
Godswill N. Anyasor
Department of Biochemistry, Ben Carson School of Medicine, College of Health and Medical Sciences, Babcock University, Ilisan-Remo, Ogun State, P.M.B. 21244, Ikeja, Nigeria
LiveDNA: 234.33615
Oluwadamilola M. Odunaike
Department of Biochemistry, Ben Carson School of Medicine, College of Health and Medical Sciences, Babcock University, Ilisan-Remo, Ogun State, P.M.B. 21244, Ikeja, Nigeria
Olufunso O. Olorunsogo
Biomembrane and Biotechnology Laboratories, Department of Biochemistry, Faculty of Basic Medical Sciences, College of Medicine, University of Ibadan, Ibadan, Nigeria
American Journal of Biochemistry and Molecular Biology
Year: 2013 | Volume: 3 | Issue: 1 | Page No.: 110-118







ABSTRACT
This study investigated the cytotoxic effect of varying doses of antimalarial coartemether (2.0, 4.0, 8.0 and 10.0 mg kg-1) in the presence of ferrous sulphate (2800 mg kg-1) for 3 days on normal rat liver mitochondrial membrane permeability transition pore opening and F1F0 ATPase activity. Swelling was estimated spectrophotometrically under succinate energized condition. Calcium ion treated mitochondria preloaded with coartemether induced swelling in a concentration dependent manner in vitro. Swelling was amplified in the presence of ferrous. Coartemether alone and combined coartemether-ferrous also induced mitochondrial swelling in the presence of spermine. In vivo study further showed that 10.0 mg kg-1 coartemether induced swelling in the presence of ferrous. Coartemether stimulated an increased activity of mitochondrial F1F0 ATPase in a concentration dependent manner. Thus, these findings indicate that coartemether at high dose in the presence of ferrous sulphate could be an inducer of mitochondrial mega pore opening and an uncoupler of oxidative phosphorylation initiating apoptosis.
PDF Abstract XML References Citation
Received: November 05, 2011;
Accepted: February 17, 2012;
Published: October 04, 2012
How to cite this article
Godswill N. Anyasor, Oluwadamilola M. Odunaike and Olufunso O. Olorunsogo, 2013. Induction and Uncoupling of Rat Liver Mitochondria by Oral Administered Coartemether. American Journal of Biochemistry and Molecular Biology, 3: 110-118.
DOI: 10.3923/ajbmb.2013.110.118
URL: https://scialert.net/abstract/?doi=ajbmb.2013.110.118
DOI: 10.3923/ajbmb.2013.110.118
URL: https://scialert.net/abstract/?doi=ajbmb.2013.110.118
INTRODUCTION
Coartemether or Coartem® (artemether-lumefantrine) is currently the most viable artemisinin combination therapy specifically indicated for the treatment of acute, uncomplicated malaria infections due to Plasmodium falciparum in patients of 5 kg b.wt. and above (WHO, 2006; Premji, 2009).
Coartemether is a fixed-dose combination tablet of 20 mg artemether and 120 mg lumefantrine in a ratio of 1:6 (Novartis Pharma AG., 2009). One of its components, artemether is a semisynthetic chiral acetal derivative of artemisinin that interferes with parasite transport proteins, disruption of mitochondrial function, inhibits angiogenesis and modulates host immune function. While, lumefantrine is a racemic mixture of a synthetic fluorine derivative formerly known as benflumetol and is structurally related to quinine, mefloquine and halofantrine. It interferes with the conversion of heme, the toxic intermediate produced during hemoglobin break-down to non-toxic hemozoin. The accumulation of heme and free radicals results in parasite death (Byakika-Kibwika et al., 2010).
Artemether, like other artemisinin-derived compounds, acts quickly to rapidly reduce the parasite burden, while lumefantrine serves as a longer-acting agent to eliminate remaining parasites. The combination is effective in parasite strains known to be resistant to traditional antimalarials such as chloroquine (Mwesigwa et al., 2010). Artemether is largely metabolized by cytochrome P450 (CYP) 3A4/5 but also by CYP2B6, CYP2C9 and CYP2C19. Metabolism through CYP3A4 produces an active metabolite, dihydroartemisinin (DHA) that contributes substantially to its antimalarial activity (Cousin et al., 2008). Lumefantrine is metabolized primarily by CYP3A4 and then undergoes glucuronidation (Hietala et al., 2010; Mwesigwa et al., 2010).
