
Ahmad Alsayed Omar
Department of Biochemistry, College of Agriculture, Zagazig University, Zagazig 44511, Egypt
LiveDNA: 20.835
Azza Hosni Mohamed
Department of Biochemistry, College of Agriculture, Mansoura University, Mansoura, Egypt
LiveDNA: 20.11836
Mahmoud Imam Nasr
Department of Molecular Biology, Genetic Engineering and Biotechnology Research Institute (GEBRI), University of Sadat, Egypt
Khalil Abdel Hameed El-Halafawy
Department of Molecular Biology, Genetic Engineering and Biotechnology Research Institute (GEBRI), University of Sadat, Egypt
Elsayed Abdel Khalek Alabsawy
Department of Molecular Biology, Genetic Engineering and Biotechnology Research Institute (GEBRI), University of Sadat, Egypt
Ahmed Ismail Hamdi
Department of Food legume Research, Field Crops Research Institute (FCRI), Agricultural Research Center (ARC), Egypt
Ehab Mohamed Zayed
Department of Cell Research, Field Crops Research Institute (FCRI), Agricultural Research Center (ARC), Egypt
American Journal of Biochemistry and Molecular Biology
Year: 2018 | Volume: 8 | Issue: 1 | Page No.: 34-47







ABSTRACT
Background and Objective: Molecular analysis of sources of resistance to plant pathogens should expedite and confirm novel gene discovery and consequently the development of disease resistant cultivars. By using parents that produce segregating populations with a large genetic variance, breeders could predict which biparental populations will have the maximum genetic variance using the coefficient of parentage or genetic distance estimates based on molecular markers data. To identify molecular markers selection to help soybean breeders in breeding programs and, to study genetic distance among the tested soybean genotypes. Materials and Methods: DNA markers were used as a molecular characterization of genetic improvement of soybean against cotton leaf warm. Four molecular analysis techniques were used in this study to differentiate among 14 genotypes of soybean. First, isozyme variation using native gel electrophoresis (Native-PAGE); secondly, Sodium Dodecyl Sulfate Polyacrylamide gel electrophoresis (SDS-PAGE) protein electrophoresis for leaves and seeds proteins characterization; thirdly, Random Amplified Polymorphic DNA (RAPD) technique using 10 mer arbitrary primers and fourthly, Restriction Fragment Length Polymorphism (RFLP) using BamHI, MspI and TaqI enzymes. Results: No clear trend has been observed in isozymes analysis regarding differentiation among 14 soybean genotypes based on their resistance to cotton leaf worm. SDS-PAGE for the water-soluble seed proteins in the soybean genotypes exposed a total number of 13 bands most of them are common for the 14 genotypes except two bands with molecular weight 220.36 and 194 KDa for that are specific for Calland genotype (moderately susceptible). Restriction site analysis of the ITS region amplified by PCR using universal primers pair was able to successfully amplify the Internal Transcribed Spacer (ITS) region of all genotypes tested. The RAPD analysis showed that primer A9B7 and A1A13 have 100% polymorphic, however primer A7 and A1 have 0 and 58% polymorphic, respectively. Conclusion: The tolerance was correlated with band density as well as with genotypes, where Forrest, Giza 111, Giza 35, Calland, Clark and Giza 82 were the most related genotypes for tolerant/moderately tolerant against cotton leaf warm. The obtained results could be used as molecular marker selection to help soybean breeders in breeding programs and also, to study genetic distance among the tested soybean genotypes.
PDF Abstract XML References Citation
Received: June 13, 2018;
Accepted: August 21, 2018;
Published: November 06, 2018
How to cite this article
Ahmad Alsayed Omar, Azza Hosni Mohamed, Mahmoud Imam Nasr, Khalil Abdel Hameed El-Halafawy, Elsayed Abdel Khalek Alabsawy, Ahmed Ismail Hamdi and Ehab Mohamed Zayed, 2018. Molecular Characterization of Soybean (Glycine max L. Merr) Genotypes Tolerant and/or Susceptible to Cotton Leaf Worm (Spodoptera littoralis). American Journal of Biochemistry and Molecular Biology, 8: 34-47.
DOI: 10.3923/ajbmb.2018.34.47
URL: https://scialert.net/abstract/?doi=ajbmb.2018.34.47
DOI: 10.3923/ajbmb.2018.34.47
URL: https://scialert.net/abstract/?doi=ajbmb.2018.34.47
INTRODUCTION
Soybean (Glycine max Merr.) is one of the important food and industrial crops in many regions of the world, because it is containing about 30% of cholesterol free oil, about 40% of protein which is similar in its nutritious value to the animal protein and beneficiary secondary metabolites such as isoflavones, phenolic compounds and saponins1. Soybean is generally susceptible to nematodes and insects. Thus, Soybean producers suffer substantial annual economic losses due to the damage caused by these invaders. The Cotton Leaf Worm (Spodoptera littoralis) is a serious pest of a large variety of crops in many parts of the world. Larvae of this pest can host on more than 80 economically important plant crops belonging to 40 families such as cotton, soybeans and other vegetables1.
Several environmental manipulations can be attained by employing several control measures like the use of chemical insecticides and cultural and physical control methods. Chemical pesticides are effectively used against insect pests however they are associated with several disadvantages including high costs and concerns about environmental pollution and food safety2. Moreover, the intensive use of broad spectrum insecticides against S. littoralis has led to the development of insect resistance to many registered pesticides3. Mohamed et al.4 studied the effect of the two elicitors methyl jasmonate (20 μM) and sodium nitroprusside (500 μM) on six soybean genotypes and to enhance the ability of susceptible genotypes to resist cotton leaf worm, treatment with methyl jasmonate was found to be more effective than sodium nitroprusside and enhanced the resistance of the susceptible genotypes.
Plant resistance to these insects can contribute to integrated pest management. To develop soybean cultivars with insect resistance, tolerant germplasms should be identified and used as resistance donor parents. Rapid progress in molecular biology and genomics have greatly improved the reliability and usefulness of a variety of DNA-based molecular markers such as random amplified polymorphic DNA (RAPD), simple sequence repeats (SSRs), amplified fragment length polymorphisms (AFLPs) and single nucleotide polymorphisms (SNPs). These DNA-based markers have many advantages over morphological and isozyme-based biochemical markers, particularly in the dramatically increased saturation of genome coverage provides much detailed information on the nature and extent of genetic variation within and between species5. Unlike molecular markers that rely on a DNA assay, morphological markers that rely on visible traits and biochemical markers based on proteins produced by genes. They may differ in a variety of ways such as their technical requirements (e.g., whether they can be automated or require use of radioactivity); the amount of time, cost and labor needed; the number of genetic markers that can be detected throughout the genome; and the amount of genetic variation found at each marker in a given population6.
