
P. Lakshmi
Department of Botany, Andhra University, Visakhapatnam, 530 003 Andhra Pradesh, India
P. Akbar Ali Khan
Department of Pharmaceutical Sciences, Andhra University, Visakhapatnam, 530 003 Andhra Pradesh, India
P. Narasimha Reddy
Department of Pharmaceutical Sciences, Andhra University, Visakhapatnam, 530 003 Andhra Pradesh, India
K. Lakshminarayana
Department of Botany, Andhra University, Visakhapatnam, 530 003 Andhra Pradesh, India
S. Ganapaty
Department of Pharmaceutical Sciences, Andhra University, Visakhapatnam, 530 003 Andhra Pradesh, India
Asian Journal of Biological Sciences
Year: 2008 | Volume: 1 | Issue: 1 | Page No.: 1-10







ABSTRACT
In the present study an attempt has been made to assess the genotypic variability among twelve species of the genus Tephrosia, distributed in Andhra Pradesh, through DNA fingerprinting using RAPD technique. Twenty OPC (Operon Biotechnologies GmbH, Germany) primers were used. The cluster analysis based on the similarity matrix was performed using the Unweighted Pair Group Method with Arithmetic Average (UPGMA) with the help of PHYLIP software ver. 3.65 pooled from all the six primers. Present study has justified to a great extent co-relating with the classification based on the morphological traits. However a distinction between some members of the genus Tephrosia is still a matter of debate. Hence further analyses are needed to determine the correct intrageneric taxonomic treatment of Tephrosia, since it represents one of the largest and most complex groups in the core tribe Millettieae of the family Fabaceae. This study represents the first approach in using nuclear DNA finger print markers as a tool to study molecular systematics of the genus Tephrosia.
PDF Abstract XML References Citation
How to cite this article
P. Lakshmi, P. Akbar Ali Khan, P. Narasimha Reddy, K. Lakshminarayana and S. Ganapaty, 2008. Genetic Relationship Among Tephrosia Species as Revealed by RAPD Analysis. Asian Journal of Biological Sciences, 1: 1-10.
URL: https://scialert.net/abstract/?doi=ajbs.2008.1.10
URL: https://scialert.net/abstract/?doi=ajbs.2008.1.10
INTRODUCTION
The genus Tephrosia is a pantropical taxa with about 400 species (Geesink, 1981) distributed chiefly in Asia, Africa, Australia and America. Tephrosia belongs to tribe Millettieae (formerly Tephrosieae), of the family Fabaceae (Leguminosae) and has earlier been classified by many taxonomists either into sections or subgenera, based mainly on the morphological traits. De Candolle (1825) classified Tephrosia into four sections namely Mundulea, Brissonia, Craccoides and Reineria. Of these only Mundulea and Reineria were represented in India. Later Bentham (1865) and Baker (1876) have classified the genus into three subgenera as Macronyx, which includes T. tenuis,Brissonia that includes T. candida and Reineria, which includes rest of the species of Tephrosia.
Wood (1949) categorized the new world species of Tephrosia into two groups, one with a glabrous style and the other with a pubescent style. Gillett (1971) adopted this classification in African species of Tephrosia. Subsequently Brummitt (1981) divided Tephrosia into two subgenera, subg. Tephrosia with a glabrous style (according to which T. hamiltonii, T. pumila, T. purpurea, T. spinosa, T. strigosa, T. villosa etc were included) and subg. Barbistyla with a trichiferous style (which includes T. candida, T. maxima, T. pulcherrima, T. tinctoria etc.). The above classification was criticized since some of the taxa (T. pumila,T. maxima etc.) showed both glabrous and trichiferous styles.
The general systematic framework of the genus Tephrosia has been focused primarily from morphological data, which is adequate and does probably reflect grouping of a phyletic nature. But there are species, whose relationships are difficult to judge, even after comprehensive morphological study. Phytochemical studies especially flavonoids might prove critical for such taximetric problems, provided the biosynthetic pathways of such phytoconstituents is known. Therefore, the data in themselves might be particularly impressive in taxonomic point of view by further understanding of the genus through genetic characterization using molecular markers. As such biochemical data and morphological data pooled together with molecular data would be helpful in sorting out the interrelationships between the various species of Tephrosia distributed in a particular geographical area. Despite, years of extensive research on the genus Tephrosia, very little is known about its phylogeny. Raina et al. (1985) studied the species variation cytogenetically (based on G+C Content) in eight species of Tephrosia. Untill to-date, no work using molecular data (DNA fingerprinting technique) has been carried out in evaluating the infrageneric relationships among the various species of the genus Tephrosia, the only exception being the usage of the molecular data, while evaluating the tribe Millettieae (Acharya et al., 2004).
