
Kamakshi Chaithri Ponnaluri
Department of Genetics, Osmania University, Hyderabad-500007, Andhra Pradesh, India
LiveDNA: 91.12972
Parimala Narne
Department of Genetics, Osmania University, Hyderabad-500007, Andhra Pradesh, India
Mohd. Siraj
Department of Medicine, Deccan College of Medical Sciences, Hyderabad-500058, Andhra Pradesh, India
Mohd. Ishaq
Department of Genetics, Osmania University, Hyderabad-500007, Andhra Pradesh, India
American Journal of Biochemistry and Molecular Biology
Year: 2013 | Volume: 3 | Issue: 1 | Page No.: 159-166







ABSTRACT
Type 2 Diabetes Mellitus (T2DM), an increasingly common serious metabolic disorder has obesity as one of its major predisposing factors. Accumulating evidence suggests that the adipocyte-derived multifunctional hormone, leptin, may play a vital role in the modulation of obesity and subsequent T2DM. The association of polymorphic tetranucleotide repeat (TTTC)n polymorphism in the 3' UTR of the human Leptin gene (LEP-tet) with T2DM was examined in the present study. The study population comprised of 481 subjects including 305 patients and 176 controls. Genotyping of the polymorphic variant was carried out by PCR followed by agarose gel electrophoresis. The alleles of the polymorphism consisted of two groups with different size distributions: the shorter group (class I) and the longer group (class II). Analysis of allele frequencies and genotype frequencies between T2DM subjects and healthy controls revealed no significant differences. Risk estimates for different combinations of the LEP-tet genotypes showed that the I/II genotype was associated with modest 1.46 fold increased risk of T2DM (p = 0.04; 95% CI = 1.01-2.12). A similar trend could not be observed after stratification of the subjects based on sex or obesity. The results obtained in the present study indicated an insignificant influence of the leptin tetranucleotide repeat polymorphism on the development of T2DM. Further studies on a similar aspect would help clarify the issue.
PDF Abstract XML References Citation
Received: March 06, 2012;
Accepted: July 04, 2012;
Published: October 04, 2012
How to cite this article
Kamakshi Chaithri Ponnaluri, Parimala Narne, Mohd. Siraj and Mohd. Ishaq, 2013. Tetranucleotide Repeat Polymorphism in the 3' UTR of the Human Leptin Gene and Risk of Type 2 Diabetes Mellitus in a South Indian Population. American Journal of Biochemistry and Molecular Biology, 3: 159-166.
DOI: 10.3923/ajbmb.2013.159.166
URL: https://scialert.net/abstract/?doi=ajbmb.2013.159.166
DOI: 10.3923/ajbmb.2013.159.166
URL: https://scialert.net/abstract/?doi=ajbmb.2013.159.166
INTRODUCTION
Type 2 diabetes mellitus (T2DM), characterized by chronic hyperglycaemia, is reaching epidemic proportions across the world. The world-wide prevalence of T2DM is estimated to rise to 5.4% by the year 2025 (King et al., 1998), which would account for nearly 300 million affected individuals. Additionally, it has been estimated that the major part of this increase would occur in the countries of India, China and the United States of America. India has the dubious distinction of being the world leader in diabetes with around 40.9 million affected individuals and the figure is expected to rise to 69.9 million by the year 2025, according to the Diabetes Atlas 2006 (Sicree et al., 2006).
Modifications in the dietary patterns owing to urbanization and decreased physical activity coupled to genetic predisposition are considered to be the main drivers of the diabetes epidemic. The substantial contributions of various genetic determinants in the predisposition to T2DM and progression to associated complications underscore the importance of genetic factors in T2DM. Several researchers across the world are involved in identification and characterization of the nature of the genetic variants that predispose humans to T2DM. The results of genome-wide association studies conducted in order to identify susceptibility genes for type 2 diabetes identified Calpain 10 (CAPN10) as a possible diabetogene (Horikawa et al., 2000). However, subsequent studies have not yielded positive associations of T2DM with polymorphic variants of CAPN10 (Hegele et al., 2001; Daimon et al., 2002). Linkage mapping analyses have also helped in the identification of the genes encoding lipin, adiponectin and PTPN1 as candidate genes for T2DM, apart from CAPN10 (Dehwah et al., 2009).