Research has identified several genes associated with insect resistance. With the development of molecular markers, there is a potential to facilitate the identification, isolation and transfer gene(s) responsible for the pest resistance. Isozymes are a class of proteins called enzymes, which act within the cell to change bio-molecules from one form to another, to produce energy and/or for structural purposes. An organism's DNA governs the synthesis of proteins from amino acids. Sometimes a change in the DNA code of an enzyme would change the surface charge of the enzyme molecule without affecting functionality. These charge isomers of an enzyme are called isozymes and they are used to assay genetic variation. Molecular markers should not be considered as normal genes, as they usually do not have any biological effect and instead can be thought of as constant landmarks in the genome. They are identifiable DNA sequences, found at specific locations of the genome and transmitted by the standard laws of inheritance from one generation to the next7.
Plant breeders would like to predict which biparental populations will have the largest genetic variance8. If the population genetic variance could be predicted, using the coefficient of parentage or genetic distance estimates based on molecular markers data8. Breeders could choose parents that produce segregating populations with a large genetic variance. The wild soybean differed remarkably from the cultivated ones in allele frequency9. The selection and breeding for yield during the past 60 years had no major influence on the protein composition, because of the limitation in the genetic diversity among the parental lines10,11.
The information provided by molecular markers to the breeders will vary depending on the type of marker system used. Each one has its advantages and disadvantages. RFLPs were the first molecular markers to be widely used. Their use is, however, time-consuming and expensive and simpler marker systems have subsequently been developed12,13. RFLPs are markers detected by treating DNA with restriction enzymes (RE)14. Differences in the lengths of DNA fragments will then be seen if the specific site for the RE has been found at different places in the genome (e.g., tip of a certain chromosome), whereas another individual has the sequence, which is not cut by the specific RE. RAPD markers were first described by Williams et al.15. They are detected using the polymerase chain reaction (PCR). The analysis for RAPD markers is quick and simple, although results are sensitive to laboratory conditions16,17. RAPD-PCR was used to analyze the genetic diversity of the 14 studied soybean genotypes and to assess their genetic relationships using similarity indices and dendrogram tree. Four arbitrary random primers were used to determine RAPD polymorphism among soybean genotypes.
This work is aimed to study the molecular characterization of fourteen soybean genotypes based on their tolerant and/or susceptible to cotton leaf worm using different molecular markers techniques to identify molecular marker selection to help soybean breeders in breeding programs and also, to study genetic distance among the tested soybean genotypes.
MATERIALS AND METHODS
Plant material: Fourteen exotic and Egyptian soybean genotypes (Glycine max L. Merr.) from the germplasm collection of Soybean Breeding Program at Agriculture Research Center (ARC), Ministry of Agriculture, Giza, Egypt were used in this study. This study was conducted from September, 2016 to December, 2017. Seeds from each genotype were selected for uniformity by choosing those of equal size and with the same color for germination. The genotypes included tolerant and susceptible genotypes as indicated in Table 1.
Native-PAGE leaf isozymes: Profile of three enzymes peroxidase (PRX), esterase (EST) and malate dehydrogenase (MDH) were compared among the tested genotypes. Dry leave sections from seeds that were just completed germination were pulverized in liquid N2 (as this was found to aid enzyme extraction) in a mortar and pestle. About 500 μL of extraction buffer (0.1 M sodium citrate, 0.2 M sodium phosphate buffer, pH 7) was mixed with 100 mg seeds powder, the suspension ground together to a slurry, transferred to 2 mL Eppendorf tubes and centrifuged at 14000×g for 10 min at 4°C. The supernatant was collected and the protein concentration was measured according to Bradford18. Equal amount of proteins (35 mg) were used for electrophoresis, which was done in vertical native polyacrylamide gel, using 10% resolving gel and 3.5% stacking gel into Mini-Protean II System® BioRad (Hercules, CA, USA). All steps were carried out at 5°C at constant amperage (14 mA) using Tris-glycine buffer, pH 8.3. The isozyme activity among the 14 genotypes was assayed, according to Wendel and Weeden19. Each staining procedure was repeated three times. For comparing different isolates, the relative position of the stained band (Rf value) was determined as the ratio of migration distance from the origin of each band to migration distance from the origin of the dye marker (bromophenol blue).
SDS-PAGE for leaf and seed protein electrophoresis: Protein extraction was performed according to Saraswati et al.20. Seeds were ground to a powder with mortar and pestle in Liquid N2. Extraction buffer [0.5 M Tris-HCl (pH 6.8), 10% SDS, Urea and 5% 2-mercaptoethanol] was added to 40 mg of seed flour as extraction liquid and mixed thoroughly in Eppendorf tubes with the vortex. The tubes were centrifuged at 10,000×g for 5 min at 4°C. The supernatant was collected and boiled for 5 min before loading onto the gel. SDS-PAGE for the seeds was performed by a standard method on a vertical slab gel using 10% polyacrylamide gel according to the method of Laemmli21 and modified by Studier22.
| Table 1: | Origin, pedigree and main characteristics of 14 tested soybean genotypes |
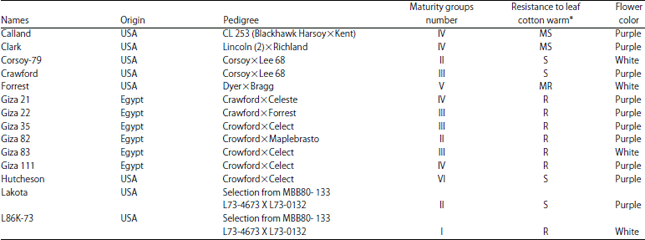 | |
*R: Resistant, S: Susceptible, MS: Moderately susceptible, MR: Moderate resistant | |
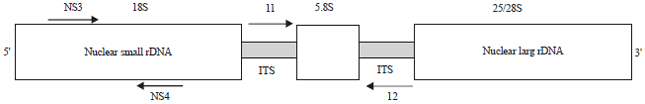 |
| Fig. 1: | Organization of the rDNA and primers proposed for soybean internal transcribed spacer (ITS). NS3 and NS4 primers produce 597 bp band while I1 and I2 primers produce 602 bp band |
Bromophenol blue was added to the supernatant as tracking dye to watch the movement of protein in the gel. After electrophoresis, the protein bands were visualized by staining with Coomassie Brilliant Blue G-250. Marker proteins (BioRad) were used as references. Molecular weights of protein bands were estimated by their relative mobility. The polymorphic bands were scored visually as present (1) or absent (0). Genetic similarity among genotypes was estimated based on Euclidian coefficients. Cluster analysis was performed using the UPGMA.