For a better understanding of the systematic relationships within the genus Tephrosia, RAPD markers were used to study the variation among twelve taxa distributed in the state Andhra Pradesh, India. Random amplified polymorphic DNA dubbed RAPD, has become a powerful tool for finger printing a variety of organisms; of bacteria, plants, animals and humans (Williams et al., 1990). RAPD`s arise by PCR with short oligonucleotides of arbitrary sequences that prime DNA synthesis from genomic sites, which they fortuitously match or almost match. The high-density genetic maps constructed through RAPD`s have closely spaced DNA markers, which are very useful for genome analysis (Williams et al., 1993). RAPD analysis in particular has found widespread application in systematics since a large number of markers can be quickly generated and scored (Rieseberg and Ellstrand, 1993). They have been successfully applied to assess genomic variability in plant species like Fragaria spp. (Thongthieng and Prasartporn, 2003), Orobanche spp. (Roman et al., 2003), Leucaena spp. (Hawkins and Harris, 1998), Oxytropis spp. (Janet Jorgensen et al., 2003), Afgekia spp. (Prathepha and Baimai, 2003) etc.
The RAPD fingerprinting technique using decanucleotide primers has presently been used to reevaluate the systematic status of the genus Tephrosia Pers, which represents one of the best examples of the complexities, where the species definitions and relationships within the genus were unclear. Further to analyze the affinities and phylogenetic relationships of the various species of the genus Tephrosia and also to trace the progenitor- Monophyletic/polyphyletic origin of the species.
MATERIALS AND METHODS
Plant Material
Twelve species of the genus Tephrosia have been collected from various regions of Andhra Pradesh State,India and two species, Crotalaria verrucosa (Fabaceae), Sorghum bicolor (Gramineae) were chosen as out groups (Table 1). All these specimens (Fig. 1) were collected and identified with the help of recent floras (Gamble and Fischer, 1918; Pullaiah and Chennaiah, 1997) and herbarium specimens available in the Department of Botany, Andhra University. Further all the above plant species were authenticated by Prof. T. Pulliah, Taxonomist, Department of Botany, Sri Krishnadevaraya University (SKU), Anantapur, India. All the voucher specimens were deposited in the Herbarium, Depatment of Botany, Andhra University, Visakhapatnam, India in April 2005.
DNA Extraction, Amplification and Electrophoresis
DNA was extracted from fresh/dried leaves of the various species of Tephrosia as described in Reineke et al. (1998) with little modifications. DNA amplification was done using decamer (10 nucleotide length) with a G+C content of above 50% as that they form strong hybrids with target DNA and to withstand 72°C of polymerase reaction. Twenty OPC (Operon Biotechnologies GmbH, Germany) primers were used.
 | |
| Fig. 1: | The various Tephrosia species collected from different regions of Andhra Pradesh, India |
| Table 1: | Plant species collected from the different areas of Andhra Pradesh authenticated and voucher specimens were deposited in the Herbarium, Department of Botany, Andhra University, Visakhapatnam |
 | |
| Table 2: | Primers code, sequence, G+C content, type and number of bands amplified in the RAPD analysis of the genus Tephrosia |
 | |
The nucleotide sequence of these primers is shown in the Table 2. Of these twenty primers, six primers were chosen that gave a reasonable number of polymorphic bands, which were scorable and consistent among the duplicates of reactions. Control reaction without genomic DNA was run for all these primer reactions. Duplicate reactions were run with DNA extracted from independent DNA extractions for each isolate.
