
Kazi Muhammad Rezaul Karim
Institute of Nutrition and Food Science, University of Dhaka, Dhaka 1000, Bangladesh
LiveDNA: 880.17997
Ahmad Husaini
Department of Molecular Biology, Faculty of Resource Science and Technology, Universiti Malaysia Sarawak, 94300 Kota Samarahan, Sarawak, Malaysia
Tasmia Tasnim
Department of Nutrition and Food Engineering, Faculty of Allied Health Science, Daffodil International University, Dhaka 1207, Bangladesh
American Journal of Biochemistry and Molecular Biology
Year: 2017 | Volume: 7 | Issue: 3 | Page No.: 118-126







ABSTRACT
Background and Objective: Glucoamylase are inverting exo-acting starch hydrolases releasing β-glucose from the non-reducing ends and are used in the manufacture of glucose, fructose syrups and other industrial purposes. The purpose of the study was to optimize the preferable environmental condition for the production of glucoamylase and its characterization from Aspergillus flavus (A. flavus) NSH9. Methodology: The effects of various parameters like carbon and nitrogen sources, temperature, pH, incubation period and other supplements were investigated for the production of glucoamylase. One-way ANOVA and independent samples t-test were used to analyze the data. Results: The maximum production of glucoamylase was observed at 25°C with initial pH 5.0 after 5 days of incubation. Depending on the type and amount of carbon sources, 6% soluble starch was considered as best for the glucoamylase production followed 2% raw sago starch. Yeast extract was considered the best as organic and urea was as inorganic sources of nitrogen. The glucoamylase from Aspergillus flavus NSH9 exhibited optimum activity in a range of pH 4.0-7.0 with a maximum activity at pH 5.0 and the pH stability (more than 95%) ranging from 4-9 at 25°C after 24 h. Conclusion: Glucoamylase from Aspergillus flavus NSH9 have high optimum temperature (70°C) for activity and good thermostablity in the absence of substrate. The crude glucoamylase was also found capable of hydrolyzing the raw starch.
PDF Abstract XML References Citation
Received: August 29, 2017;
Accepted: October 09, 2017;
Published: October 18, 2017
How to cite this article
Kazi Muhammad Rezaul Karim, Ahmad Husaini and Tasmia Tasnim, 2017. Production and Characterization of Crude Glucoamylase from Newly Isolated Aspergillus flavus NSH9 in Liquid Culture. American Journal of Biochemistry and Molecular Biology, 7: 118-126.
DOI: 10.3923/ajbmb.2017.118.126
URL: https://scialert.net/abstract/?doi=ajbmb.2017.118.126
DOI: 10.3923/ajbmb.2017.118.126
URL: https://scialert.net/abstract/?doi=ajbmb.2017.118.126
INTRODUCTION
Glucoamylase (Glucan 1,4-α-glucohydrolase, EC 3.2.1.3) is an exo-acting enzyme that yields β-D-glucose from the nonreducing ends of amylose, amylopectin and glycogen by hydrolyzing α-1,4 linkages in a consecutive manner. To a lesser extent, it also has the ability to hydrolyze α-1, 6 linkages and there are α-1, 3 linkages albeit at a much slower rate, resulting in glucose as the end-product and hence called the saccharifying enzyme1. Glucoamylase (GA) is produced by a large number of microbes, including fungi, bacteria and yeast2. Nevertheless, most of the glucoamylase used in the industry is derived from Aspergillus oryzae, Aspergillus niger and Rhizopus oryzae3,4. The advantage of using micro-organisms for the production of enzymes is that bulk production is economical and microbes are easy to manipulate to obtain enzymes with desired characteristics. Fungal enzymes are preferred over other microbial sources owing to their widely accepted generally regarded as safe (GRAS) status5. Amylase or glucoamylase has a significant role in starch processing in the food industries, such as for the fructose and glucose syrup that are produced from liquefied starch with an action of amylase and glucoamylase6,7. Amylase is also used in bioethanol, confectionary, pharmaceuticals, beverage and different fermented food in industries7-9. Enzyme production and characterization of various sources is the new field of research in biotechnology. Thermostability is a desired characteristic of most of the industrial enzymes and each application of industrial enzymes requires unique properties with respect to specificity, stability, temperature and pH dependence10. At the same time, raw starch degrading glucoamylase that is capable of hydrolyzing starch at moderate temperature could save the energy and reduce the cost of production in starch processing industry11. Therefore, a study on the enzymes that is either thermostable or capable of degradingraw starch granules without gelatinization would be crucial to conserve the energy consumed. The present study was undertaken to optimize the growth conditions for maximal production of the glucoamylase by Aspergillus flavus NSH9 in liquid culture and its characterization.