Restriction fragment length polymorphism (RFLP): Internal Transcribed Spacer (ITS) defined as each repeat unit consists of the genes coding the small subunit (16-18S), large subunit (23-28S) and the 5.8S (Fig. 1)23,24. Total DNA was isolated from young leaves using the CTAB methods as described by Doyle and Doyle25. The universal primers NS3, NS4, I1 and I2 were used for PCR amplification of rDNA26. Amplification reactions were performed in a 25 μL volume, containing: 20 mM Tris-HCl (pH = 8.4), 50 mM KCl, 2.0 mM MgCl2, 200 μM of each dNTPs, 1 μM of each primer, 30 ng of genomic DNA and 1.5 U of Taq DNA polymerase (Life Technologies, Switzerland). The temperature profile was as follows: initial denaturation at 94°C for 3 min; 35 cycles of denaturation at 94°C for 1 min, annealing at 55°C for 1 min and extension at 72°C for 2 min and a final extension step at 72°C for 7 min. Amplified PCR products were digested with the three restriction enzymes, that is, BamHI, MspI and TaqI as recommended by the manufacturer. To determine RFLP polymorphism among the fourteen soybean genotypes, the digested DNA fragments were separated on 0.6% agarose gels, visualized under UV illumination after staining with ethidium bromide and photographed. This was yielded specific restriction profiles that enabled direct visual identification of the 14 genotypes. To determine band size, a 100 base-pair ladder (Amersham Biosciences, CA, USA) was used in each electrophoretic run as a standard. All restriction patterns were coded in the binary form and analyzed using NTSYS-pc package27. A simple matching coefficient was calculated to construct a similarity matrix and the UPGMA algorithm was used to perform hierarchical cluster analysis and to construct a dendrogram.
Random amplified polymorphic DNA (RAPD): RAPD was used as described by Williams et al.15 for the identification of markers associated with the most sensitive and resistance among the 14 soybean genotypes for cotton leaf worm. Total DNA was isolated from young leaves using the CTAB extraction technique described by Doyle and Doyle25, with the exception that the extraction buffer contained 2% CTAB reagent with 100 mM Tris-HCl pH 8.0, 20 mM EDTA, 0.7 M NaCl, 0.3 M sorbitol, 0.5% polyvinylpyrrolidone, 2% polyethylene glycol and 0.3% 2-mercaptoethanol. For RAPD analysis, PCR amplifications were carried out in a total volume of 25 μL. The reaction contained 20 ng template DNA, 400 nM primer, 20 mM TRIS-HCl, 50 mM KCl, 1.5 mM MgCl2, 200 nM each dNTP and 1.0 unit Taq DNA polymerase (Life Technologies). Four different primers were used for this experiment as follow; A1A13 (5’-CAGGCCCTTCCA GCACCCAC-3’), A9B7 (5’-GGTGACGCAGGGGTAACGCC-3’), A1, (5’-CGAGCCCTTCCAGCACCCAC-3’) and A7 (5’-GAAACGGGTG GTGATCGCAG-3’). PCR was performed in MJ PTC-100 (MJ Research, Waltham, Mass., USA) thermocycler. The thermal cycle used was initial step at 94°C for 4 min and then 40 cycles of 94°C for 1min (denaturation), 36°C for 1min (annealing) and 72°C for 2 min (extension) followed by a final cycle of 94°C for 1 min, 35°C for 1 min and 72°C for 7 min for final extension. DNA fragments generated by amplification were separated according to size on 2% (w/v) agarose gels run in 1X TAE buffer (40 mM TRIS-Acetate, pH 9.0, 1.0 mM EDTA), stained with ethidium bromide, visualized by UV transilluminator and photographed by a GelCam camera, Polaroid. To determine band size, a 100 base-pair ladder (Amersham Biosciences) was used in each electrophoretic run as a standard. To generate a binary matrix for morphological data, the presence or absence of a band for the 14 individual’s genotypes with the four primers used in the RAPD analyses was marked as 1 for present or 0 for absent. Morphological variables were standardized and Euclidean distances were calculated following the procedure described by Sokal and Michener28 and Roldan-Ruiz et al.29.
RESULTS AND DISCUSSION
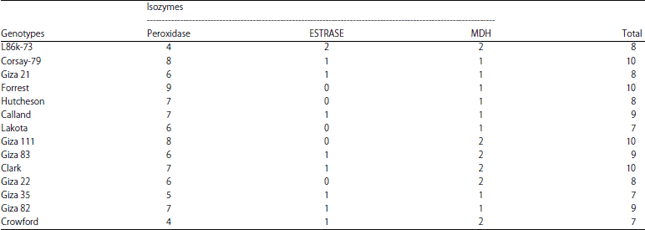
Leaf isozymes: The leaf isozymes polymorphism among soybean genotypes are presented in Fig. 2 and Table 2.