The RAPD analysis was performed as per the standard methods of Williams et al. (1990). Each amplification reaction mixture of 25μL consisted of 10 μL of DNA (20 ng), 2.5 μL of 10X assay buffer (100 mM Tris-HCl, pH 8.3, 0.5 M KCl and 0.01% gelatin, 1.5 mM MgCl2), 0.5 μL of DNTP mix, 1 μL of primer (20 pm) and 1 μL of 1 U Taq DNA polymerase (Bangalore Genei Pvt.Ltd., Bangalore, India) and 10 μL of sterile double distilled water. Amplification reactions were performed using a thermocycler-Eppendorf master cycler gradient (Eppendorf Netheler-Hinz, GMBH, Hamburg, Germany), programmed as: initial denaturation for 5 min at 95°C, followed by 45 cycles of denaturation for 1 min at 95°C, annealing for 1 min at 37°C and extention for 2 min at 72°C and a final extension for 7 min at 72°C using the fastest available temperature transitions. A control reaction without DNA was setup for every set of reactions.
Amplified DNA fragments along with a size marker Lambda DNA/Hind III digest, (Bangalore Genei Pvt.Ltd., Bangalore, India) for band size comparison, were resolved electrophoretically on 1.5% Agarose gel stained with ethidium bromide (10 mg mL-1) at 50 V/cm for 5 h using 1X TAE buffer and photographed under UV illumination with a Kodak DC 120 digital camera mounted on a dark hood. The photographs were transferred to a PC using Kodak digital transfer software. The gels were scored conservatively. The reproducibility of all initially scored bands were rechecked by comparing banding patterns of individual plants that were rerun to check the cross comparability among gels.
Statistical Analysis
The banding patterns obtained from the RAPD fingerprinting were analyzed to estimate the genetic relationship among the twelve species of the genus Tephrosia. A 0/1 data matrix was constructed for each primer by scoring each species for the presence (1) or absence (0) of each band. To minimize errors RAPD fingerprints were evaluated four times with time intervals by different individuals. The less intense high molecular weight minor bands (which may be a result of primer annealing to template regions with fortuitous matching) were marked separately.
To account for the dominant nature of RAPD markers, allele frequencies were estimated using two different methods (Krauss, 2000), a Bayesian method with uniform prior distribution of allele frequencies (Zhivotovsky, 1999) and the square root method of the null homozygote frequency (Nei and Li, 1979). Calculations were performed using the AFLP-SURV 1.0 software (Vekemans et al., 2002). To examine relationships among species of Tephrosia, a cluster analysis was conducted based on the similarity matrix between each pair of individuals in the dataset using the Unweighted Pair Group Method with Arithmetic average UPGMA (Nei and Li, 1979) with the help of PHYLIP software ver. 3.65 (Felsenstein, 1989). The tree thus generated was explored using the programme Tree Explorer of MEGA version 3.0 (Kumar et al., 2004).
RESULTS
A dendrogram (Fig. 3) was constructed from the similarity values obtained in pair wise comparisons among the species using data, pooled from all the six primers. Cluster analysis revealed rooted groups with a similarity of 25% among the various species of the genus Tephrosia. Out of the twenty primers tested, six primers gave reproducible, clear polymorphic bands (Fig. 2). A total of 172 Polymorphic bands were identified with an average of 27 bands per primer. The size of the amplified fragments ranged from 560 kb to 200 bp (Table 2). Less intense bands which were considered as minor bands, the bands which were shared by not more than four isolates were considered as rare and those bands which were represented alone were considered as unique. The dendrogram constructed excluding minor bands did not differ much from the cluster diagram that was constructed, including both the major and minor bands (Fig. 3). The dendrogram was constructed considering all bands which resulted into five groups with an overall similarity of just 24% among all the sixteen OTU`s (Operational Taxonomic Units). Group II was the largest with six OTU`s, which was further grouped into IIA and IIB with a similarity of 35%. The outgroups Sorghum bicolor and Crotalaria verrucosa were clustered with Tephrosia strigosa with a similarity of 5%.
Group 1: It includes two OTU`s T. purpurea and T. spinosa with a similarity of 20%.
Group 2: It is the largest of all with a similarity of 35% including six OTU`s which were further grouped into two sub-groups designated as Group 2A and Group 2B. Group 2A includes T. pumila, T. villosa and T. hirta and Group IIB includes T. pulcherima, T. tinctoria and T. maxima.
Group 3: It includes two OTU`s, T. procumbens and T. calophyla with a 25% similarity.