MATERIALS AND METHODS
Production of glucoamylase and selection of optimum temperature and incubation period: The study was carried out from December, 2013-December, 2015, at the department of Molecular biology, Universiti Malaysia Sarawak, Kuching, Malaysia. As for the enzyme induction, the actively growing fungal mycelium was transferred from Potato Dextrose Agar (PDA) plate to a culture medium containing (in g L –1) of: 20 g raw sago starch, 3 g KH2PO4, 1 g (NH4)2SO4, 0.5 g MgSO4.7H2O and 4 g of yeast extract (raw sago starch were sterilized separately in a dry oven). One piece (5 mm2 in diameter) of 7 days old fungal culture was used for the fermentation of 50 mL MSM medium containing 2% (w/v) raw sago starch. The incubation was carried out at different temperature ranging from 20-40°C for 7 days on a rotary shaker at150 rpm. Samples were withdrawn from cultures for each batch (temperature) at different time intervals of 1, 2, 3, 4, 5, 6 and 7 days. The samples were then filtrated through muslin cloth before centrifuged at 6000 rpm at 4°C for 20 min to remove the mycelia. The supernatant was harvested and then filtered through a Whatman filter paper No. 1and used for the assay of glucoamylas activity and the remaining sample was stored at-20°C for further analysis.
Optimum pH for glucoamylase production: For determining the effect of pH on enzyme production, various pH values of initial cultures were adjusted to 3, 4, 5, 6, 7 and 8 to find its optimum value at optimum temperature and incubation period.
Effect of carbon and nitrogen sources: Four carbon sources such as raw sago starch, soluble starch, raw cassava starch and raw sweet potato starch in GA production were studied by using different concentration (1, 2, 4 and 6%) at optimum temperature, pH and incubation period. To observed the nitrogen effect on glucoamylase production, five nitrogen sources: Organic (3 g L –1) yeast extract, beef extract and inorganic: NaNO3, (NH4)2SO4 and urea were added at the same concentration ofnitrogen {0.318 g L –1 N equivalent of 1 g of (NH4)2SO4} with 2% raw sago starch as carbon sources at optimum temperature, pH and incubation period.
Effect of inoculum size on biomass and glucoamylase production: To measure the effect of inoculum size on glucoamylase production, 1 piece (5 mm) to 5 pieces of 7 days old fungal cultures were added in the fermentation media. The incubation was carried out at room temperature (25-27°C) for 5 days in a rotary shakerat 150 rpm. The amount of biomass produced after fermentation in different plugs were collected by filtered the culture through a Whatman filter paper No 1. After that the biomass were kept in oven at 65°C for 24 h for drying to measure the weight.
Glucoamylase and protein assay: Glucoamylase activity was determined by transferring a volume of 0.5 mL of crude enzyme followed by the addition of 0.5 mL of 1% soluble starch solution in 0.1 M sodium acetate buffer (pH5.0) and incubated at 55°C for 30 min. The released glucose was measured using 3, 5-Dinitrosalicyclic acid (DNS) reagent according to Miller12 and measured at 540 nm. The enzyme activities were calculated using a calibration curve prepared with D-glucose as standard. One unit of glucoamylase activity was defined as the amount of enzyme that released 1 μmol of glucose equivalent per min from soluble starch under the assay condition (at 55°C and pH 5.0).
Raw starch degrading enzymes (RSDE) refer to enzymes that can directly degrade raw starch granules below the gelatinization temperature of starch. The degrading ability of crude glucoamylase preparation towards raw starch were determined by mixing 0.5 mL of enzyme preparation with 0.5 mL of 1% raw sago starch in 0.1M sodium acetate buffer, pH 5. After 24 h of incubation at 37°C with shaking at 150 rpm, the supernatant was collected for enzymatic hydrolysis.