| Table 2: | Total number of bands from three isozymes of different soybean genotypes |
 | |
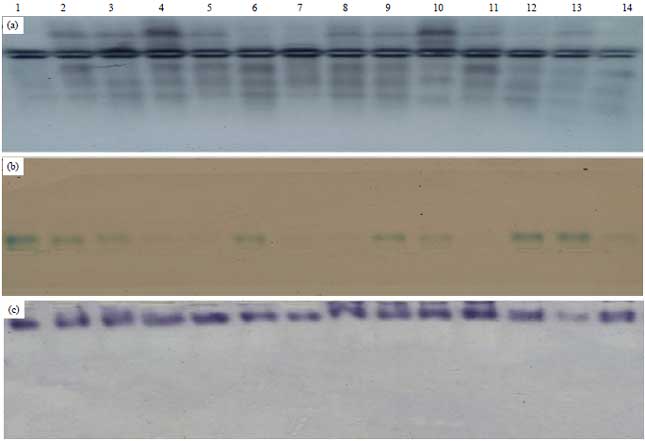 |
| Fig. 2(a-c): | Activity analysis of isozymes in soybean genotypes based on Natve-PAGE electrophoresis, (a) Peroxidase (PRX), (b) Esterase (EST) and (c) Malate dehydrogenase (MDH). Soybean genotypes: 1: L86K-73, 2: Corsay-79, 3: Giza 21, 4: Forrest, 5: Hutcheson, 6: Calland, 7: Lakota, 8: Giza 111, 9: Giza 83, 10: Clark, 11: Giza 22, 12: Giza 35, 13: Giza 82 and 14: Crowford |
| Table 3: | Similarity indexes among the 14 soybean genotypes based on leaf and seed protein SDS-PAGE |
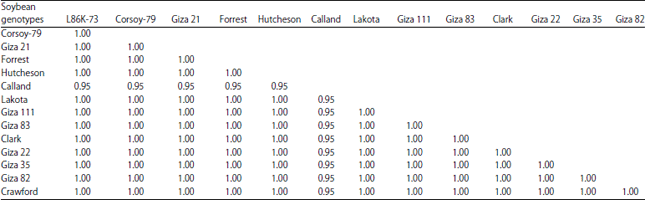 | |
The Data showed a qualitative analysis depends on the number of obtained bands from each soybean genotypes. In Peroxidase (Fig. 2a), the data indicated that the moderately resistances genotypes have a few bands between 7 and 9 (lane 4, 6 and 10) while the tolerance and susceptible genotypes had a number of bands between 4 and 8 (lanes, 1, 3, 8, 9, 11, 12, 13 for tolerant and lanes 2, 5, 7, 14 for susceptible). Isozymes Peroxidase presented bands as a positive marker for Forrest genotypes (Table 2). Moreover, the genotypes L86K-73 and Crowford did not give any bands in a negative form which is considered as negative markers. On the other hand, the density of bands was highest in Forrest and Clark, which were moderately resistance and moderately susceptible to cotton leaf worm, respectively. In Esterase (Fig. 2b) only L86K-73 genotype presented 2 bands which were considered as positive markers. High dense bands were observed in Giza 35, Giza 82 and L86K-73 which considered tolerant genotypes. Regards to malate dehydrogenase (MDH)(Fig. 2c) genotypes L86K-73, Giza 111, Giza 83, Clark, Giza 22 and Crowford had 2 bands while the other genotypes had one band. The above results indicated that no clear trend has been observed in isozymes analysis regarding differentiation among 14 soybean genotypes based on their resistance to cotton leaf worm. Isozymes as a molecular marker were not able to detect significant different between the tested soybean genotypes according to their tolerant/susceptible to cotton leaf worm.
SDS-PAGE of seed proteins: The results of SDS-PAGE for the water-soluble leaf proteins in the soybean genotypes (Fig. 3a) revealed a total number of 8 bands with molecular weights (MW) ranging from about 14.4-126.3 KDa. Data showed that all bands were common for the 14 soybean genotypes (monomorphic), however, they differed in their intensity. The densitometric analysis (Table 3) of SDS-protein banding patterns of the leaf among the studied genotypes was found to be not informative regarding resistance/susceptible of soybean genotypes to cotton leaf worm. On the other side, the results of SDS-PAGE for the seed water-soluble proteins in the soybean genotypes (Fig. 3b) exposed a total number of 13 bands with molecular weights (MW) ranging from about 9.1-220.36 KDa. Data showed that all bands are common for the fourteen genotypes except two bands with molecular weight 220.36 and 194 KDa for Calland genotype (moderately susceptible) can be considered as specific bands, which were differed in their intensity. The overall results of total protein pattern obtained by SDS-PAGE did not show clear-cut regards to monitoring molecular markers resistance/ susceptible to cotton leaf worm of the fourteen soybean genotypes based on dendrogram for the genetic distances relationships among the genotypes by similarity indices data of protein SDS-PAGE analysis (Fig. 3c). These results are in agreement with similar results also in soybean presented by Helms et al.8 and Zietkiewicz et al.30.
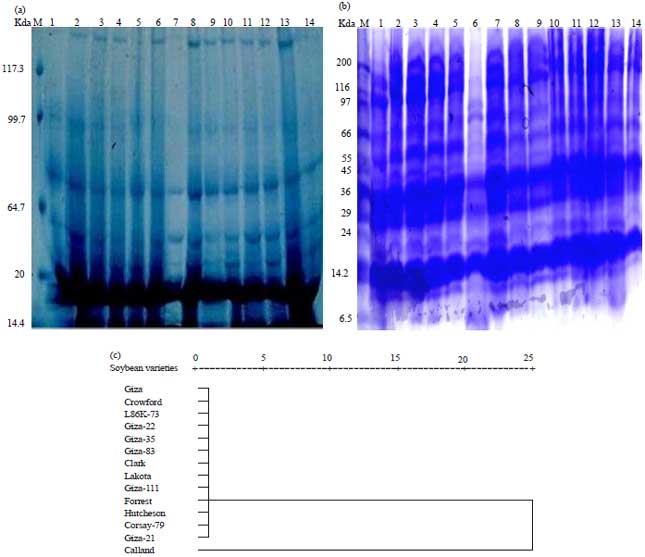
Restriction fragment length polymorphism (RFLP) for ITS: RFLP was also used to analyze the genetic diversity of the fourteen soybean genotypes and to assess their genetic relationships using similarity indices and dendrogram tree. The PCR amplification using primers based on nucleotide sequences of soybean ITS regions (Fig. 1) was performed. Three restrictions enzymes BamHI, MspI and Taql were used to cut the PCR products to determine RFLP polymorphism among soybean genotypes. The digested products were separated on agarose gel electrophoresis yielded specific restriction profiles that enabled direct visual identification of the genotypes (Fig. 4). Restriction site analysis of the ITS region amplified by PCR using universal primers pair was able to successfully amplify the ITS region of all genotypes tested, providing a 602 bp PCR product. The patterns of the RFLP analysis were analyzed for each genotype with BamHI (Fig.4a) produced a similar band 285 bp for each soybean genotype.
 |
| Fig. 3(a-c): | Protein analysis using SDS-PAGE, (a) SDS-PAGE banding patterns for leaves water-soluble protein of soybean genotypes, (b) SDS-PAGE banding patterns for seeds of water-soluble protein of soybean genotypes and (c) Dendrogram for the genetic distances relationships among the fourteen genotypes based on similarity indices data of protein SDS-PAGE analysis. Soybean genotypes: 1: L86K-73, 2: Corsay-79, 3: Giza 21, 4: Forrest, 5: Hutcheson, 6: Call, 7: Lakota, 8: Giza 111, 9: Giza 83, 10: Clark, 11: Giza 22, 12: Giza 35, 13: Giza 82 and 14: Crowford. M: Protein marker |
Digestion of PCR products by MspI (Fig. 4b) resulted one band 870 bp with L86K-73 and four bands (65, 275, 417 and 730 bp) with the rest genotypes. While, Taql (Fig. 4c) enzyme give six bands (30, 50, 70, 90, 430 and 580 bp) in all genotypes except L86K-73 and Corsay-79 which gave only one band at 1400 and 30 bp, respectively.