The highly reactive endoperoxide moiety in artemisinins is thought to be crucial for their mode of action but the exact mechanism remains controversial (Del Pilar Crespo et al., 2008). Several models have been proposed including a Fenton-type reaction where artemisinins generate reactive oxygen species and carbon-centered radical molecules that modify proteins of Plasmodium parasites. Other studies suggest that artemisinin inhibit the Ca2+-dependent SERCA-like ATPase PfATP6 upon activation by Fe2+ from hemoglobin. Another mechanism is the disruption of the mitochondrial membrane potential as suggested from data of yeast model (Li et al., 2005).
Mitochondrial respiration depends on the flow of electrons through four oligomeric respiratory complexes that comprise the electron transport chain. The energy released by electron flow through the respiratory complexes is conserved in an electrochemical potential consisting of a proton gradient and membrane potential produced by the coupled translocation of protons through the inner mitochondrial membrane at Complexes I, II and IV. Energy stored in the electrochemical potential is coupled to ATP synthesis by translocation of protons into the mitochondrial matrix through complex V (ATP synthase) (Brand and Nicholis, 2011).
The list of described inducers of the Ca2+-dependent mitochondrial mega channel (Membrane Permeability Transition (MPT) pore, is long and includes many different chemical and physical factors all synergistic to Ca2+(Gunter and Pfeifer, 1990; Zoratti and Szabo, 1995). As a result of MPT pore opening intramitochondrial solutes of molecular mass lower than 1.5 kDa equilibrate with those in cytosol (Zoratti and Szabo, 1995). This is accompanied by the activation of mitochondrial respiration, the loss of ions accumulated in the matrix and high amplitude swelling of mitochondria. Recent findings suggest MPT involvement in either programmed mitochondrial destruction (Zorov et al., 1992; Brand and Nicholis, 2011) and hence, in mitochondrial selection (Skulachev, 1996) or in programmed cell death (Zamzami et al., 1995). The mitochondrial contribution to apoptosis opens a vast field for investigating new mitochondrial related diseases.
Several studies have tested coartemether and its components in a complete range of acute and subchronic animal toxicology studies, including reproductive toxicology, genotoxicity and juvenile animal studies (Raji et al., 2005; Efferth and Kaina, 2010; Onyesom and Agho, 2011). Mechanistic neurotoxicity studies were performed in both rats and dogs to evaluate functional and histopathologic changes (Oyemitan et al., 2007; Cousin et al., 2008; Ajibade et al., 2011). Despite these findings there is a paucity of large-scale clinical trials suitable to detect rare but significant toxicity especially when artemisinin combine therapy is prescribed alongside with ferrous in severe anemia conditions. Although, Mpiana et al. (2007) have reported that endoperoxide lactone based drugs could form complexes with heme and hemin. Therefore, attempts had been made to further investigate the effect of cytotoxic effect of coartemether on mitochondrial integrity.
MATERIALS AND METHODS
Chemicals: Coartemether tablets (Norvatis, Pharma AG, Switzerland), ferrous sulphate tablets (Pharmacy unit, University College of Health, Ibadan, Nigeria), mannitol, sucrose (BDH Chemicals Ltd; Pools, England), HEPES (May and Baker Lab; USA), EGTA, bovine serum albumin (Sigma Chemical Co; USA), spermine (Research Biochemical, USA) and all other reagents used were of analytical grade.
Animal: Male albino Wistar rats (120-150 g) were obtained from Preclinical Animal House, Physiology Department, University of Ibadan, Ibadan, Nigeria. The animals were maintained in cages acclimatized for two weeks in accordance to good laboratory animal care practice at the departmental animal house. Tap water and commercial pelleted feed were provided under standard conditions of temperature 28±2°C and a 12 h light/dark cycle.
Experimental design: Oral coartemether tablets were dissolved in sunflower oil as vehicle and administered orally to the test groups using an oral dosing needle for 3 days in accordance to WHO recommendation for treatment of uncomplicated malaria. Animals were assigned randomly into six groups of three rats each. Group I: untreated normal rats; group II: (control) normal rats were given oral sunflower oil (1 cm3), group III-VI were coadministered coartemether (2.0, 4.0, 8.0 and 10.0 mg kg-1 body weight) and ferrous sulphate (4.3 mg kg-1 b.wt.) respectively. The animals were euthanized 24 h after fasting overnight by cervical dislocation at the end of treatment. Subsequently, the rat liver was excised, trimmed of excess tissue and subjected through the standard protocol for mitochondria isolation. Mitochondria isolated from Group I were preloaded with varying concentrations of coartemether (600, 1200, 2400 and 3000 μg mL-1) and ferrous sulphate (1280 μg mL-1) to assess the in vitro effects on mitochondria MPT.