Besides genome-wide association studies, case-control association studies involving candidate gene Polymorphism are being actively carried out across the world. Candidate gene association studies carried out in different parts of the world have identified PPARγ gene as one of the most promising candidates for T2DM (Dehwah et al., 2009). The PPARγ Pro12Ala polymorphic variant has been shown to be strongly associated with T2DM (Dehwah et al., 2008a). Likewise, Polymorphism in the Angiotensin Converting Enzyme and Catalase genes have shown encouraging results in their association with T2DM (Dehwah et al., 2008b; Vasudevan et al., 2008; Dhanapal et al., 2010).
Genes encoding proteins involved in modulating insulin resistance and adiposity are considered to play a key role in the determination of the onset of type 2 diabetes. The gene encoding the adipocytokine, leptin is one such putative candidate gene which holds promise.
Leptin is encoded by the LEP gene, which is present on chromosome 7 (Isse et al., 1995). Leptin is a 16 kD protein that plays a crucial role in regulation of body weight by inhibiting food intake and stimulating energy expenditure. Thus, a fully functional leptin is expected to help individuals maintain ideal body weight without them going into obesity. In fact, in mice, a mutation in the ob gene (mouse homolog of the human LEP gene) has been found to lead to early-onset obesity, causing them to weigh three to four times more than normal mice (Ingalls et al., 1996). These ob/ob mice have leptin deficiency and also exhibit severe insulin resistance and consequent diabetes, suggesting that leptin might play a key role in glucose homeostasis. Additionally, studies on animal models of obesity and diabetes have shown that leptin plays an important role in the maintenance of insulin sensitivity and reversal of the diabetic phenotype (Muzzin et al., 1996; Sivitz et al., 1997; Chinookoswong et al., 1999).
The relationship between serum leptin levels and manifestation of T2DM is more complex in humans with some studies reporting elevated leptin levels being associated with insulin resistance (Segal et al., 1996; Boden et al., 1997; Assal et al., 2007). Leptin in circulation has been demonstrated to exist in free form as well as bound to proteins and quantitative studies have shown that the ratio of free leptin to total leptin correlated positively with BMI, insulin and insulin resistance (Mohiti et al., 2009). Yet others reported lack of any effect of leptin on insulin resistance (Zierath et al., 1998). In humans, a deletion mutation at codon 133 of the LEP gene was found to cause severe leptin deficiency and obesity (Montague et al., 1997). Recently, Shintani et al. (1996), identified a novel microsatellite polymorphism in 3' UTR of the LEP gene (LEP-tet) which was studied for possible association with T2DM and hypertension.
Literature on the relationship between LEP-tet Polymorphism and predisposition to T2DM is sparse in the Indian context owing to which the association of the LEP-tet variant with susceptibility to T2DM has been evaluated in this study.
MATERIALS AND METHODS
The study group comprised 481 unrelated participants of which 305 subjects were with T2DM and 176 were normal healthy controls, who were randomly recruited for the study. None of the controls had any first degree and/or second degree relatives affected with type 1 diabetes, T2DM and hypertension. All the control subjects had fasting glucose <100 mg dL-1 and post lunch blood glucose <140 mg dL-1 and were aged above 50 years. All the T2DM patients were recruited from Kamineni Hospitals, Hyderabad and were diagnosed in accordance with the American Diabetes Association guidelines (American Diabetes Association, 2009). Informed consent was obtained from all the participants of the study and the study was approved by the institutional ethical committee.
Anthropometric data for calculation of Body Mass Index (BMI), clinical information and pedigree details were recorded in a specified proforma. Two milliliter of whole blood through venipuncture was obtained from all participants and genomic DNA was isolated by the rapid salting-out method (Lahiri et al., 1992).
The tetranucleotide repeat (TTTC)n polymorphism in the 3' UTR of the human leptin gene (LEP-tet) was detected by Polymerase Chain Reaction (PCR) as reported previously (Shintani et al., 1996). The primer sequences were as follows: Forward primer-- 5' AGT TCA AAT AGA GGT CCA AAT CA 3' and Reverse primer-- 5' TTC TGA GGT TGT GTC ACT GGC A 3'. The PCR mixture contained 0.1 μg genomic DNA, 25 picomoles of each primer, 0.8 mmol L-1 of each deoxynucleoside triphosphate (Eppendorf, Hamburg, Germany), 1.5 U Taq polymerase (Bangalore Genei, India) and 1X Taq polymerase reaction buffer in a total volume of 25 μL. The PCR cycling reactions were performed in an iCycler machine (BioRad Laboratories, Hercules, California, USA) with an initial denaturation step of 94°C for 5 minutes followed by 35 cycles of 94°C for 30 seconds, 54°C for 30 seconds and 72°C for 1 minute and a final extension of 72°C for 5 minutes. The PCR products were electrophoresed in 2% agarose gels and were visualized by staining with ethidium bromide. The alleles for the LEP-tet polymorphism are classified into two groups: the shorter one (termed class I) with allele size from 150-166 bp and the longer one (termed class II) with allele size from 220-252 bp.