Group 4: This group includes only one species T. hamiltonii, which showed much diversity with the rest of the species.
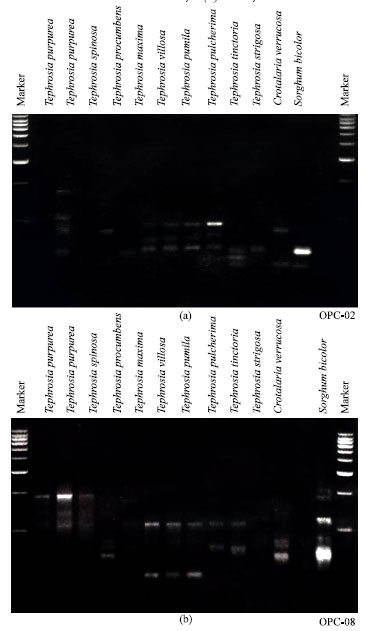 | |
| Fig. 2: | Ethidium bromide stained agarose gels showing RAPD-PCR Banding patterns of the genomic DNA of various species of Tephrosia with primers (a) OPC-02 and (b) OPC-08 (Operon Technologies Inc. Almeda, CA) |
DISCUSSION
De Candolle (1825) classified Tephrosia into four sections, Mundulea Brissonia, Craccoides and Reineria. Of these only two sections Mundulea and Reineria were represented in India. In the present work, of the 12 taxa studied, seven were assignable to the De Candolle`s system (1825), while others were new inclusions. T. spinosa, T. tinctoria, T. maxima, T. pumila, T. villosa, T. hirta and T. purpurea that belongs to the Reineria section of De Candolle`s classification, were also justified with the present RAPD fingerprinting analysis.
Subsequently, Brummitt (1981) categorized Tephrosia into two subgenera, subgenus Tephrosia with a glabrous style and subgenus Barbistyla with a trichiferous style. According to this classification, T. hamiltonii, T. pumila, T. purpurea, T. spinosa, T. strigosa and T. villosa of the present study were included in the sub-genus Tephrosia, while T. tinctoria, T. maxima and T. pulcherrima belong to the subgenus Barbistyla.
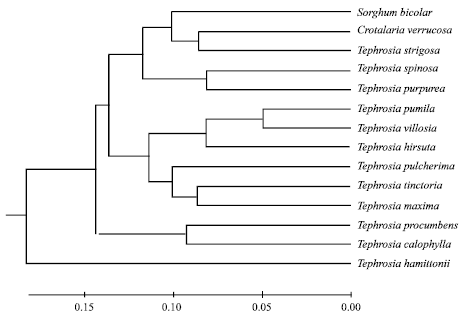 | |
| Fig. 3: | Dendrogram showing the genotypic relationship among the species of the genus Tephrosia |
The present study has much deviated from the above classification, since only T. pumila and T. villosa were grouped together, but were clustered along with T. tinctoria, T. maxima, T. pulcherrima as a major Group 2 and more over the species T. maxima and T. pulcherrima showed the presence of both glabrous and trichiferous style. Therefore, the results obtained through RAPD fingerprinting are quiet contrasting with Brummitt`s classification.
Recently Subba Rao and Shanmukha Rao (1993, 1995) classified the genus Tephrosia on the basis of differential distribution of the non-protein amino acids and numerical taxonomy (based on epidermis, leaf architecture, seed morphology and chemotaxonomy) covering 18 taxa distributed in the South Indian regions. Of the 18 species, 9 species were included in our present study and the rest of the two species T. procumbens and T. hirta are of new inclusion. According to the numerical taxonomy, the genus Tephrosia was clustered into 11 groups that showed no clear demarcation and indication of interrelationship among the various species. The RAPD analysis supported the above classification in the separation of T. purpurea from T. hamiltonii and inclusion of T. maxima and T. pumila in the same group.
Relation of the RAPD Fingerprints to the Morphological Traits
Tephrosia strigosa which was earlier treated separately (in macronyx) by Bentham (1865) and Baker (1876) on the basis of its simple leaf, in contrast to the typical compound leaves of Tephrosia, was separated along with the out groups Sorghum (Gramineae) and Crotalaria (Fabaceae) in the dendrogram (Fig. 3) justifying its diversification from the rest of the taxa. Thus the RAPD fingerprinting relationships co-related with the morphological classification.