Mitochondrial isolation: Rat liver mitochondria isolated by conventional differential centrifugation in a buffer containing 210 mM mannitol, 70 mM sucrose, 5 mM HEPES (pH 7.4) and 1 mM EGTA (Schneider and Hogeboom, 1950); EGTA was omitted in the final wash solution. Protein content was estimated by Folin-Ciocalteu method using Bovine Serum Albumin as standard (Lowry et al., 1951).
Assessment of mitochondrial swelling: Mitochondrial swelling was assessed according to the method of Lapidus and Sokolove (1993). Changes in absorbance of mitochondria were monitored at 540 nm in a 6405 Jenway UV-visible spectrophotometer. Mitochondria (0.4 mg mL-1) were suspended in a medium containing 210 mM mannitol, 70 mM sucrose, 5 mM HEPES-KOH (pH 7.4), 0.8 μM rotenone and 5 mM succinate. Swelling was triggered by Ca2+ while spermine serves as an inhibitor.
Assessment of mitochondrial F1F0 ATPase activity: Mitochondria isolated from untreated normal rats in another set of experiment were preloaded with varying concentrations of 10.0, 20.0, 30.0, 40.0, 50.0 and 60.0 mg mL-1 coartemether to assess the effects on mitochondrial F1F0 ATPase activity. Mitochondrial adenosine triphosphatase or F1F0 ATPase activity was determined by a modified method of Lardy and Wellman (1953). Each reaction vessel contained 65 mM tris-HCl (pH 7.4), 1 mM ATP and 25 mM sucrose. The reaction was started by the addition of mitochondrial fraction (0.4 mg mL-1) vortex for 30 min at 25°C. The reaction was stopped by the addition of 8 mL of 10% trichloroacetic acid to each test tube then centrifuged at speed of 3000 g. The deproteinized supernatant was kept for phosphate determination.
Determination of inorganic phosphate: This was performed according to the method described by Fiske and Subbarow (1925) modified by Bababunmi and Bassir (1972). 0.4 mL of perchloric acid was added to 5.0 mL of the deproteinized supernatant in a test tube. This was followed by addition of 0.4 mL of 5% ammonium molybdate and 0.2 mL of a 0.2% freshly prepared solution of ascorbic acid. The tube was thoroughly mixed, gently shaken and allowed to stand for 20 min. A standard solution of potassium dihydrogen phosphate (0.2 mg Pi per 5 mL) was similarly treated. The intensity of the blue colour which developed was read at 680 nm using a spectrophotometer. Water blank was used to set the instrument at zero.
Calculation:
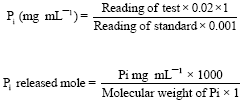 |
Mole Pi released mL-1 mitochondrial protein is given by the expression:
RESULTS
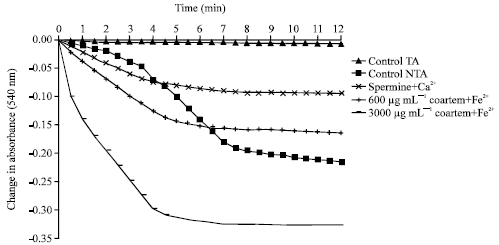
Triggering agent (Ca2+) induced MPT or swelling in a succinate energized rat liver mitochondria under a normal respiration sucrose-phosphate buffer. However, mitochondrial membrane was intact without any observable swelling in the absence of a triggering agent (Fig. 1). In vitro study showed that mitochondria preloaded with 600, 1200, 2400, 3000 μg mL-1 coartemether induced opening of the MPT pore in a concentration dependent manner with minimal and maximal swelling inductions at 600 and 3000 μg mL-1 coartemether, respectively. Conversely, ferrous sulphate at 1280 μg mL-1 did not induced mitochondria swelling (Fig. 1). However, coartemether preloaded mitochondria in the presence of ferrous showed a significantly large amplitude (p<0.05) of swelling in a concentration dependent manner (Fig. 2).
Spermine inhibited swelling induced at low concentration of coartemether (600 μg mL-1) and ferrous while high concentration of coartemether (3000 μg mL-1) and ferrous induced mitochondria swelling in the presence of spermine (Fig. 3).
In the animal study, co-administered ferrous sulphate (4.3 mg kg-1) and coartemether at 2.0, 4.0, 8.0 mg kg-1 did not induce mitochondria MPT both in the absence Ca2+. However, there was significantly high (p<0.05) amplitude of liver mitochondria swelling in the animals co-administered with 10 mg kg-1 coartemether and ferrous sulphate (4.3 mg kg-1) (Fig. 4).