Statistical analyses: Descriptive characteristics of group variables are expressed as Mean±SE. The variation in lipid profiles and other continuous variables between patients and controls was tested for significance using independent sample t-test. Percentage distribution of the LEP-tet genotypes was calculated and the significance was estimated by using the chi-square (χ2) test. The risk estimates for various combinations of genotypes of the LEP-tet polymorphism were obtained by calculating the Odds Ratio (OR) and its 95% CI and p-value. The two tailed p-values for 2x2 contingency tables were calculated using the Fisher’s exact test. All the statistical analyses were conducted using GraphPad Prism version 5.04 for Windows, GraphPad Software, California, USA.
RESULTS
Table 1 summarises the epidemiological and biochemical characteristics of the study subjects. The mean age at sampling of the control subjects was 54.6±1.2 years and was not significantly different (p = 0.19) from that of the patients which was 56.2±0.5 years. The mean age at onset of T2DM in the patient group was 45.8±0.5 years and the mean duration was 10.8±0.4 years. The control subjects had a mean BMI of 27.0±1.2 kg m-2 which was not significantly different from that of the patients which was 27.2±0.3 kg m-2. Significant elevation (p<0.0001) of mean plasma levels of fasting and post lunch blood glucose in the T2DM patients compared with that in the controls was observed.
| Table 1: | Mean distribution of different epidemiological and biochemical parameters observed in T2DM and controls |
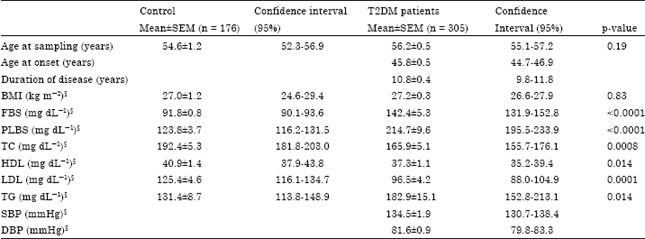 | |
§n: 294 in T2DM group and n: 170 in control group, BMI: Body mass index, FBS: Fasting blood sugar, PLBS: Post lunch blood sugar, TC: Total cholesterol, HDL: High density lipoprotein cholesterol, LDL: Low density lipoprotein cholesterol, TG: Triglycerides, SBP: Systolic blood pressure, DBP: Diastolic blood pressure | |
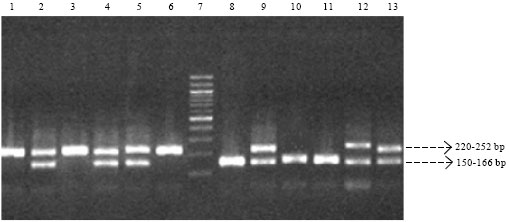 |
| Fig. 1: | Representative gel photograph for the LEP-tet polymorphism, Lane 7: 100 bp DNA ladder; Lanes 8, 10, 11: Genotype I/I, Lanes 2, 4, 5, 9, 12, 13: Genotype I/II, Lanes 1, 3, 6: Genotype II/II |
With respect to the fasting levels of lipids in our study group, we observed that the mean levels of total cholesterol (Controls: 192.4±5.3, T2DM: 165.9±5.1, p = 0.0008) and LDL cholesterol (Controls: 125.4±4.6, T2DM: 96.5±4.2, p = 0.0001) were significantly lower in the T2DM subjects while the mean triglyceride levels (controls: 131.4±8.7, T2DM: 182.9±15.1, p = 0.014) were significantly higher in the T2DM subjects. Mean levels of HDL cholesterol were lower in the diabetic group (37.3±1.1) when compared with that in the controls (40.9±1.4) and this difference was found to be significant statistically (p = 0.014).