Characteristics of crude glucoamylase
Effect of ph and temperature: The optimum pH for activity was determined by measuring the glucoamylase activity at 55°C using various types of buffers. The following of 0.1 M buffer systems were used: Sodium citrate, ACS reagent grade (pH 3.0), sodium acetate (pH 4.0-5.0), potassium phosphate (pH 6.0-7.0) and tris-HCI (pH 8.0-9.0). All the chemicals used this study was ACS reagent grade. The optimum temperature for activity was assayed by measuring enzyme activity at optimum pH (0.1 M sodium acetate buffer, pH 5.0) over different temperature ranging from 20-90°C. For this method, 1% soluble starch in 0.1 M acetate buffer solution at optimum pH in different tube were incubated over different temperature ranging from 20-90°C for 30 min to make it prewarm.
pH stability and thermostability of crude enzyme: For pH stability, the crude glucoamylase was dispersed (1:1) into 0.1 M buffer solution pH 3.0 (sodium citrate), pH 4.0-5.0 (sodium acetate), pH 6.0-7.0 (potassium phosphate) and pH 8.0-9.0 (Tris-HCI) and incubated at 25°C for 24 h, after that an aliquot was used to determine the remaining activity at optimum pH and temperature. Thermal stability of the crude glucoamylase was determined by incubating in 0.1 M sodium acetate buffer pH 5.0, at 50-70°C for 120 min. Time course aliquots were withdrawn, cooled down in ice bath and assayed under standard conditions at optimum pH (pH 5.0) and temperature (70°C).
Statistical analysis: Data were expressed as Mean±SD for parametric values. Independent sampled t-test was used for normally distributed data. One-way ANOVA was used to measure the group difference and Tukey HSD test was used as significant test. A p-value of <0.05 was considered as significant. All the statistical analysis was performed with the SPSS 16.0 (SPSS Inc, Chicago, IL, USA).
RESULTS
Effect of incubation periods and temperature on glucoamylase production: A temperature range from 20-40°C was used for the production of glucoamylase in submerged fermentation (Fig. 1). In every stage of fermentation for glucoamylase production, triplicate flasks were used to minimize the error. In every stage of fermentation temperature, the enzyme production increased with time interval and decreased after 5 days. The maximum glucoamylase activity (0.77±0.01 U mL –1 min –1) was observed in the culture media after 5 days of incubation periods at 25°C (Fig. 1) but the activity was not significantly higher from other temperatures value (same incubation periods) except at 40°C (p<0.05, by Tukey test).
Effect of initial media pH on glucoamylase production: A range of pH from pH 3.0-8.0 was used for the production of glucoamylase in submerged fermentation to measure the optimum pH for glucoamylase production by A. flavus NSH9. The highest glucoamylase production of 0.77±0.01 U mL –1 min –1 was observed after 5 days of cultivation at pH 5.0, at 25°C (Fig. 2), which was significantly (p<0.05) higher compared to all pH media by Tukey HSD. Glucoamylase production is incerased from pH 3.0-5.0, after that decreased (Fig. 2) and the glucoamylase production was significantly different at different pH media as shown by one way ANOVA (p = 0.000).
Effect of carbon and nitrogen sources on glucoamylase production: Glucoamylase production in different carbon sources such as raw sago starch, soluble starch, raw cassava starch and raw sweet potato starch at various concentrations (1-6%) were observed (Fig. 3).
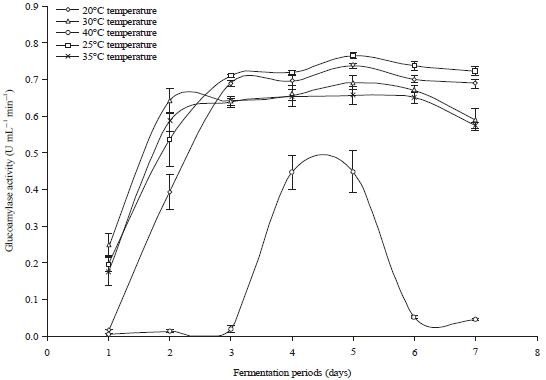 |
| Fig. 1: | Glucoamylase activity in different fermentation periods with temperature at pH 5.0 Error bars show the standard deviation among three independent observations |
 |
| Fig. 2: | Glucoamylase activity at different pH at 25°C and 5 days of Incubation time Error bars show standard deviation among three independent observations |
All the carbon sources supported the enzyme production by the culture media depending on the concentration. The highest glucoamylase activity (0.78±0.01 U mL –1 min –1) was found at 6% soluble starch after 120 h fermentation at 25°C with optimum pH (5.0), followed by 2% raw sago starch (0.77±0.01 U mL –1 min –1), 1% (w/v) raw cassava starch (0.74±0.02 U mL –1 min –1) and 1% (w/v) raw sweet potato starch (0.72±0.02 U mL –1 min –1) (Fig. 3). Glucoamylase activity increased with increasing concentration of soluble starch up to 6%, however in the case of raw cassava and raw sweet potato starch, the productions were lower at higher concentration. In case of raw sago starch glucoamylase activity initially increase up to 2%, after that, at higher concentration impaired the glucoamylase production.