Group 1: It includes T. spinosa and T. purpurea, the present clustering was in agreement with morphological traits, since both the species are mostly morphologically similar except for the presence of stipulous spines, flower colour and pod characters of T. spinosa.
Group 2A: This cluster includes T. pumila, T. villosa and T. hirta. The species T. villosa and T. hirta which were treated earlier as synonyms in different floras (Gamble and Fischer, 1918; Pullaiah and Chenniah, 1997) has showed only 75% similarity.
However, further evidence is required, since RAPD fingerprinting alone cannot justify the treatment of both the species as distinct., T. villosa and T. pumila clustered in RAPD dendrogram was in agreement with morphological traits since both of them share many morphological characters (leaf-let size, flower and pod characters).
Group 2B: It includes T. pulcherrima, T. tinctoria and T. maxima. RAPD dendrogram is slightly in contrast with morphological traits, since T. tinctoria and T. maxima, which were clustered together showed differences in leaflet size and flower colour and the T. tinctoria showed more morphological similarity with T. pulcherrima when compared to T. maxima.
Group 3: It includes T. procumbens and T. calophylla, both of which show quiet contrasting morphological characters. T. procumbens is a creeper with compound leaves and white flowers. T. calophylla is endemic to South India and was included in the sub-genus Reineria by Baker (1876). This taxon is found to possess a number of unique characters namely woody herb, simple leaves, tuberous root, brick red flowers etc. Therefore the RAPD fingerprinting is not in agreement with morphological traits in the above grouping.
Group 4:
It includes only a single species T. hamiltonii. The RAPD dendrogram in this case is in agreement with numerical and chemotaxonomic evidence (Subba Rao and Shanmukha Rao, 1990, 1995) where T. hamiltonii is shown to be much diverged from T. purpurea, though they morphologically reassembled each other. This was further strengthened by the isolation of a new flavonoid flemichapparin-B from T. hamiltonii, that which was not reported in T. purpurea (Rajani and Sarma, 1988). Therefore the present evidence from the RAPD cluster analysis also supports the isolation of T. hamiltonii not only from T. purpurea but also from the rest of Tephrosia studied; as such T. hamiltonii represents the lone constituent of a separate Group 4.
CONCLUSIONS
The inter-relationship between the various species of the genus Tephrosia has been a matter of debate. Some of the species, which are traditionally considered to be closely related, showed much diversification in chemotaxonomic and numerical taxonomic evaluation (T. pulcherrima and T. tinctoria, T. villosa and T. pumila etc.). Our present study has justified to a great extent co-relating with classification based on the morphological traits, however a distinction between some members of the genus Tephrosia (T. calophylla and T. procumbens, T. maxima and T. tinctoria etc.) is still a difficult task. As such, further analyses are needed to determine the correct infrageneric taxonomic treatment of the genus Tephrosia, since it represents one of the largest and most complex groups in the core tribe Millettieae of the family Fabaceae.
This study represents the first approach in using nuclear DNA finger print markers as a tool to study molecular systematics in the genus Tephrosia. Pers. The analysis of additional species and the use of different nuclear molecular markers, such as ITS (Internal Transcribed Spacers) or plastid/ mitochondrial DNA (RbCL, trn K, mat K etc.,) sequences will improve the accuracy of resolution of genetic relationships and contribute to a more accurate classification of the genus Tephrosia.
ACKNOWLEDGMENTS
One of the authors (L P) is thankful to University Grants Commission (UGC), New Delhi, for a fellowship (SRF/ No. F.17-116/98(SA-I).
REFERENCES
- Acharya,L., A.K.Mukherjee and P.C Panda, 2004. Genome relationship among nine species of Millettieae (Leguminosae: Papilionoideae) based on Random Amplified Polymorphic DNA (RAPD). Z. Naturforsch., 59: 868-873.
Direct Link - Brummitt, R.K., 1981. Reconsideration of the genera ptycholobium, Caulocarpus, Lupinophyllum and Requienia in relation to Tephrosia (Leguminosae: Papilionoideae). Kew. Bull., 35: 459-473.
