
Mohamed A. Kandil
Department of Economic Entomology and Pesticide, Faculty of Agriculture, Cairo University, Giza, Egypt
Amal Y. Saleh
Department of Economic Entomology and Pesticide, Faculty of Agriculture, Cairo University, Giza, Egypt
Wafaa H. El Dieb
Central Agricultural Pesticide Laboratory, Agricultural Research Center, Dokki, Giza, Egypt
Sayeda F. Farghaly
Central Agricultural Pesticide Laboratory, Agricultural Research Center, Dokki, Giza, Egypt
Asian Journal of Biological Sciences
Year: 2008 | Volume: 1 | Issue: 1 | Page No.: 33-38







ABSTRACT
An investigation was performed to determine the possible role of metabolism in two resistant strains to thiomethoxam and profenofos by whitefly Bemisia tabaci. Selective synergists were used to study the involvement of hydrolytic or oxidative enzymes in the resistance mechanism of the resistant strains. Resistance level was decreased markedly when DEF (S,S,S-tributyl phosphotithioate) synergized thiamethoxam, suggesting the involvement of increased detoxication by esterases as a part of resistance mechanism. In addition thiamethoxam was synergized moderately when mixed with 25 ppm from TCP (tricresylphosphate) confirming the role of nonspecific esterases in thiamethoxam resistance. Piperonyl Butoxide (PB) synergized profenofos toxicity in resistant strain especially when profenofos mixed with 50 ppm PB which gave synergistic ratio 24.08. The role of diethylmaleate (DM) as inhibitor for glutathione transferases was clear in the case of profenofos resistant strain. However, it exhibited slightly synergistic action in the case of thiomethoxam resistant strain.
PDF Abstract XML References Citation
How to cite this article
Mohamed A. Kandil, Amal Y. Saleh, Wafaa H. El Dieb and Sayeda F. Farghaly, 2008. Resistance Mechanisms of Whitefly Bemisia tabaci (Homoptera: Aleyrodidae) to Thiamethoxam and Profenofos. Asian Journal of Biological Sciences, 1: 33-38.
URL: https://scialert.net/abstract/?doi=ajbs.2008.33.38
URL: https://scialert.net/abstract/?doi=ajbs.2008.33.38
INTRODUCTION
The tobacco whitefly, Bemisia tabaci Gennadius (Homoptera: Aleyrodidae) is a serious pest in many cropping systems throughout the world. B. tabaci is a common pest, particularly in cotton, vegetables and ornamental crops, both directly through feeding and as a vector of numerous plant pathogenic viruses (Byrene et al., 2003). Insecticides have been extensively used for its control and resistance to various insecticides has been reported in different countries and Egypt (Kandil et al., 2006). In Egypt, organophosphates and carbamates have commonly been used in the past and in the 1980 and insect growth regulator, buprofezin, was thought likely to be a useful insecticide for controlling this pest (El-Kady and Devine, 2003). However, it is slow acting and has little effect on the adults and eggs. As an advance on these, imidacloprid was introduced in the early 1990’s and having high activity and long-lasting effect, it has become the primary insecticides for controlling B. tabaci. Similar compounds belonging to this class of insecticide and introduced afterwards, e.g., acetamiprid and thiamethoxam, which exhibit to a great or less extent the same efficacy characteristics and were shown to have an identical mode of action by binding to the same site of action nicotinic acetylcholine receptor (nAChR) (Nauen et al., 2000). Resistance to imidacloprid has been reported in a range of species including silver leaf whitefly (Bemisia argentifollii), western flower thrips (Frankliniella occidentalis), Colorado potato beetle (Leptinotarsa decemlineata), German cockroach (Blattela germanica) and housefly (Musca domestica) (Grafius and Bishop, 1996; Wen and Scott, 1997). Furthermore, a re-testing of (non-selected) laboratory maintained and field (Q-type) strains from Almeria revealed strong cross-resistance to thiamethoxam and acetamiprid as well (Elbert and Nauen, 2000).
PB and DEF are metabolic synergists, so the increases in toxicity that occur when they are co-administered with an insecticide are commonly attributed to their inhibition of important detoxifying enzyme systems (Nauen et al., 2002). Substantial increases in the toxicity of synergized insecticides to R-insects are usually taken as good circumstantial evidence that enhanced metabolism is an important resistance factor (Lee et al., 2006).