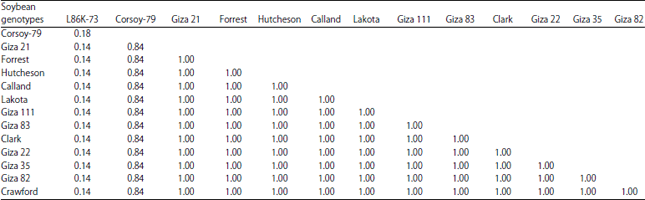
Data in Fig. 4d and Table 4 showed similarity indexes was 1.00 in all genotypes except L86k-73 and Corsay give 0.182 and 0.842, respectively. The dendrogram that shows the genetic distance among 14 soybean genotypes based RFLP data presented in Fig. 4d. The similarity indices of tested genotypes based on PFLP for ITS- PCR using 3 restriction enzymes is presented in Table 4. The data indicated that there are 12 genotypes genetically related. Olsen and Woese24 explained ribosomal DNA (rDNA) sequences have been aligned and compared in several living organisms and this approach has provided a wealth of information about phylogenetic relationships. Studies of rDNA sequences have been used to infer phylogenetic history across a very broad spectrum, from studies among the basal lineages of life to relationships among closely related species and populations.
 |
| Fig. 4(a-d): | Restriction fragment length polymorphism (RFLP) for ITS using PCR and restriction enzymes, (a) BamHI, (b) MspI, (c) Taql and (d) Dendrogram for the genetic distances relationships among the fourteen genotypes based on similarity indices data of RFLP analysis. Soybean genotypes: 1: L86K-73, 2: Corsay-79, 3: Giza 21, 4: Forrest, 5: Hutcheson, 6: Calland, 7: Lakota, 8: Giza 111, 9: Giza 83, 10: Clark, 11: Giza 22, 12: Giza 35, 13: Giza 82 and 14: Crowford. MM: DNA molecular marker |
| Table 4: | Similarity indices among the fourteen soybean genotypes based on RFLP-PCR using 3 restriction enzymes |
 | |
The reasons for the systematic versatility of rDNA include the numerous rates of evolution among different regions of rDNA (both among and within genes), the presence of many copies of most rDNA sequences per genome and the pattern of concerted evolution that occurs among repeated copies. These features facilitate the analysis of rDNA by direct RNA sequencing, DNA sequencing (either by cloning or amplification) and restriction enzyme methodologies. The RFLP markers that used in this experiment may not have been linked to the resistant/susceptible trait to cotton leaf worm in the tested soybean genotypes. In the future, it may be necessary to identify a large number of markers closely linked to this trait.
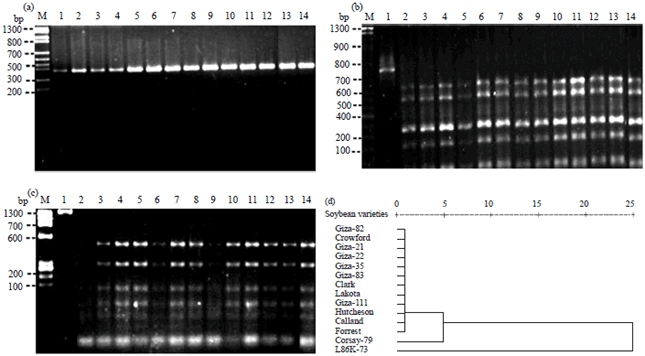
Random amplified polymorphic DNA (RAPD): The resulted amplified fragments are shown in Fig. 5. Banding patterns were scored as present (1) or absent (0). All the 4 primers successfully amplified DNA fragments for all tested soybean genotypes with a total number of 62 fragments were visualized across the different genotypes.
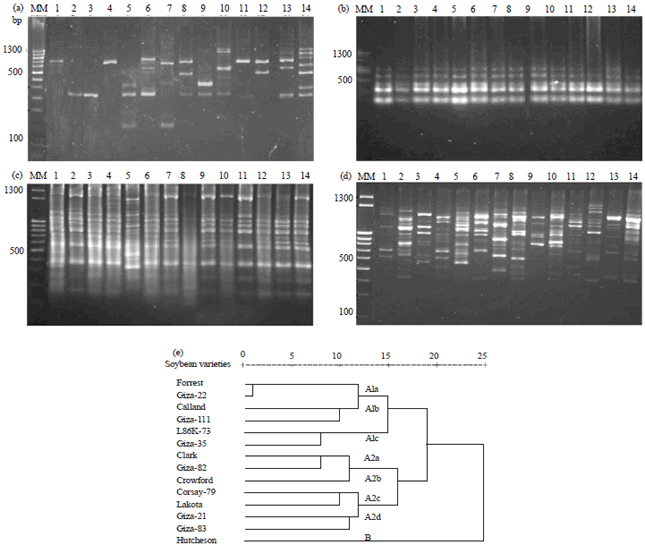 |
| Fig. 5(a-e): | DNA polymorphism based on RAPD-PCR analysis of the soybean genotypes (a) Primer A9B7, (b) Primer A7, (c) Primer A1, (d) Primer A1A13 and (e) Dendrogram for the genetic distances relationships among the 14 genotypes based on similarity indices data of RAPD analysis. Soybean genotypes: 1: L86K-73, 2: Corsay-79, 3: Giza 21, 4: Forrest, 5: Hutcheson, 6: Calland, 7: Lakota, 8: Giza 111, 9: Giza 83, 10: Clark, 11: Giza 22, 12: Giza 35, 13: Giza 82 and 14: Crowford. MM: DNA molecular marker |
The pattern produced by primer A9B7 showed a maximum number of 12 DNA fragments ranging in molecular sizes between 130-1670 bp (Fig. 5a). Twelve polymorphic fragments (100%) were observed among the genotypes, whereas, no monomorphic fragments were observed. Crowford genotype showed the maximum number of fragments, while the lowest number appeared in Forrest genotype. Primer A7 showed a maximum of five DNA fragments with molecular sizes ranged from 150-670 bp (Fig. 5b). Zero polymorphic fragments (0%) and monomorphic (100%) were recorded in all genotypes. Primer A1 exhibited nineteen DNA fragments ranging in molecular sizes from 50-1080 bp (Fig. 5c). Eleven polymorphic fragments (58%) were observed, while the other eight fragments (42%) were monomorphic. Figure 5d showed that primer A1A13 gave 26 polymorphic fragments (100%) with molecular sizes ranging from 270-1007 bp. Moreover, no monomorphic was detected among these genotypes using A1A13 primer. Crowford genotype showed the maximum number of fragments, while the lowest number appeared in L86K-73 genotype. The genotypes had specific molecular markers in bands 960 bp for Hutcheson, 740 bp for Forrest and 360 bp for Giza, as positive markers.