CrossRefDirect Link - Felsenstein, J., 1989. PHYLIP-phylogeny inference package (Version 3.2). Cladistics., 5: 164-166.
Direct Link - Hawkins, J.A. and S.A. Harris, 1998. RAPD characterisation of two neotropical hybrid legumes. Plant. Syst. Evol., 213: 43-55.
CrossRefDirect Link - Janet Jorgensen, L., S. Ivana, B. Christian and C. Elena, 2003. Implications of ITS sequences and RAPD markers for the taxonomy and biogeography of the Oxytropis campestris and O. arctica (Fabaceae) complexes in Alaska. Am. J. Bot., 90: 1470-1480.
Direct Link - Krauss, S.L., 2000. Accurate gene diversity estimates from amplified fragment length. Mol. Ecol., 9: 1241-1245.
Direct Link - Kumar, S., K. Tamura and M. Nei, 2004. MEGA3: Integrated software for molecular evolutionary genetics analysis and sequence alignment. Brief. Bioinform., 5: 150-163.
CrossRefPubMedDirect Link - Nei, M. and W.H. Li, 1979. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA., 76: 5269-5273.
CrossRefPubMedDirect Link - Prathepha, P. and V. Baimai, 2003. Molecular characterization of the divergence of rare species of the genus Afgekia (Papilionoideae, tribe Tephrosieae) by RAPD markers and nucleotide sequence analysis. Sci. Asia, 29: 13-20.
Direct Link - Raina, S.N., P.K. Srivastav and S.R. Rao, 1986. Nuclear DNA variation in tephrosia. Genetica, 69: 27-33.
CrossRef - Reineke, A., P. Karlovsky and C.P.W. Zebitz, 1998. Preparation and purification of DNA from insects for AFLP analysis. Insect Mol. Biol., 7: 95-99.
CrossRefDirect Link - Rieseberg, L.H. and N.C. Ellstrand, 1993. What can molecular and morphological markers tell us about plant hybridization? Crit. Rev. Plant Sci., 12: 213-241.
CrossRefDirect Link - Roman, B., C. Alfaro, A.M. Torres, M.T. Moreno, Z. Satovic, A. Pujadas and D. Rubiales, 2003. Genetic relationships among Orobanche species as revealed by RAPD analysis. Ann. Bot., 91: 637-642.
CrossRefDirect Link - Rao, S.J.V. and S.S.R. Rao, 1990. Chemotaxonomic delimitation of tephrosia hamiltonii drumm. ex gamble and T. purpurea (L.) pers. Ind. J. Bot., 13: 36-39.
Direct Link - Rao, J.V.S. and S.R.S. Rao, 1993. Phenolic compounds in the taxonomy of Tephrosia Pers. (Leguminosae). Feddes Repert, 104: 245-250.
Direct Link - Subba Rao, J.V. and S.R.S. Rao, 1995. Numerical taxonomy of some Indian Tephrosia Pers.(Fabaceae). J. Indian Bot. Soc., 74: 149-153.
Direct Link - Thongthieng, T. and P. Smitamana, 2003. Genetic relationship in strawberry cultivars and their progenies analyzed by isozyme and RAPD. ScienceAsia, 29: 1-5.
Direct Link - Vekemans, X., Beauwens, M. Lemaire and I. Roldan-Ruiz, 2002. Data from amplified fragment length polymorphism (AFLP) markers show indication of size. Mol. Ecol., 11: 139-151.
Direct Link - Williams, J.G.K., A.R. Kubelik, K.J. Livak, J.A. Rafalski and S.V. Tingey, 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res., 18: 6531-6535.
CrossRefPubMedDirect Link - Williams, J.G.K., M.K. Hanafey, J.A. Rafalski and S.V. Tingey, 1993. Genetic analysis using random amplified polymorphic DNA markers. Methods Enzymol., 218: 704-740.
Direct Link - Wood, Jr., C.E., 1949. The American barbistyled species of Tephrosia (Leguminosae). Rhodora., 51: 193-384.
Direct Link - Zhivotovsky, L.A., 1999. Estimating population structure in diploids with multilocus dominant DNA markers. Mol. Ecol., 8: 907-913.
Direct Link
Dayo Reply
Good work....it helps me in my project and seminar...thanks for such a good work..that helps me well.