In the present study a thiomethoxam and profenofos resistant strains of B. tabaci were selected in the laboratory. Selective synergists were used to elucidate the potential of metabolic mechanisms involved in both resistant strains.
MATERIALS AND METHODS
Insects
A susceptible laboratory strain was collected in 1997 and has remained in culture without any exposure to insecticides. A field strain was collected from Beheira governorate in July 2001. All insect strains were reared on cotton plants in the laboratory as described by Coudriet et al. (1985).
Insecticides and Synergists Used
Insecticides used in this study were: Thiamethoxam (Actara 25% WP), profenofos (Selecron 72% EC). Synergists used in this study were: DEF (S,S,S-tributylphosphorotrithioate), diethylmaleate (DM), Piperonyl-butoxide (PB) and tricresylphosphate (TCP). Test concentrations were prepared by diluting the insecticides in water.
Bioassay
The bioassay method for obtaining concentration-response was described by Prabhaker et al. (1985). Briefly, attached cotton leaves were dipped for 5 sec in 100 mL of the desired concentration of each insecticide. Twenty adult whiteflies were exposed to the treated leaves confined in small cages of polyurethane foam each concentration and replicated five times. Mortality was recorded 24 h after treatment and LC50 values were determined on the bases of standard probit analysis.
A field strain was subjected to laboratory selection pressure with each insecticide i.e., profenofos or thiamthoxam at a level producing 50% mortality to the adult stage. The level of developing resistance was determined at generations 3, 5, 7, 9 and 11 for each resistant strain Resistance Ratio (RR) were determined by dividing the LC50’s of the R-strain by the LC50’s of the S-strain and relative resistance were determined by dividing the LC50’s of the R-strain by the LC50’s of the field-strain. The results of selection and development of resistance are present in Table 1 and 2.
The synergistic of DEF, PB, DM and TCP on resistance were evaluated in both profenfos and thiamethoxam strains because DEF is known to act by inhibiting esterases, PB inhibits the mixed function oxidases DM inhibits glutathione transferases and TCP inhibits non-specific esterases.
| Table 1: | Development of resistance in the whitefly, Bemisia tabaci to profenofos |
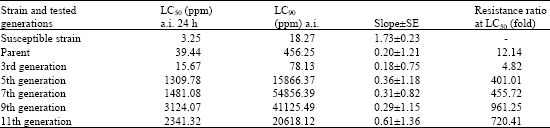 | |
| Resistance ratio = LC50 of R-strain/LC50 of S-strain | |
| Table 2: | Development of resistance in the whitefly, Bemisia tabaci to thiamethoxam |
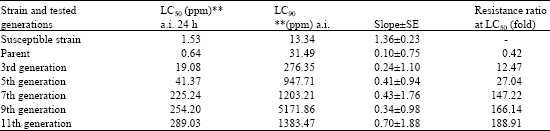 | |
| Resistance ratio = LC50 of R-strain/LC50 of S-strain, **ppm calculated as active ingredient | |
Three levels 50, 25 and 10 ppm from each synergist were used to evaluate their synergistic combination with thiamethoxam or profenofos.
At least six concentrations were tested for each insecticide alone and in combination with synergists. The mortality for treatments was corrected for control mortality by using Abbott’s formula (Abbott, 1925) and the LC50’s were estimated by probit analysis (Finney, 1971). Toxicity of the insecticides + synergist compared with the insecticide alone was expressed as Synergistic Ratio (SR) LC50 of insecticide alone divided by LC50 of insecticide + synergist. The high value of synergistic ratio indicates the presence and the relative importance of detoxication enzymes responsible for a resistance.
RESULTS AND DISCUSSION
Resistance ratios for white fly profenofos-resistant strain as set up in Table 1 (RR = 720.L1) after selection for eleven generations. However, in the case of thiamethoxam the RR was 188.91-fold. This reflects the acceleration of building up resistance to profenofos faster than in the case of thiamethoxam as neonicoticnoid insecticides. In addition this may be due to the intensive used by organophosphate insecticides especially profenofos against whitefly in Egypt. However, thiomethaxam is recently introduced to be one the non-conventional pest control agent against white fly.