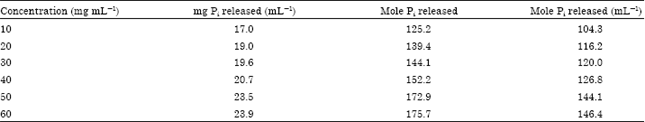
The result also revealed that varying concentrations of coartemether (10, 30, 50 and 60 mg mL-1) elevated the mitochondrial F1F0 ATPase activity in a concentration dependent manner with minimal (104.4 mol Pi/mg protein/ min) and maximal (146.4 mol Pi /mg protein/ min) activities at 10 and 60 mg mL-1, respectively (Table 1).
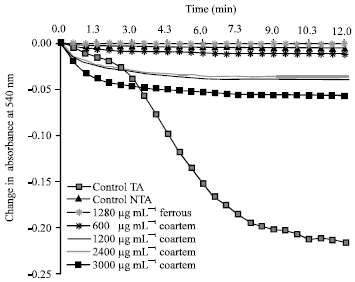 |
| Fig. 1: | Change in absorbance (540 nm) for 12 min by ferrous sulphate (Fe2+), varying concentrations of coartemether on mitochondrial membrane permeability transition pore in the energized by sodium succinate, TA: Triggering agent, NTA: Non triggering agent |
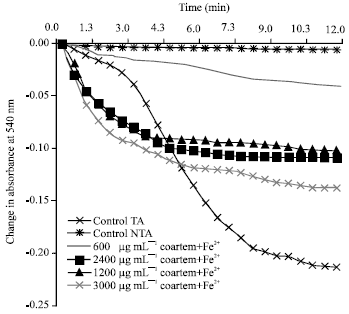 |
| Fig. 2: | Change in absorbance (540 nm) for 12 min by varying concentrations of coartemether in the presence of ferrous sulphate on mitochondrial membrane permeability transition pore energized by sodium succinate, TA: Triggering agent, NTA: Non triggering agent |
| Table 1: | The Hydrolysis of ATP by mitochondrial F1F0 ATPase by varying concentrations of coartemether |
 | |
 |
| Fig. 3: | Change in absorbance (540 nm) for 12 min by two concentrations of coartemether (lowest and highest) combined with ferrous sulphate on mitochondria permeability transition pore in the presence of spermine energized by sodium succinate, TA: Triggering agent, NTA: Non triggering agent |
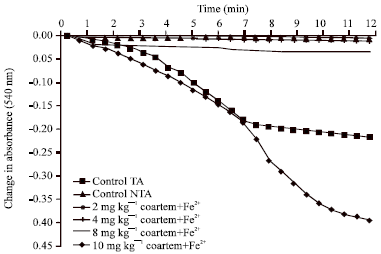 |
| Fig. 4: | Change in absorbance (540 nm) for 12 min in vivo effect of co-administered Fe2+ (4.3 mg kg-1) and varying dose of coartemether on mitochondria permeability transition pore in the absence of a triggering agent energized by sodium succinate, TA: Triggering agent, NTA: Non triggering agent |
DISCUSSION
In the present study, rat liver mitochondria preloaded with varying concentrations of coartemether in vitro induced mitochondrial Membrane Permeability Transition (MPT) pore in a concentration dependent manner. This supports the theory implicating mitochondrial MPT pore formation (swelling) as the possible mode of action for artemisinin-related compounds (Eckstein-Ludwig et al., 2003; Wang et al., 2010). Studies had shown that the parasiticidal activity of artemether resides on the peroxyl ring of dihydroxyartemisinin metabolite (Krishna et al., 2004; Efferth and Kaina, 2010). The cleavage of an endoperoxide bridge generates free radicals (Cumming et al., 1997). Free radicals have been associated with mitochondrial MPT pore opening (Del Pilar Crespo et al., 2008). Mitochondrial membrane permeabilization results in the release of cytochrome c that trigger apoptosis by caspase-cascade pathways, consequently leading to cell death (Galluzi et al., 2009). Furthermore, the addition of varying concentrations of coartemether to the calcium ion preloaded mitochondria in the presence of ferrous sulphate further increased the swelling of mitochondria. Previous studies reported that iron bioactivates artemisinin into a free radical through an iron-mediated cleavage (Dhingra et al., 2000; Balint, 2001; Noori et al., 2004). This study also showed that coartemether in the presence of iron could serve as a cytotoxic agent against cancerous cells. Other studies had proved that the combined administration of dihydroxyartemisinin and iron retarded the growth rate of tumor (Moore et al., 1995).
The induction of mitochondrial MPT by coartemether at high concentrations in the presence of a natural inhibitor spermine in vitro suggests that the cytotoxic effect of coartemether could be at high concentrations.
