers of oxygen through photosynthesis, provide a substrate for algae and shelter for many invertebrates, aid in nutrient cycling to and from the sediments and help stabilize river and stream banks (Kaufman, 1989).
Aquatic angiosperms are divided into four classes (free-floating, submersed, floating-leaved and emergent) based on whether they are rooted in the substrate and their leaf locations in relation to the water surface (Bell, 1991). The flowers of most aquatic angiosperms must be elevated above the water in order for pollination to occur; entomophily is pollination by insects and anemophily is by wind; very rarely does pollen transfer occurs underwater (hydrophily) (Riemer, 1984).
Aquatic angiosperms represent a considerable part of the world’s flora. About 79 families and 380 genera contain aquatic species (Cook, 1992). Because of their anatomical, morphological, reproductive and physiological peculiarities, the aquatic plants attract a special interest from biologists (Perveen, 1999). However, literature dealing with the pollen morphology of aquatic flowering plants is rather scarce.
Many authors have studied the pollen morphology of some species of aquatic families such as Lemnaceae (Aiken, 1978), Potamogetonaceae (Perveen, 1999), Hydrocharitaceae (Takahashi, 1994; Tanaka et al., 2004), Podostemaceae (Obson et al., 2000), Haloragaceae (Labdolt, 1986; Sorsa, 1988), Najadaceae (Shuang-Quan, 2001), Ceratophyllaceae (Takahashi, 1995), Rubbiaceae (Lacroix and Kemp, 1997), Callitrichaceae (Cooper et al., 2000) and Nymphaceae (Shiga and Kadono, 2007).
There are no previous studies of pollen morphology of aquatic plants from Saudi Arabia. The present study aims to study the pollen morphology of six species of aquatic angiosperms, belonging to five genera distributed in five families using both light and scanning electron microscopy.
MATERIALS AND METHODS
Pollen samples were collected from aquatic angiosperms found in different locations of Saudi Arabia (Table 1). The pollen grains were prepared for Light Microscopy (LM) according to Erdtman (1952).
| Table 1: | Aquatic plant species collected from Saudi Arabia for pollen morphological studies |
 | |
The pollen grains were mounted in unstained glycerin jelly and observations were made with a Nikon Type-2 microscope, under E40, 0.65 and oil immersion (E100, 1.25), using a 10x eyepiece. For scanning electron microscopic (SEM) studies, pollen grains were suspended in a drop of water and directly transferred with a fine pipette to a metallic stub using double-sided adhesive tape and coated with gold in a sputtering chamber (Ion-sputter JFC-1100). Coating was restricted to 150A. The SEM examination was carried out on a Jeol microscope JSM-T200. The measurements were based on 15-20 readings from each specimen. Polar Area Index (PAI) was expressed by the ratio of the polar area dimension to the largest transverse dimension of the pollen grains and coefficient of variation (CV) express the standard deviation as a percentage in terms of the arithmetic mean are also given. Polar axis (P), equatorial diameter (E), colpi length, apocolpium, meocolpium, spine length and exine thickness were measured.
The terminology used in this study is in accordance with Erdtman (1952), Kremp (1965), Moore and Webb (1978) and Faegri and Iversen (1992).
RESULTS
Aquatic angiosperms exhibited great diversity in their pollen characters, such as in shape, size, aperture, polarity and tectum type. However, the exine pattern and apertural types are the most significant pollen characters.
The pollen grains of aquatic plant species studied here are mostly apolar or isopolar rarely heteropolar. The shape of pollen grains is commonly spheroidal or oblatespheroidal. Exine sculpturing is also extremely varied, ranging from reticulate, fossulate, foveolate, scarbate, echinate-spinulose or are olate. Apertures are mostly colpate or porate. Non-aperturate pollen is found in Elodea, Potamogeton and Ruppia.
Description of Pollen Grains: Elodea canadensis Rich. (Fig. 1-3). Pollen grains radially symmetrical, isopolar, spheroidal, size: (19.20-) 22.28±0.65 (-30.11) μm in diameter, C.V. 7.92. nonaperturate. Exine (0.40-) 0.83±0.20 (-1.25) μm thick, C.V. 4.01, sexine slightly thicker than nexine. Tectum densely echinate, spines pointed, (4.60-) 5.08±1.03 (-5.34) μm, Lemna gibba L. (Fig. 4). Pollen grains radially symmetrical, apolar, spheroidal, size: (20-) 21.13±0.37 (-22.51) μm in diameter, C.V.3.91.monoporate, pore±circular, c.0.5 μm in diameter. Exine (1.25-) 1.42±0.16 (-1.75) μm thick, C.V. 25.12, sexine thicker than nexine. Tectum spinulose.