All the nitrogen sources that were tested in this study had on positive effect on glucoamylase production from A. flavus NSH9. Highest glucoamylase production was observed from yeast extract, followed by beef extract, urea, NaN03 and (NH4)2SO4 (Fig. 4). Yeast extract (0.70±0.02 U mL –1 min –1) and urea (0.43±0.06 U mL –1 min –1) were the best organic and inorganic sources of nitrogen, respectively.
Effect of inoculum size on biomass and glucoamylase production: The fungal biomass decreased with higher amount of fungal plugs (Fig. 5). In Fig. 5, it was observed that, negative correlation between biomass production and amount of fungal plugs and the fungal biomass production was significantly different in different inoculum size (p = 0.000). There was significant negative correlation between enzyme activity and fungal biomass (r = -0.644, p = 0.010).
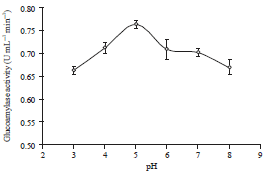
Characteristic of crude glucoamylase: Glucoamylase activity was observed in all pH (pH 3-9) and the values were shown to be significantly different (p = 0.000) via one-way ANOVA (Fig. 6a). The glucoamylase presented optimum activity at a range of pH 4.0-7.0 and the highest activity was at pH 5.0.
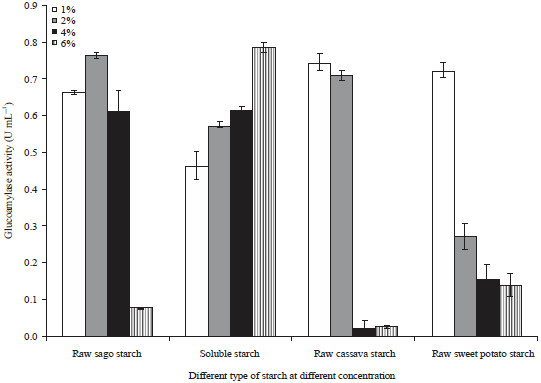 |
| Fig. 3: | Production of glucoamylase by A. flavus NSH9 grown on different types and concentration of starch incubated at 25°C for 5 days. Four types of starch such as raw sago, soluble, raw cassava and raw sweet potato starches (1-6%) were used for fermentation and highest production was at 6% soluble starch Each value represents the mean of three observations with standard deviation |
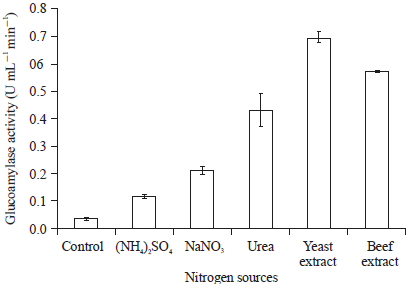 |
| Fig. 4: | Production of glucoamylase by different type of nitrogen at 25°C and 5 day of fermentation Error bars show standard deviation among three independent observations |
After the optimum level of pH, glucoamylase activity gradually declined with higher pH. The value was significantly higher from the activity at all pH (p<0.05, Tukey HSD test) except at pH 4.0.
Glucoamylase activity at different temperatures (20-90°C) was observed and the activity was significantly different (p = 0.000) by one-way ANOVA (Fig. 6b). The highest enzyme activity, 0.83±0.02 U mL –1 min –1, was obtained at 70°C (Fig. 6b) but the value was not significantly higher from temperature 60 and 80°C by Tukey HSD test. The result indicated that, the optimum temperature for glucoamylase activity was from 60-80°C.