Figure 1 depicts the electrophoretic pattern of the PCR amplicons which have been resolved in a 2% agarose gel. The LEP-tet polymorphism comprised two groups-class I and class II based on the number of tandem repeats of the tetranucleotide sequence (TTTC). The results of the genotyping of the LEP-tet polymorphism are presented in Table 2. The frequency of the class I allele was 47.5% in the T2DM group as against the 49.4% in the control subjects. The class II allele showed a frequency of 52.5% in the T2DM group compared with 50.6% in the control group.
| Table 2: | Genotype and allele frequencies of LEP-tet polymorphism in T2DM patients and controls with respect to gender and obesity |
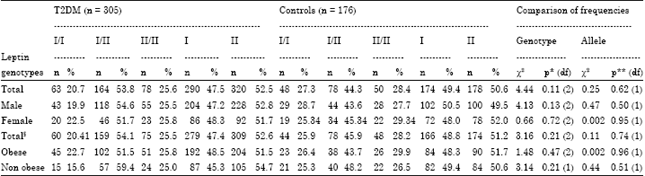 | |
§n: 294 for patients and n: 170 for controls for obesity, *p-value for genotypic frequencies between T2DM and control subjects, **p-value for allele frequencies between T2DM and control subjects, χ2: Male patients vs female patients: 0.31 (p = 0.86), χ2: Obese patients vs. non obese patients: 2.35 (p = 0.31) | |
| Table 3: | Test for deviation from Hardy Weinberg equilibrium |
 | |
nI/I(e): Observed number of subjects with Genotype I/I(expected number), nI/II(e): Observed number of subjects with Genotype I/II (expected number), nII/II(e): Observed number of subjects with Genotype II/II(expected number ), p (Pearson): Pearson's goodness-of-fit chi-square (degree of freedom = 1), p (Llr): Log likelihood ratio chi-square (degree of freedom = 1) | |
| Table 4: | Risk estimations for different combinations of LEP-tet I/II genotypes in T2DM patients and controls |
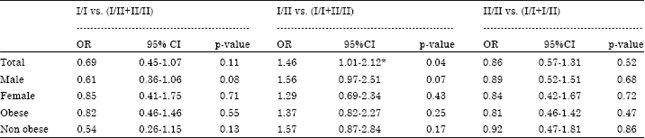 | |
*Statistically significant (p = 0.04) OR and its 95%CI as computed by Fisher’s exact test | |
A perusal of Table 2 reveals that the allele frequencies did not differ significantly (p = 0.62) between the T2DM subjects and control subjects. No significant differences in the allele frequencies between patients and controls were observed even after stratification of the subjects by gender and obesity. As regards the genotype frequencies, the I/II genotype was the most frequent (53.8% in the T2DM group and 44.3% in the control group). The genotype frequencies also did not differ significantly among the total subjects of the two study groups. No significant differences between genotype frequencies were observed even after stratification of the study subjects by obesity and gender. The results of the test for deviation from Hardy-Weinberg Equilibrium are given in Table 3. The genotype frequencies showed no significant deviations from Hardy-Weinberg equilibrium. The risk estimates for type 2 diabetes with different combinations of genotypes (Table 4) reveal that the I/II genotype is showing a modest OR of 1.46 (95% CI: 1.01-2.12, p = 0.04).
DISCUSSION
T2DM is considered to be a complex disorder involving multiple genetic and environmental triggers. Studies across the world are underway to decode the genetic component of type 2 diabetes. Among a vast array of genetic factors responsible for T2DM, those that control adiposity and energy homeostasis play a crucial role in the setting in of insulin resistance and consequent type 2 diabetes. Obesity, which is a result of disturbance of energy homeostasis, namely, imbalance between energy intake and energy expenditure, is characterised by the presence of excess adipose tissue which is a major source of endogenous production of Non Esterified Fatty Acids (NEFAs) through lipolysis. The increased NEFAs in the circulation result in insulin resistance, which can further lead to the development of T2DM (Randle et al., 1963).
The archetypical view of the adipose tissue being an inert storage compartment has changed significantly over the past twenty years. Apart from the secretion of NEFAs, adipocytes are also a source of several metabolically active molecules, termed as adipocytokines or adipokines, such as leptin, adiponectin, resistin, TNF-α etc., most of which play crucial roles in the modulation of energy expenditure and insulin resistance (Cook et al., 1987; Scherer et al., 1995). The recent cloning and characterization of the human homolog of the mouse ob gene, --- the LEP gene --- raised hopes about explaining the possible connection between T2DM and obesity (Zhang et al., 1994). Plasma leptin and adiponectin levels have been reported to influence insulin resistance and progression to T2DM (Daimon et al., 2002; Abd El-Ghaffar and El-Said, 2006; Assal et al., 2007; Mohiti et al., 2009).