Various factors cause resistance in insects, including oxidative degradation by Mixed Function Oxidases (MFOs), hydrolytic degradation by esterases or glutathione transrerances and target site insensitivity (Oppenoorth and Welling, 1976). Detoxication is considered one of the major mechanisms of insecticide resistance (Ishaaya, 2000). Therefore synergists, as inhibitors of metabolic processes, were used to clarify the mechanism of resistance in Bemesia tabaci.
The results in Table 3 and 4 showed the response of profenofos and thiamethoxam whitefly resistant strains to the insecticide alone or to the insecticide + synergists. Data in Table 3 clearly show that piperonyl butoxide PB (mfos inhibitor) enhanced the toxicity of profenofos against R-strain when co-administered with 50 ppm PB which exhibited SR = 24.08 and the level of resistance decreased to ca., 30 fold. In addition diethylmalate DM (glutathione transferases inhibitor) was moderately synergized profenofos (SR ca. 11). This finding indicated major contribution by either mixed function oxidases or glutathione transferases to profenofos resistance in B. tabaci. Segall and Casida (1982) found that profenofos is sterospecifically converted to more potent inhibitor of acetylcholinesterase by mouse liver microsomal mixed function oxidase system. The chiral (-) isomer became a 34 fold better inhibitor of acetylcholiesterase in vitro while the less toxic (+) isomer was deactivated by factor of 2. Recently, Lee et al. (2006) added that pyraclofos, prothiofos, sulphrofos and profenofos are organophosphorus (OP) insecticides exhibited an insecticidal mechanism distinctive compared with that of OP insecticides. It is widely accepted hat O-ethyl, S-n-propyl phosphorothiolaates are activated by oxidation of sulfur atom and their activated forms show very high activity, but are too unstable.
| Table 3: | Synergism of profenofos by DEF, PB, DM and TCP in the adults of Bemisia tabaci profenfos resistant strain |
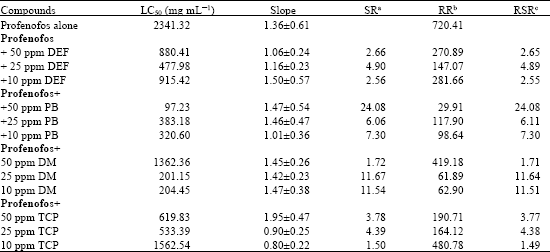 | |
| aSR: Synergist Ratio (LC50 of unsergized/LC50 of synergized treatment), bRR: Resistance Ratio (LC50 of R strain/LC50 of S-strain, cRSR: Relative synergism (RR of unsynergized/RR of synergized treatment) | |
| Table 4: | Synergism of thiamethoxam by DEF, PB, DM and TCP in the adults of Bemisia tabaci thiamethoxam resistant strain |
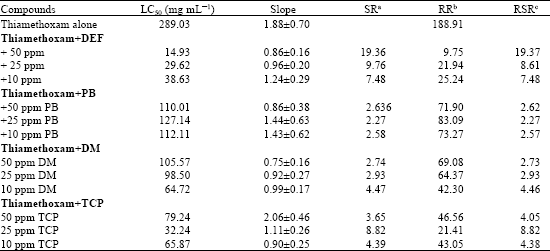 | |
| aSR: Synergist ratio (LC50 of unsergized/LC50 of synergized treatment), bRR: Resistance Ratio (LC50 of R strain/LC50 of S-strain, cRSR: Relative synergism (RR of unsynergized/RR of synergized treatment) | |
However, DEF and TCP (esterases inhibitors) slightly synergized profenofos. This suggests that hydrolytic enzymes not play a significant role in the resistance of whitefly to profenofos.
The above mentioned may be explain why PB (mfo’s inhibitor) and DM (glutathione transferases inhibitor) enhanced the toxicity of profenofos to profenofos resistant strain. Therefore, in this study we can draw attention to the role of mfo and glutathione transeferases as a possible mechanism of resistance of whitefly to profenofos.
In synergism study of thiamethoxam by DEF, PB, DM and TCP the results in Table 4 showed that DEF was the most effective synergist. The toxicity of thiomethoxam to R-strain was increased with SR = 19.36 and resistance ratio decreased to about 9-fold. In addition TCP (nonspecific esterases inhibitor) was moderately synergized thiamethoxam. However, PB and DM have a limited synergism when mixed with thiamethoxam.