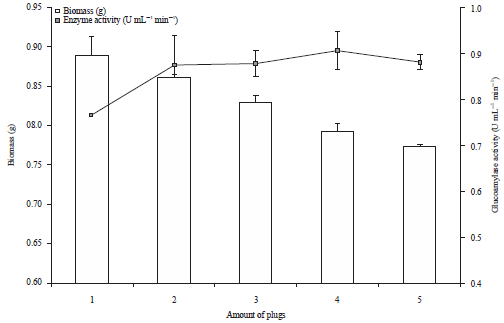 |
| Fig. 5: | Effect of inoculum size on glucoamylase and biomass production Error bars show standard deviation among three independent observations |
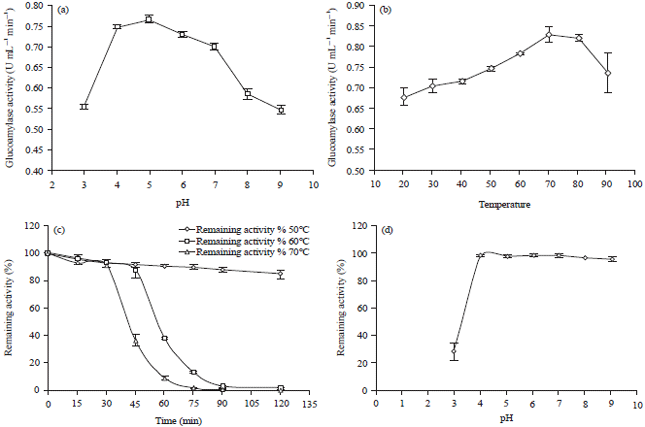 |
| Fig. 6: | Characterization of the crude glucoamylase, (a) Effect of pH on enzyme activity, (b) Effect of temperature on enzyme activity, (c) Thermostability of glucoamylase from 50-70°C, remaining activity was assay at optimum pH (pH 5.0) and temperature (70°C) and (d) pH stability of enzyme after 24 h incubation at 25°C and the assay condition was at optimum pH (pH 5.0) and temperature (70°C). Each value in the panel represents the Mean±SD (n = 3) Error bars show standard deviation among three independent observations |
The glucoamylase from A. flavus NSH9 was stable at 50°C (Fig. 6c). Approximately, 85% residual activity was observed after 120 min at 50°C (Fig. 6c). The glucoamylase activity loss was only about 13% of the initial activity after 45 min at 60°C, after that the activity sharply decreased with increasing time (Fig. 6c). In case of higher temperature at 70°C, more than 90% of residual activity was observed after 30 min (Fig. 6c), after that the activity sharply decreased with increasing time.
Glucoamylase from A. flavus NSH9 was about 95% stable for 24 h at 25°C at pH ranging from 4-9 (Fig. 6d). There were no significant differences in activity at zero (0) min and at 24 h in the different pH buffer solution except at pH 3.0 determined by independent sample t-test (p<0.05).
The raw starch degrading ability of crude glucoamylase was 0.78 U mL –1 h –1 from 1% raw sago starch at 37°C and it was measured at optimum pH (pH 5.0) and temperature (70°C).
DISCUSSION
Optimization of glucoamylase production: Fermentation duration has an important role in substrate utilization and enzyme production13 and also due to the growth phase of microorganism that fluctuate from each other2,14. The highest glucoamylase activity in this study was observed at 5 days of incubation period and many researchers also reported that the optimal production of glucoamylase at 4-5 days of incubation for most of the fungi but depending on substrate types and concentration14,15. These findings in this study were in parallel to previous researchers such as Chimata et al.16, Parbat and Singhal17 and Matthias15 on Aspergillus sp. reported the maximum glucoamylase at the 5th days of incubation. Many researchers supported the finding of optimum fermentation temperature for glucoamylase production of this study. For example, Aspergillus sp. A3 gave GA titres at 30°C under optimum SSF process conditions18, Fusarium solani at (35±1)°C2, 30°C by Alva et al.19 and 35°C for Aspergillus niger20. The finding was in parallel with previous studies on optimum pH for glucoamylase production. In many studies, the highest glucoamylase production was recorded at pH 5.0, such as, from Aspergillus sp.14, from F. solani2, from Aspergillus sp. A318 and from Acremonium sp.21. Hydrogen ion concentration in the fermentation media is one ofthe significant features for microbial growth and secretion of an extracellular enzyme2,14. Sometimes the extracellular enzyme was inactivated in the media due to much fluctuation of pH from its optimal level resulting hamper the substrate utilization22,13. Most of the yeast and fungi needed an acidic environment (pH 4.0-6.5)14 but the optimal production of glucoamylase was observed at pH 4.0-5.0 for many fungi.