| Table 5: | Total number of amplified fragments and specific markers of the 14 soybean genotypes based on RAPD-PCR analysis using 4 primers |
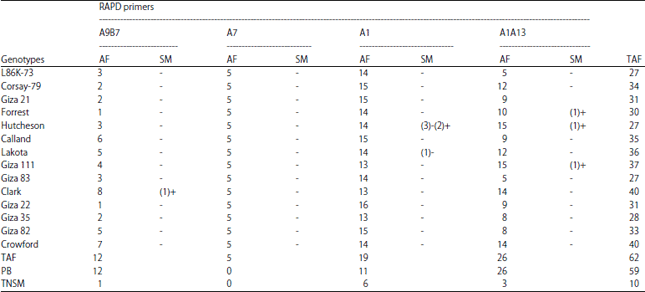 | |
TAF: Total amplified fragments, AF: Amplified fragments, SM: Specific markers including either the presence (+) or absence (-) of a fragment, PB: Polymorphic bands, and TNSM: Total number of specific markers | |
| Table 6: | Similarity indexes among the 14 soybean genotypes based on RAPD-PCR using 4 primers |
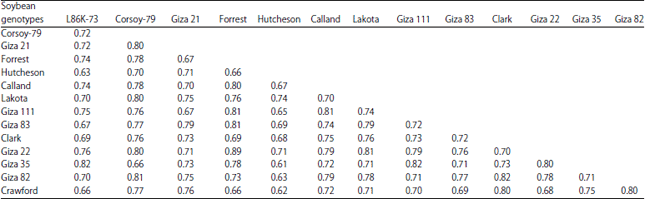 | |
These results exposed success in amplifying DNA fragments from soybean genotypes. Polymorphism levels differed from one primer to the other, the number of total amplified fragments (TAF) and polymorphic bands (PB) for each primer, amplified fragments (AF) and specific markers (SM) for soybean genotypes using the 4 primers are shown in Table 5. These high levels of polymorphism served in soybean molecular markers, which can be used to discriminate each soybean genotypes from the others. There were several specific fragments either present in only one genotype and absent in all others (positive marker) or absent in only one genotype and present in all others (negative marker) (Table 5).
By looking at these results, it could be concluded that: (1) Primer A9B7 showed specific fragment (520 bp) as positive marker for Clark genotype, (2) Primer A1 showed six specific fragments four of them as negative markers, three for Hutcheson genotype (430, 280 and 50 bp) and one for Lakota genotype (810 bp) and also two positive markers (360 and 320 bp) for Hutcheson, (3) Primer A1A13 showed three specific fragments as positive markers 960 bp for Hutcheson, 740 bp for Forrest and 360 bp for Giza 111 and (4) Primer A7 did not show specific fragment for any genotype. Results of cluster analysis (similarity index) and dendrogram for the genetic relationships based on RAPD-PCR data among the 14 soybean genotypes across the four primers using UPGMA computer analysis presented in Fig. 5e and Table 6. The highest similarity value recorded was 0.89 between Forrest which considers moderately resistance and Giza 22 which is tolerance genotypes, while the lowest similarity value recorded was 0.61 between Hutcheson (susceptible) and Giza 35 (tolerance) genotypes.
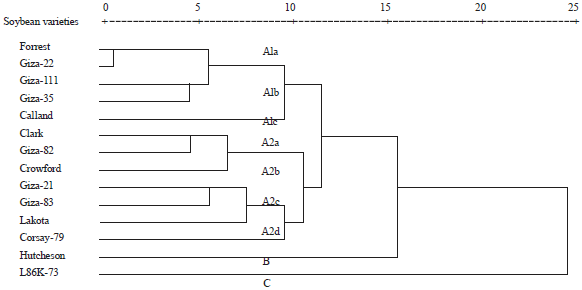 |
| Fig. 6: | Dendrogram for the genetic distances relationships among the 14 genotypes based on similarity indices data of protein SDS-PAGE, RFLP and RAPD analysis |
The fourteen soybean genotypes were separated into two clusters (Fig. 5e). Cluster A included two subcluster, the subcluster A1 consisted of three groups, group A1a [Forrest (moderately resistance) and Giza 22 (tolerance)], group A1b [Calland (moderately resistance) and Giza 111 (tolerance)] and group A1c [Giza 35 and L86K-73 (tolerance)]. While subcluster A2 comprised four groups, group [A2a (Clark (moderately resistance) and Giza 82 (tolerance)], group A2b (Crowford only susceptible genotype), group A2c [Lakota and Corsay-79 (susceptible)] and group A2d [Giza 21 and Giza 83 (tolerance)]. Single, dominant R-genes to plant pathogens often occur in multigene clusters in many of the plant genomes that have been studied31 including soybean32. The second cluster B was contained Hutcheson genotype (Fig. 5e). The dendrogram reflects the performance of genotypes based on their susceptible to cotton leaf worm. For example, the genotypes in subcluster A1 (Forrest, Giza 22, Calland, Giza 111, Giza 35 and L86K-73) all are resistance except Calland which considered moderately resistance. On the other hand, the most susceptible genotypes Hutcheson is located alone in cluster B. Therefore, RAPD-PCR technique was used as a tool for classification the different soybean genotypes according to their susceptibility to cotton leave worm. PAPD-PCR technique has been used previously to identify soybean genotypes based on their performance to insect resistance. For example, Carvalho et al.33, who used micro satellites (Satt187 and Satt309) and three RAPD markers (OPAG-05946, OPF-041038 and OPAQ-011987) to identify molecular markers associated with the resistance to race 3 of the soybean cyst nematode (SCN). Narvel et al.34 noted that there has been a limited success over the past three decades in the development of superior soybean genotypes with insect resistance. Success may be hampered by the quantitative nature of resistance and by linkage drag from tolerant plant introduction (PI) donor parents. Soybean insect resistance quantitative trait loci (SIR QTLs) have been identified from PI 229358 and PI 171451 by restriction fragment length polymorphism (RFLP) analysis8,34. RAPD data defined as a measure of alike in state for predominantly non-coding regions of the genome8. Because the soybean genome comprises of approximately 40% single-copy and 60% repetitive DNA35, the DNA markers should be scattered in the same ratio among these two DNA classes. Though the single-copy DNA and repetitive DNA classes encompass expressed sequences, most of the genome complexity is not associated with expressed genes36. Consequently, most RAPD markers would be associated with non-expressed and neutral sequences that should not be affected by selection. Thus, the marker data should be reflective of the genetic distance among the genotypes.