As well known thiamethoxam is a second-generation neonicotinoid compound with systemic, stomach and contact activity. It belongs to thianicotinyl subclass that interferes with the nicotinic acetylcholine receptors in the insect’s nervous system. To explore the role of detoxication enzyme in resistant mechanism, Rauch and Nauen (2003) carried out a biochemical studies on the impact of enzymes in neonicotinoid resistance. The results showed that monooxygenase activity increased 2-3 fold in white fly moderately resistant strains (RF approximately 30), even 5-6 fold in highly resistant strain (RF approximately 1000). Only monoxyenase activity correlated with imidacloprid, thiamethoxam and acetamprid resistance and therefore seems to be the enzyme system responsible for neonicotinoid resistance strains from Spain.
On the other hand and in agreement with our results Byrene et al. (2003) demonstrated a lack of any significant NADPH dependent metabolism of imidacloprid in relation to resistance in B-type B. tabaci originating from Guatemala.
Generally Ishaaya (2001) reported that the mechanisms of resistance to insecticides acting outside the nervous system (e.g., insect growth regulators in Bacillus thuringiensis or to more novel neurotoxin (e.g. neonicotinoids) are less understood, but it is likely that they are subjected to detoxification and/or target site alteration.
CONCLUSION
The present study has shown that synergists can be used to identify the role of metabolic enzymes in the mechanism of resistance in whitefly to thiamethoxam and profenofos insecticides. Piperonyl butoxide and diethylmaleate emphasized the role of mixed function oxidases and glutathione transferase as major factors in detoxication mechanism in profenofos resistant strain.
High synergistic ratio obtained from mixing profenofos and DEF, reflects the role of estrases in the detoxication mechanism found in thiamethoxam resistant strain.
REFERENCES
- Abbott, W.S., 1925. A method of computing the effectiveness of an insecticide. J. Econ. Entomol., 18: 265-267.
CrossRefDirect Link - Byrene, F.J., S. Castle, N. Prabhaker and N.C. Toscano, 2003. Biochemical study of resistance to imidacloprid in B-biotype Bemisa tabaci from Guatemala. Pest Manage. Sci., 59: 347-352.
CrossRefPubMedDirect Link - Elbert, A. and R. Nauen, 2000. Resistance of Bemisia tabaci (Homoptera: Aleyrodidae) to insecticides in southern Spain with special reference to neonicotinoides. Pest Manage. Sci., 56: 60-64.
Direct Link - El-Kady, H. and G.J. Devine, 2003. Insecticide resistance in Egyptian populations of the cotton whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae). Pest Manage. Sci., 59: 865-871.
Direct Link - Finney, D.J., 1971. Probit Analysis. 3rd Edn., Cambridge University Press, London, UK., pp: 76-80.
CrossRefDirect Link - Lee, Si Woo, K. Ohta, S. Tashiro and T. Shono, 2006. Metabolic mechanisms of the housefly (Musca domestica) resistant to pyraclofos. Pestic. Biochem. Physiol., 85: 76-83.
CrossRefDirect Link - Nauen, R., N. Stumpf and A. Elbert, 2002. Toxicological and mechanistic studies on neonicotinoid cross resistance in Q-type Bemisia tabaci (Hemiptera: Aleyrodidae). Pest Mange. Sci., 58: 868-875.
PubMedDirect Link - Prabhaker, N., D.L. Coudriet and D.E. Meyerdirk, 1985. Insecticide resistance in the sweetpotato whitefly, Bemisia tabaci (Homoptera: Aleyrodidae). J. Econ. Enomol., 78: 748-752.
CrossRefDirect Link - Rauch, N. and R. Nauen, 2003. Identification of biochemical markers linked to neonicotinoid cross resistance in Bemisia tabaci (Hemiptera: Aleyrodidae). Arch. Insect Biochem. Physiol., 54: 165-176.
CrossRefPubMedDirect Link - Wen, Z. and J.G. Scott, 1997. Cross-resistance to imidacloprid in strains of German cockroach (Blatella germanica) and housefly (Musca domestica). Pestic. Sci., 49: 367-371.
CrossRef