The enzyme production is greatly varied depending on type and amount of substrate. When the substrate level was increased beyond a certain limit the enzyme activity started to decrease. This decline may be attributed to partial adsorption of enzyme to the substrate. For example, Matthias15, reported that Rhizopus sp. (RS-6b), Mucor sp. (MS-3) and Aspergillus sp. (AS-2a) can produce maximum glucoamylase with 8% gelatinized potato starches compared to gelatinized cassava, corn, soluble starch and raw starch but Gomes et al.22, reported that cassava starch showed to be a better substrate than corn starch for glucoamylase production by A. flavus A1.1. Yeast extract was the best as organic and urea as inorganic sources of nitrogen among the tested five sources of nitrogen in this study was in agreement with previous study23. Yeast extract has been reported to significantly influence enzyme production in previous studies2,18,24,25, also reported that urea was a good inorganic source of nitrogen both SSF and SmF that is similar this study. Yeast extract was considered as the best, may be due to the significant biomass production compare to other nitrogen sources tested in the study and also it contain amino acids and ammonium ions (NH4)26. A few studies reported that, present of both inorganic and organic nitrogen sources enhanced the raw starch degrading enzyme (RSDE) production21.
Enzyme production and biomass formation are also affected by inoculum density or size of inoculum. Microbial growth and enzyme formation are significantly inhibited by more inoculum size but lower levels take more time for fermentation2,14,27, which is supported by this study. This study supported the finding by Bhatti et al.2, as they reported that enzyme activity increased with an early addition of inoculum size for Fusarium solani under SSF.
Characterization of crude glucoamylase: Fungal glucoamylases are active at acidic condition as reported in previous studies28,29 and the maximum catalytic activity of glucoamylase at pH 5.0 was determined by our finding reinforces the finding in many other studies in A. niger (pH 4.8)30 and in F. solani (pH 4.5)2. The optimal pH value of 4.0-6.0 was recorded from many starch-hydrolysing enzymes from the fungal origin, also similar to this study22,31,32. Many researchers investigated the optimum temperature for glucoamylase activity. For example, 60°C the optimal temperature was observed from A. flavus HBF3429 and 65°C from A. flavus A1.131 for glucoamylase activity. A higher optimum temperature (70°C) of glucoamylase activity reported in this study is similar to the glucoamylase from Thermomyces lanuginosus A13.3722 and Scytalidium thermophilium33. This finding is in contrast with the lower optimum temperature of 40-45°C for glucoamylase from Fusarium solani32 and A. niger34.
Thermostability is the ability of enzyme to resist thermal unfolding in the absence of substrates, while thermophilicity is the capability of enzymes to work at elevated temperatures in the presence of substrate. The crude glucoamylase from A. flavus NSH9 was stable at 50°C temperature. It was observed that F. solani glucoamylase denature rapidly at beyond 60°C32 as they observed the half-life only 26 min at 60°. Compared to other fungal glucoamylases such as from A.citrinus31, F. solani32 and study by Itkor et al.35, glucoamylase from A. flavus NSH9 was more stable at the higher temperature. The stability of the enzyme in a wide range of pH is extremely desirable in the industry29. Meanwhile, pH stability of crude glucoamylase reported in this study was almost similar as reported by many other researchers such as, from A. flavus22 was stable at pH (5.0-9.0), A. flavus29 at pH (pH 3.0-8.0) and other studies28,36.
CONCLUSION
Glucoamylase is an important industrial enzyme in the starch processing industry. In this study, crude glucoamylase production was successfully optimized by employing raw sago starch under submerged fermentation from A. flavus NSH9, and it was at 25°C and at pH 5.0 after 5 days of fermentation. Yeast extract was the best source of nitrogen for glucoamylase production and also highest glucoamylase production was recorded at 4 fungal plugs. Crude GA exhibited optimum catalytic activity at pH 5.0 and temperature of 70°C.