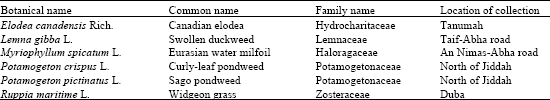
Myriophyllum spicatum L. (Fig. 5-7). Pollen grains are generally radially symmetrical, isopolar, sub-oblate to oblate, 4-zonocolpate, colpi short, elliptic.
 | |
Fig. 1-4: | Pollen grains of Elodea canadensis |
1. | LM photograph showing polar view. The exine covered with densely pointed spines. Scale Bar = 1.0 µm |
2. | LM photograph showing polar view of pollen grain with spines cover the overall pollen surface. Scale Bar = 1.0 µm |
3. | SEM photograph showing the echinate tectum. Scale Bar = 10 µm |
4. | Pollen grain of Lemna gibba. SEM photograph showing the echinate tectum. Scale Bar = 10 µm |
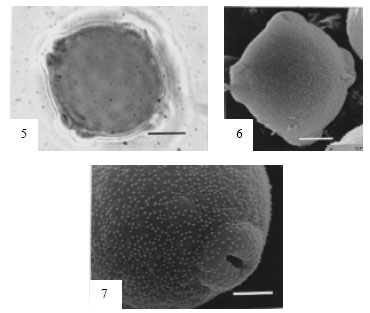 | |
Fig. 5-7: | Pollen grains of Myriophyllum spicatum |
5. | LM photograph showing equatorial view. Note 4 colpi. Scale Bar = 1.0 µm |
6. | SEM photograph showing 4 colpi. Scale Bar = 5.0 µm |
7. | SEM photograph showing magnified part of the pollen grain. Note scabrate-punctate tectum and raised annulus around the colpus. Scale Bar = 10 µm |
 | |
Fig. 8-10: | Pollen grains of Potamogeton crispus |
8. | LM photograph showing polar view. Exine and intine are clearly differentiated. Scale Bar = 1.0 µm |
9. | SEM photograph showing coarsely reticulated tectum. Scale Bar = 5.0 µm |
10. | SEM photograph showing a magnified part of pollen with the pattern of reticulation on tectum. Scale Bar = 5.0 µm |
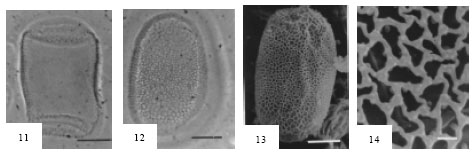 | |
Fig. 11-14: | Pollen grains of Potamogeton pectinatus |
11. | LM photograph showing polar view. Scale Bar = 1.0 µm |
12. | LM photograph showing polar view with reticulated tectum. Scale Bar = 1.0 µm |
13. | SEM photograph showing coarsely reticulated tectum. Scale Bar = 10 µm |
14. | SEM photograph showing a magnified part of pollen indicating the pattern of reticulation on tectum. Scale Bar = 5.0 µm |
 | |
Fig. 15-17: | Pollen grains of Ruppia maritima |
15. | LM photograph showing polar view. Note reticulated exine at the center of the pollen and smoothed exine at the extremities. Scale Bar = 1.0 µm |
16. | LM photograph showing polar view with reticulate and smooth parts of the exine. Scale Bar = 1.0 µm |
17. | SEM photograph showing elongated shape of the pollen and coarsely reticulated tectum. Scale Bar = 10 µm |
Tectum scabrate-punctate. size: Polar axis P(18.59-) 19.54±0.16 (-20.02) μm, CV 3.27 and equatorial diameter E(21.54-) 23.64±0.23 (-24.31) μm, C.V. 3.84. 4-5 colpate, short colpi (1.43-) 2.86±0.13 (-3.95) μm long. Exine 1.43 μm thick, sexine thicker than nexine.
Potamogeton crispus L. (Fig. 8-10). Pollen grains radially symmetrical, apolar, spheroidal, size: (21.25-) 24.28±0.71 (-30.11) μm in diameter, CV 8.93 nonaperturate. Exine (0.50-) 0.83±0.22 (-1.25) μm thick, CV 5.01, sexine slightly thicker than nexine. Tectum coarsely retucilate with irregular pattern of muri, lumina 0.41-2.11 μm in diameter.
Potamogeton pictinatus L. (Fig. 11-14). Pollen grains radially symmetrical, apolar, spheroidal, size: (32.50-) 35.11±1.87 (-37.51) μm in diameter, CV 16.11 nonaperturate. Exine c. 0.12 μm thick, sexine thicker than or as thick as nexine. Tectum coarsely reticulate with irregular pattern of muri, lumina variable in size and shape, 0.22 μm in diameter.