Mutations in the mouse ob gene cause severe obesity and T2DM in mice. While mutations in the human LEP gene have not been able to explain the wide prevalence of obesity, common polymorphic variants have been found to be associated with obesity in humans. Common polymorphic variants in the LEP gene have also been reported to influence serum leptin levels, BMI, hypertension and T2DM.
No significant differences in the allele and genotype frequencies between T2DM subjects and healthy controls were observed. A modest risk of 1.46 (p = 0.04) was observed with I/II genotype in the present study which could not be replicated when the study subjects were stratified by obesity and gender. The risk engendered by the LEP-tet polymorphism could be speculated to be the result of alteration of circulating leptin levels by virtue of the polymorphism. Considering that the polymorphism is present in the 3' UTR of the LEP gene, it can be surmised that the expression of the LEP gene and consequent serum leptin levels might be altered due to the polymorphic variant. It is important to remember that in complex traits such as T2DM, the lack of a strong association with a given polymorphic variant does not necessarily preclude the lack of effect of that variant on disease phenotype, because, the susceptibility to disease may be conferred by combinations of variants rather than a single polymorphism.
CONCLUSION
To conclude, individuals heterozygous for the LEP-tet polymorphism possess a statistically significant 1.46 fold risk of T2DM. To the best of our knowledge, this is the first report pertaining to the association of LEP-tet polymorphism with T2DM in a South Indian population. Future studies in different populations involving the LEP-tet polymorphic variant in conjunction with various other polymorphic variants in the LEP gene and the interactions thereof would help in elucidating the role of leptin in the modulation of T2DM.
ACKNOWLEDGMENTS
We thank all the study participants. Kamakshi Chaithri Ponnaluri is a recipient of Senior Research Fellowship from Council of Scientific and Industrial Research, New Delhi, India. We thank University Grants Commission, New Delhi, India for providing Research Fellowship to Parimala Narne.
REFERENCES
- Assal, H.S., M. Fath-Allah and A. Elsherbiny, 2007. Serum leptin and adiponectin in obese diabetic and non-diabetic. J. Med. Sci., 7: 865-869.
CrossRefDirect Link - Boden, G., X. Chen, J.W. Kolaczynski and M. Polansky, 1997. Effects of prolonged hyperinsulinemia on serum leptin in normal human subjects. J. Clin. Invest., 100: 1107-1113.
CrossRefPubMedDirect Link - Chinookoswong, N., J.L. Wang and Z.Q. Shi, 1999. Leptin restores euglycemia and normalizes glucose turnover in insulin-deficient diabetes in the rat. Diabetes, 48: 1487-1492.
CrossRefPubMedDirect Link - Cook, K.S., H.Y. Min, D. Johnson, R.J. Chaplinsky, J.S. Flier, C.R. Hunt and B.M. Spiegelman, 1987. Adipsin: A circulating serine protease homolog secreted by adipose tissue and sciatic nerve. Science, 237: 402-405.
CrossRefPubMedDirect Link - Daimon, M., T. Oizumi, T. Saitoh, W. Kameda and H. Yamaguchi et al., 2002. Calpain 10 gene polymorphisms are related, not to type 2 diabetes, but to increased serum cholesterol in Japanese. Diabetes Res. Clin. Pract., 56: 147-152.
CrossRefPubMedDirect Link - Dehwah, M.A.S., Z. Shuang and Q.Y. Huang, 2008. The association between ACE gene I/D polymorphism and type 2 diabetes in han Chinese in hubei. Int. J. Osteoporosis Metabolic Disorders, 1: 1-7.
CrossRefDirect Link - Dehwah, M.A.S., Z. Shuang, W. Yan, P. Chan and Q.Y. Huang, 2008. Type 2 diabetes in han chinese in hubei. J. Applied Sci., 8: 1235-1241.
CrossRefDirect Link - Dehwah, M.A.S., Z. Shuang, W.Z. Hua, W. Min and Q.Y. Huang, 2009. Type 2 diabetes: Genetic advance and aetiology. J. Applied Sci., 9: 3407-3423.
CrossRefDirect Link - Dhanapal, K., N. Selvan and V. Dhananjeyan, 2010. A study on catalase activity and its genetic polymorphism in diabetes mellitus patients. J. Biol. Sci., 10: 653-657.
CrossRefDirect Link - Abd El-Ghaffar, N. and N.H. El-Said,, 2006. Hypoadiponectinaemia in Egyptian patients with type II diabetes mellitus with vascular complications. J. Med. Sci., 6: 626-630.