SIGNIFICANCE STATEMENTS
This study discovers the new source of raw starch degrading glucoamylase from Aspergillus flavus NSH9 that can be beneficial for enzyme industry. This study will help the researcher to uncover the critical areas of thermostable raw starch degrading enzyme that many researchers were not able to explore. Thus, this study unveils a new strain, which offers an attractive alternative source of industrial enzyme for starch processing.
REFERENCES
- Mertens, J.A. and C.D. Skory, 2006. Isolation and characterization of a second glucoamylase gene without a starch binding domain from Rhizopus oryzae. Enzyme Microb. Technol., 40: 874-880.
CrossRefDirect Link - Bhatti, H.N., M.H. Rashid, R. Nawaz, M. Asgher, M. Perveen and A. Jabbar, 2007. Optimization of media for enhanced glucoamylase production in solid-state fermentation by Fusarium solani. Food Technol. Biotechnol., 45: 51-56.
Direct Link - Biesbeke, R., E. Record, N. van Bieze, M. Heerikhuisen and A. Franken et al., 2005. Branching mutants of Aspergillus oryzae with improved amylase and protease production on solid substrates. Applied Microbiol. Biotechnol., 69: 44-50.
CrossRefDirect Link - Wang, X.J., J.G. Bai and Y.X. Lian, 2006. Optimization of multienzyme production by two mixed strains in solid-state fermentation. Applied Microbiol. Biotechnol., 73: 533-540.
CrossRefDirect Link - Sindhu, R., G.N. Suprabha and S. Shashidhar, 2009. Optimization of process parameters for the production of a-amylase from Penicillium janthinellum (NCIM 4960) under solid state fermentation. Afr. J. Microbiol. Res., 3: 498-503.
Direct Link - Nguyen, Q.D., J.M. Rezessy-Szabo, M. Claeyssens, I. Stals and A. Hoschke, 2002. Purification and characterisation of amylolytic enzymes from thermophilic fungus Thermomyces lanuginosus strain ATCC 34626. Enzyme Microb. Technol., 31: 345-352.
CrossRefDirect Link - Abdalwahab, S.A., S.A. Ibrahim and E.S. Dawood, 2012. Culture condition forthe production of glucoamylase enzyme by different isolates of Aspergillus spp. Int. Food Res. J., 19: 1261-1266.
Direct Link - Pandey, A., P. Nigam, V.T. Soccol, D. Singh and R. Mohan, 2000. Advances in microbial amylases. Biotechnol. Applied Biochem., 31: 135-152.
PubMedDirect Link - Kumar, P. and T. Satyanarayana, 2009. Microbial glucoamylases: Characteristics and applications. Crit. Rev. Biotechnol., 29: 225-255.
CrossRefDirect Link - Asgher, M., M.J. Asad, S.U. Rahman and R.L. Legge, 2007. A thermostable α-amylase from a moderately thermophilic Bacillus subtilis strain for starch processing. J. Food Eng., 79: 950-955.
CrossRefDirect Link - Leveque, E., S. Janecek, B. Haye and A. Belarbi, 2000. Thermophilic archaeal amylolytic enzyme. Enzyme Microbial Technol., 26: 3-14.
CrossRefDirect Link - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - El-Gendy, M.M.A., 2012. Production of glucoamylase by marine endophytic Aspergillus sp. JAN-25 under optimized solid-state fermentation conditions on agro residues. Aust. J. Basic Applied Sci., 6: 41-54.
Direct Link - Matthias, C.O., 2013. Optimization of α-amylase and glucoamylase production from three fungal strains isolated from Abakaliki, Ebonyi State. Eur. J. Exp. Biol., 3: 26-34.
Direct Link - Chimata, M.K., P. Sasidhar and S. Challa, 2010. Production of extracellular amylase from agricultural residues by a newly isolated Aspergillus species in solid state fermentation. Afr. J. Biotechnol., 9: 5162-5169.