Ruppia maritima L. (Fig. 15-17). Pollen grains radially symmetrical, apolar, elongated (boomerang-shaped), size: (20.24-) 23.26±0.69 (-29.10) μm in diameter, CV 8.91, nonaperaturate. The exine is reticulate in pattern in the central portion of the grain and appears smooth at the knob-like extremities. Exine (0.58-) 0.82±021 (-1.23) μm thick, CV 5.00, sexine slightly thicker than nexine. Tectum coarsely reticulate with irregular pattern of muri, lumina 0.40-2.10 μm in diameter.
DISCUSSION
Based on apertural type, three distinct pollen types are recognized. Pollen Type-I is characterized by its non-aperturate pollen. Four species (Elodea canadensis, Potamogeton crispus, P. pectinatus and Ruppia maritima) have non-aperturate pollen. However, in these species monads are found (Sorsa, 1988; Pettitt and Jermy, 1975; Richardson, 1983). Pollen Type-II is easily recognized by having porate pollen. Pollen Type-III is readily delimited by having colpate pollen. The number of clips varies from mono-pentacolpate. In the present study, aquatic monocots have 3-5-colpate such as Myriophyllum spicatum.
In this study, the most of the aquatic taxa are wind pollinated. There is a correlation between pollen morphology and pollination mechanism (Van Vierssen et al., 1982; Tanaka et al., 2004). A definite relationship is exhibited between pollen characters and pollination types especially in entomophilly and anemophily. Pollen grains of entomophilous taxa are characterized by compound apertures i.e., 3-colporate, prolate-spheroidal shape, generally large, thick walled, sticky and with reticulate tectum, while pollen grains of anemophilous taxa are with simple apertures i.e. monoporate, spheroidal, small, thin walled, dry and with scabrate-areolate tectum (Shuang-Quan et al., 2001).
Underwater pollination systems in angiosperms are derived from aerial floral systems (Philbrick and Les, 1996). Exine reduction or complete exine loss appear to be an essential component in the evolution of hydrophily (Philbrick, 1991). This has been documented in hydrophilous members of Hydrocharitaceae (Pettitt, 1981) such as Elodea canadensis.
In future work, transmission electron microscopy and character state reconstruction of the exine structures and sculptures using a molecular phylogenetic tree are needed. There is a correlation between the variation in the exine and pollination mechanism (Tanaka et al., 2004).
REFERENCES
- Cooper, R.L., J.M. Osborn and C.T. Philbrick, 2000. Comparative pollen morphology and ultrastructure of the callitrichaceae. Am. J. Bot., 87: 161-175.
Direct Link - Lacroix, C.R. and J.R. Kemp, 1997. Developmental morphology of the androecium and gynoecium in Rubbia maritima L.: Considerations for pollination. Aquat. Bot., 59: 253-262.
CrossRef - Obson, J.M., S.P. O`Neill and G. El-Ghazaly, 2000. Pollen morphology and ultrastructure of Marathrum schiedeanum (Podostemaceae). Grana, 39: 221-225.
CrossRef - Perveen, A., 1999. A palynological survey of aquatic flora of Karachi-Pakistan. Tr. J. Bot., 23: 309-317.
Direct Link - Pettitt, J.M., 1981. Reproduction in seagrasses: Pollen development in Thalassia hemprichii, Halophila stipulacea and Thalassodendron ciliatum. Ann. Bot., 48: 609-622.
Direct Link - Philbrick, C.T., 1991. Hydrophily: Phylogenetic and evolutionary considerations. Rhodora, 93: 36-50.
Direct Link - Philbrick, C.T. and D.H. Les, 1996. Evolution of aquatic angiosperm reproductive systems. Bioscience, 46: 813-826.
Direct Link - Shiga, T. and Y. Kadono, 2007. Natural hybridization of the two Nuphars species in Northern Japan: Homoploid hybrid speciation in progress. Aquat. Bot., 86: 123-131.
Direct Link - Shuang-Quan, H., G. You-Hao, G. W. Robert, S. Yao-Hua and S. Kun, 2001. Mechanism of underwater pollination in Najas marina (Najadaceae). Aquat. Bot., 70: 67-78.
Direct Link - Sorsa, P., 1988. Pollen morphology of Potamogeton and Groenlandia (Potamogetonaceae) and its taxonomic significance. Ann. Bot. Fennici., 25: 179-199.
Direct Link - Takahashi, M., 1994. Pollen development in a submerged plant, Ottelia alismoides (L.) Pers. (Hydrocharitaceae). J. Plant Res., 107: 161-164.
CrossRef - Tanaka, N., K. Uehara and J. Murata, 2004. Correlation between pollen morphology and pollination mechanisms in the Hydrocharitaceae. J. Plant Res., 117: 265-276.
CrossRef - Van Vierssen, W., R.J., Van WijkVan and J.R. Der Zee, 1982. On the pollination mechanism of some eurysaline Potamogetonaceae. Aquat. Bot., 14: 339-347.
CrossRef