Combined data based on SDS-PAGE protein profile, RFLP and RAPD: A combined analysis based on protein electrophoresis, RFLP and RAPD-PCR analyses was carried out using UPGMA computer program and shown in Fig. 6 and Table 7. The highest similarity index recorded was 0.944 between L86K-73 (tolerance) and Hutcheson (susceptible) genotypes, while the lowest similarity index 0.700 was observed between Forrest (moderately resistance) and Giza 22 (tolerance) genotypes. Dendrogram for the genetic relationships among the fourteen soybean genotypes across the three techniques results was carried out as in Fig. 6.
| Table 7: | Similarity indices among the 14 soybean genotypes based on protein SDS-PAGE, RFLP and RAPD-PCR using 3 restriction enzymes |
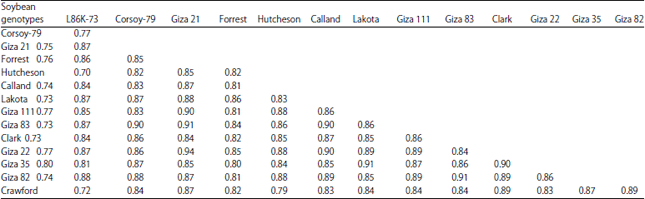 | |
The fourteen soybean genotypes were separated into three clusters; the cluster A was the largest and divided into two subcluster. Within the subcluster A1, three groups appeared; group A1a contained Forrest and Giza 22 genotypes, group A1b contained Giza 111 and Giza 35 genotypes and group A1c contained Calland only. The Second subcluster A2 consists of four groups separated into two divisions (Fig. 6). The first division appeared in two groups A2a and A2b. The group A2a has Clark and Giza 82, while the second group A2b contained Crowford only. The second division has groups from three to four. The group number A2c contained Giza 21, Giza 83 and Lakota, group A2d contained Corsay-79 only. On the other side, cluster B included Hutcheson while cluster C included L86K-73. However, the combined data of the three techniques used (protein, RAPD and PFLP) produced the same cluster results as the RAPD technique. That is to confirm that RAPD can be considered as a reliable technique for studying relationships among soybean genotypes. In the study of genetic diversity the use of protein electrophoresis, RFLP and RAPD analyses seemed to be powerful tools and could discriminate among the fourteen soybean genotypes9. Casas et al.37 confirmed that both protein and RAPD results appear to play an important role in the differentiation among different cultivars. In the current study, protein analysis did not provide enough information to segregate the fourteen tested genotypes but the RAPD data separated the genotypes into different groups based on their tolerant/susceptible to cotton leaf worm. Moreover, Li et al.38 studied the feasibility of developing RAPD-based diagnostic dot blot tests to separate field isolates of soybean cyst nematode (Heteroder aglycines) with different virulence to tolerant cultivars of soybean. Yencho et al.39 mentioned that molecular markers can be used to increase our understanding of the mechanisms of plant resistance to insects and develop insect tolerant crops.
CONCLUSION
No clear trend had been observed in isozymes analysis regarding differentiation among 14 soybean genotypes based on their resistance to cotton leaf worm. SDS-PAGE for the water-soluble seed proteins in the soybean genotypes exposed a total number of 13 bands most of them are common for the fourteen genotypes except two bands with molecular weight 220.36 and 194 KDa for that are specific for Calland genotype (moderately susceptible). Restriction site analysis of the ITS region amplified by PCR using universal primers pair was able to successfully amplify the Internal Transcribed Spacer (ITS) region of all genotypes tested. RAPD analysis using four different primers showed that primer A9B7 and A1A13 have 100% polymorphic, however primer A7 and A1 have 0 and 58% polymorphic, respectively. This high level of polymorphism could serve as molecular markers in soybean that could be used to discriminate each soybean genotype according to the susceptibility/resistance to cotton leave worm. A combined analysis based on protein electrophoresis, RFLP and RAPD-PCR analyses showed that the highest similarity index recorded was 0.944 between L86K-73 (tolerance) and Hutcheson (susceptible) genotypes, while the lowest similarity index 0.700 was observed between Forrest (moderately resistance) and Giza 22 (tolerance) genotypes.
SIGNIFICANCE STATEMENT
The combined data of three different techniques used (protein, RFLP and RAPD) produced the same cluster results as the RAPD technique. Therefore, RAPD can be considered as a reliable technique for studying relationships among soybean genotypes. In the study of genetic diversity, the use of protein electrophoresis, RFLP and RAPD analyses seemed to be powerful tools and could discriminate among the fourteen soybean genotypes.
REFERENCES
- Sakthivelu, G., M.K. Akitha Devi, P. Giridhar, T. Rajasekaran, G.A. Ravishankar, T. Nedev and G. Kosturkova, 2008. Drought-induced alterations in growth, osmotic potential and in vitro regeneration of soybean cultivars. Gen. Applied Plant Physiol., 34: 103-112.
Direct Link - Mohamed, H.I. and A.G. Abd-El Hameed, 2014. Molecular and biochemical markers of some Vicia faba L. genotypes in response to storage insect pests infestation. J. Plant Interact., 1: 618-626.
CrossRefDirect Link - Aydin, H. and M.O. Gurkan, 2006. The efficacy of spinosad on different strains of Spodoptera littoralis (Boisduval) (Lepidoptera: Noctuidae). Turk. J. Biol., 30: 5-9.
Direct Link - Mohamed, H.I., A. Haleem, M.A. Mohammed and A.M. Mogazy, 2016. Effects of plant defense elicitors on soybean (Glycine max L.) growth, photosynthetic pigments, osmolyts and lipid components in response to cotton worm (Spodoptera littoralis) infestation. Bangladesh J. Bot., 45: 597-604.
Direct Link - Storfar, A., 1996. Quantitative genetics: A promising approach for the assessment of genetic variation in endangered species. Trends Ecol. Evol., 11: 343-348.