Direct Link - Ellaiah, P., K. Adinarayana, Y. Bhavani, P. Padmaja and B. Srinivasulu, 2002. Optimization of process parameters for glucoamylase production under solid state fermentation by a newly isolated Aspergillus species. Process Biochem., 38: 615-620.
CrossRefDirect Link - Alva, S., J. Anupama, J. Savla, Y.Y. Chiu and P. Vyshali et al., 2007. Production and characterization of fungal amylase enzyme isolated from Aspergillus sp. JGI 12 in solid state culture. Afr. J. Biotechnol., 6: 576-581.
Direct Link - Marlida, Y., N. Saari, Z. Hassan and S. Radu, 2000. Improvement in raw sago starch degrading enzyme production from Acremonium sp. endophytic fungus using carbon and nitrogen sources. Enzyme Microb. Technol., 27: 511-515.
CrossRefDirect Link - Gomes, E., S.R. de Souza, R.P. Grandi and R. da Silva, 2005. Production of thermostable glucoamylase by newly isolated Aspergillus flavus A 1.1 and Thermomyces lanuginosus A 13.37. Braz. J. Microbiol., 36: 75-82.
CrossRefDirect Link - Lagzouli, M., M. Zakaria, A. Ali, W. Senhaji and M. Ouhssine et al., 2007. Optimization of growth and extracellular glucoamylase production by Candida famata isolate. Afr. J. Biotechnol., 6: 2590-2595.
Direct Link - Bertolin, T.E., W. Schmidell, A.E. Maiorano, J. Casara and J.A. Costa, 2003. Influence of carbon, nitrogen and phosphorous sources on glucoamylase production by Aspergillus awamori in solid state fermentation. Zeitschrift fur Naturforschung C, 58: 708-712.
CrossRefDirect Link - Anto, H., U.B. Trivedi and K.C. Patel, 2006. Glucoamylase production by solid-state fermentation using rice flake manufacturing waste products as substrate. Bioresour. Technol., 97: 1161-1166.
CrossRefPubMedDirect Link - Mohamed, L., M. Zakaria, A. Ali, E.K. Youssfi and E. Mohamed et al., 2007. Optimization of composition of media for the production of extracellular glucoamylase by Candida guilliermendii. Pak. J. Biol. Sci., 10: 3322-3329.
CrossRefDirect Link - Norouzian, D., A. Akbarzadeh, J.M. Scharer and M.M. Young, 2006. Fungal glucoamylases. Biotechnol. Adv., 24: 80-85.
CrossRefDirect Link - Koc, Q. and K. Metin, 2010. Purification and characterization of a thermostable glucoamylase produced by Aspergillus flavus HBF34. Afr. J. Biotechnol., 9: 3414-3424.
Direct Link - Jafari-Aghdam, J., K. Khajeh, B. Ranjbar and M. Nemat-Gorgani, 2005. Deglycosylation of glucoamylase from Aspergillus niger: Effects on structure, activity and stability. Biochim. Biophys. Acta (BBA)-Proteins Proteomics, 1750: 61-68.
CrossRefDirect Link - Niaz, M., M.Y. Ghafoor, A. Jabbar, E. Rasul, A. Wahid and R. Ahmed, 2004. Isolation and purification of glucoamylases from Arachniotus citrinus under solid phase growth conditions. Int. J. Biol. Biotechnol., 1: 15-23.
Direct Link - Bhatti, H.N., M.H. Rashid, R. Nawaz, M. Asgher, R. Perveen and A. Jabbar, 2007. Purification and characterization of a novel glucoamylase from Fusarium solani. Food Chem., 103: 338-343.
CrossRefDirect Link - Aquino, A.C.M.M., J.A. Jorge, H.F. Terenzi and M.L. Polizeli, 2001. Thermostable glucose-tolerant glucoamylase produced by the thermophilic fungus Scytalidium thermophilum. Folia Microbiol., 46: 11-16.
CrossRefDirect Link - Itkor, P., O. Shida, N. Tsukagoshi and S. Udaka, 1989. Screening for raw starch digesting bacteria. Agric. Biol. Chern., 53: 53-60.
CrossRefDirect Link - Moreira, F.G., V. Lenartovicz and R.M. Peralta, 2004. A thermostable maltose-tolerant α-amylase from Aspergillus tamari. J. Basic Microbiol., 44: 29-35.
CrossRefDirect Link