CrossRefDirect Link - Hegele, R.A., S.B. Harris, B. Zinman, A.J.G. Hanley and H. Cao, 2001. Absence of association of type 2 diabetes with CAPN10 and PC-1 polymorphisms in Oji-Cree. Diabetes Care, 24: 1498-1499.
CrossRefPubMedDirect Link - Horikawa, Y., N. Oda, N.J. Cox, X. Li and M. Orho-Melander et al., 2000. Genetic variation in the gene encoding calpain-10 is associated with type 2 diabetes mellitus. Nat. Genet., 26: 163-175.
Direct Link - Ingalls, A.M., M.M. Dickie and G.D. Snell, 1996. Obese, a new mutation in the house mouse. Obes. Res., 4: 101-101.
CrossRefPubMedDirect Link - Isse, N., Y. Ogawa, N. Tamura, H. Masuzaki and K. Mori et al., 1995. Structural organization and chromosomal assignment of the human obese gene. J. Biol. Chem., 270: 27728-27733.
CrossRefDirect Link - King, H., R.E. Aubert and W.H. Herman, 1998. Global burden of diabetes, 1995-2025: Prevalence, numerical estimates and projections. Diabetes Care, 21: 1414-1431.
CrossRefPubMedDirect Link - Lahiri, D.K., S. Bye, J.I. Jr. Nurnberger, M.E. Hodes and M. Crisp, 1992. A non-organic and non-enzymatic extraction method gives higher yields of genomic DNA from whole-blood samples than do nine other methods tested. J. Biochem. Biophys. Meth., 25: 193-205.
Direct Link - Mohiti, J., F. Talebi and M. Afkhami-Ardekani, 2009. Circulation free leptin in diabetic patients and its correlation to insulin level. Pak. J. Biol. Sci., 12: 397-400.
CrossRefPubMedDirect Link - Montague, C.T., I.S. Farooqi, J.P. Whitehead, M.A. Soos and H. Rau et al., 1997. Congenital leptin deficiency is associated with severe early-onset obesity in humans. Nature, 387: 903-908.
CrossRefPubMedDirect Link - Muzzin, P., R.C. Eisensmith, K.C. Copeland and S.L. Woo, 1996. Correction of obesity and diabetes in genetically obese mice by leptin gene therapy. Proc. Natl. Acad. Sci., 93: 14804-14808.
Direct Link - Randle, P.J., P.B. Garland, C.N. Hales and E.A. Newsholme, 1963. The glucose fatty-acid cycle its role in insulin sensitivity and the metabolic disturbances of diabetes mellitus. Lancet, 281: 785-789.
CrossRefDirect Link - Scherer, P.E., S. Williams, M. Fogliano, G. Baldini and H.F. Lodish, 1995. A novel serum protein similar to C1q produced exclusively in adipocytes. J. Biol. Chem., 270: 26746-26749.
Direct Link - Segal, K.R., M. Landt and S. Klein, 1996. Relationship between insulin sensitivity and plasma leptin concentration in lean and obese men. Diabetes, 45: 988-991.
CrossRefPubMedDirect Link - Shintani, M., H. Ikegami, E. Yamato, Y. Kawaguchi and T. Fujisawa et al., 1996. A novel microsatellite polymorphism in the human OB gene: A highly polymorphic marker for linkage analysis. Diabetologia, 39: 1398-1401.
CrossRefDirect Link - Sivitz, W.I., S.A. Walsh, D.A. Morgan, M.J. Thomas and W.G. Haynes, 1997. Effects of leptin on insulin sensitivity in normal rats. Endocrinology, 138: 3395-3401.
CrossRefPubMedDirect Link - Vasudevan, R., P. Ismail, J. Stanslas and N. Shamsudin, 2008. Association of G2350A polymorphism of angiotensin converting enzyme gene with essential hypertension and type 2 diabetes mellitus in Malaysian subjects. J. Biol. Sci., 8: 1045-1050.
CrossRefDirect Link - Zhang, Y., R. Proenca, M. Maffei, M. Barone, L. Leopold and J.M. Friedman, 1994. Positional cloning of the mouse obese gene and its human homologue. Nature, 372: 425-432.
CrossRefDirect Link - Zierath, J.R., E.U. Frevert, J.W. Ryder, P.O. Berggren and B.B. Kahn, 1998. Evidence against a direct effect of leptin on glucose transport in skeletal muscle and adipocytes. Diabetes, 47: 1-4.
CrossRefPubMedDirect Link - American Diabetes Association, 2009. Diagnosis and classification of diabetes mellitus. Diabetes Care, 32: S62-S67.
CrossRefDirect Link