CrossRefDirect Link - Choudhary, K., O.P. Choudhary and N.S. Shekhawat, 2008. Marker assisted selection: A novel approach for crop improvement. Am.-Eur. J. Agron., 1: 26-30.
Direct Link - Semagn, K., A. Bjornstad and M.N. Ndjiondjop, 2006. An overview of molecular marker methods for plants. Afr. J. Biotechnol., 5: 2540-2568.
Direct Link - Helms, T., G. Vallad, P. McClean and J. Orf, 1997. Genetic variance, coefficient of parentage and genetic distance of six soybean populations. Theor. Applied Genet., 94: 20-26.
CrossRefDirect Link - He, X.D., G. Zhong, G.J. Yi, Z.Z. Yong, C.S. Yi, H. Fukushi and Y. Shimamoto, 1999. Genetic diversity and evolutionary tendency detected by isoenzyme, RFLP and RAPD markers in wild and cultivated soyabeans in China. Scient. Agric. Sin., 32: 16-22.
Direct Link - Woodfield, D.R. and J.R. Caradus, 1994. Genetic improvement in white clover representing six decades of plant breeding. Crop Sci., 34: 1205-1213.
CrossRefDirect Link - Zarkadas, C.G., C. Gagnon, S. Gleddie, S. Khanizadeh, E.R. Cober and R.J.D. Guillemette, 2007. Assessment of the protein quality of fourteen soybean [Glycine max (L.) Merr.] cultivars using amino acid analysis and two-dimensional electrophoresis. Food Res. Int., 40: 129-146.
CrossRefDirect Link - Junyi, G., 2006. Segregation analysis on genetic system of quantitative traits in plants. Front. Biol. China, 1: 85-92.
CrossRefDirect Link - Malaviya, D.R., B. Kumar, A.K. Roy, P. Kaushal and A. Tiwari, 2005. Estimation of variability of five enzyme systems among wild and cultivated species of trifolium. Genet. Resour. Crop Evol., 52: 967-976.
CrossRefDirect Link - Williams, J.G.K., A.R. Kubelik, K.J. Livak, J.A. Rafalski and S.V. Tingey, 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res., 18: 6531-6535.
CrossRefPubMedDirect Link - Kaushal, P., D.R. Malaviya, A.K. Roy, B. Kumar and A. Tiwari, 2005. Trifolium alexandrinum × T. resupinatum-Interspecific hybrids developed through embryo rescue. Plant Cell. Tissue Organ Cult., 83: 137-144.
CrossRefDirect Link - Kumar, B., D.R. Malaviya, A.K. Roy and P. Kaushal, 2008. Isozyme variability in Trifolium alexandrinum accessions. Indian J. Genet. Plant Breed., 68: 195-200.
Direct Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Saraswati, R., T. Matoh, T. Sasai, P. Phupaibul, T.A. Lumpkin, M. Kobayashi and J. Sekiya, 1993. Identification of Sesbania species from electrophoretic patterns of seed proteins. Trop. Agric., 70: 282-285.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Studier, F.W., 1973. Genetic analysis of non-essential bacteriophage T7 genes. J. Mol. Biol., 79: 227-236.
CrossRefDirect Link - Hillis, D.M. and M.T. Dixon, 1991. Ribosomal DNA: Molecular evolution and phylogenetic inference. Quart. Rev. Biol., 66: 411-453.
CrossRefPubMedDirect Link - Olsen, G.J. and C.R. Woese, 1993. Ribosomal RNA: A key to phylogeny. FASEB J., 7: 113-123.
CrossRefPubMedDirect Link - Weisburg, W.G., S.M. Barns, D.A. Pelletier and D.J. Lane, 1991. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol., 173: 697-703.
CrossRefPubMedDirect Link - Sokal, R.R. and C.D. Michener, 1958. A statistical method for evaluating systematic relationships. Univ. Kansas Sci. Bull., 38: 1409-1438.
Direct Link - Roldan-Ruiz, I., F.A. van Eeuwijk, T.J. Gilliland, P. Dubreuil and C. Dillmann et al., 2001. A comparative study of molecular and morphological methods of describing relationships between perennial ryegrass (Lolium perenne L.) varieties. Theor. Applied Genet., 103: 1138-1150.
CrossRefDirect Link - Zietkiewicz, E., A. Rafalski and D. Labuda, 1994. Genome fingerprinting by Simple Sequence Repeat (SSR)-anchored polymerase chain reaction amplification. Genomics, 20: 176-183.
CrossRefPubMedDirect Link - Meyers, B.C., S. Kaushik and R.S. Nandety, 2005. Evolving disease resistance genes. Curr. Opin. Plant Biol., 8: 129-134.
CrossRefDirect Link - Kanazin, V., L.F. Marek and R.C. Shoemaker, 1996. Resistance gene analogs are conserved and clustered in soybean. Proc. Natl. Acad. Sci. USA., 93: 11746-11750.
CrossRefDirect Link - Carvalho, G.A., T. Sediyama, A.L.A. Marin, E.G. Barros and M.A. Moreira, 2002. Identification of RAPD markers linked to a soybean stem canker resistance gene. Fitopatol. Bras., 27: 474-478.
CrossRefDirect Link - Narvel, J.M., D.R. Walker, B.G. Rector, J.N. All, W.A. Parrott and H.R. Boerma, 2001. A retrospective DNA marker assessment of the development of insect resistant soybean. Crop Sci., 41: 1931-1939.
CrossRefDirect Link - Goldberg, R.B., 1978. DNA sequence organization in the soybean plant. Biochem. Genet., 16: 45-58.
CrossRefDirect Link - Goldberg, R.B., G. Hoschek, J.C. Kamaley and W.E. Timberlake, 1978. Sequence complexity of nuclear and polysomal RNA in leaves of the tobacco plant. Cell, 14: 123-131.
CrossRefDirect Link - Casas, A.M., E. Igartua, G. Balaguer and M.A. Moreno, 1999. Genetic diversity of Prunus rootstocks analyzed by RAPD markers. Euphytica, 110: 139-149.
CrossRefDirect Link - Li, J., J. Faghihi, J.M. Ferris and V.R. Ferris, 1996. The use of RAPD amplified DNA as markers for virulence characteristics in soybean cyst nematode. Fundam. Applied Nematol., 19: 143-150.
Direct Link - Yencho, G.C., M.B. Cohen and P.F. Byrne, 2000. Applications of tagging and mapping insect resistance loci in plants. Ann. Rev. Entomol., 45: 393-422.
CrossRefPubMedDirect Link