Animal study showed that coartemether at the therapeutic dose range of 2-4 mg kg-1 b.wt. did not induce mitochondrial swelling in the absence of a calcium ion. This seems to be in agreement with Norvatis Pharma claim on the clinical safety of the therapeutic dose range (Novartis Pharma AG., 2009). This observation might have resulted from the action of CYP3A4 isoenzyme metabolizing and degrading the total amount of drug that reaches the receptor/active site to elicit a pharmacological response (Cousin et al., 2008; Mwesigwa et al., 2010). However, swelling was induced at high dose of 10 mg kg-1 b.wt. coartemether.
Further studies indicated that coartemether elevated the hydrolysis of ATP to ADP and inorganic phosphate by mitochondrial F1F0 ATPase in a concentration dependent manner. This was determined spectrophotometrically by the increase in concentration of released inorganic phosphate Pi. This observation could also be accounted through the induction of mitochondrial MPT by coartemether in vitro which have already compromised the intactness of the mitochondria. The mitochondrial F1F0 ATPase or ATP synthase is known to harnesses the proton gradient generated during the transfer of electron along the respiratory chain and couples it to the oxidative phosphorylation of ADP and inorganic phosphate to produce ATP required for diverse biochemical and cellular functions (Vinogradov, 2000; Nelson and Cox, 2008). The collapse in mitochondrial electrochemical gradient could result in the hydrolysis of ATP by F1F0-ATPase for the proton gradient recovery. This turns F1F0-ATPase into a consumer rather than being a producer of ATP in failing cells (Zablockaite et al., 2007; Nelson and Cox, 2008).
CONCLUSION
This study indicates that coartemether at high dose in the presence of iron could have a profound cytotoxic effect on mitochondrial membrane permeability transition pore and it also could serve as an uncoupler of mitochondria respiration. However, additional studies are required to probe further into the effect of coartemether on cytochrome c release and mechanisms associated with apoptosis to gain more insight into the toxicological pathway.
REFERENCES
- Ajibade, A.J., P.B. Fakunle and P.D. Shallie, 2011. Some histological observations and microstructural changes in the nissl substances in the cerebellar cortex of adult wistar rats following artesunate administration. Curr. Res. Neurosci., Vol.2.
Direct Link - Bababunmi, E.A. and O. Bassir, 1972. Effects of aflatoxin B1 on the swelling and adenosine triphosphatase activities of mitochondria isolated from different tissues of the rat. FEBS Lett., 26: 102-104.
Direct Link - Balint, G.A., 2001. Artemisinin and its derivative: An important new class of antimalarial agents. Pharmacol. Ther., 90: 261-265.
CrossRefDirect Link - Brand, M.D. and D.G. Nicholis, 2011. Assessing mitochondrial dysfunctions in cells. Biochem. J., 435: 297-312.
CrossRefDirect Link - Byakika-Kibwika, P., M. Lamorde, H. Mayanja-Kizza, C. Merry, B. Colebunders and J. van Geertruyden, 2010. Update on the efficacy, effectiveness and safety of artemether-lumefantrine combination therapy for treatment of uncomplicated malaria. Ther. Clin. Risk Manage., 6: 11-20.
CrossRefDirect Link - Cumming, J.N., P. Ploypradith and G.H. Posner, 1996. Antimalarial activity of artemisinin (qinghaosu) and related trioxanes: Mechanism(s) of action. Adv. Pharmacol., 37: 253-297.
CrossRefPubMedDirect Link - Dhingra, V., K.V. Rao and M.L. Narasu, 1999. Current status of artemisinin and its derivatives as antimalarial drugs. Life Sci., 66: 279-300.
CrossRefPubMedDirect Link - Eckstein-Ludwig, U., R.J. Webb, I.D.A. van Goethem, J.M. East and A.G. Lee et al., 2003. Artemisinins target the SERCA of Plasmodium falciparum. Nature, 424: 957-961.
CrossRefDirect Link - Efferth, T. and B. Kaina, 2010. Toxicity of the antimalarial artemisinin and its derivative. Crit. Rev.Toxicol., 40: 405-421.
CrossRefPubMedDirect Link - Fiske, C.H. and Y. Subbarow, 1925. The colorimetric determination of phosphorus. J. Biol. Chem., 66: 375-400.
CrossRefDirect Link - Galluzi, L., E. Morselli, O. Kepp and G. Kroemer, 2009. Targeting post mitochondrial effectors of apoptosis for neuroprotection. Biochem. Biophys. Acta Bioenergetics, 1787: 402-413.
Direct Link - Gunter, T.E. and D.R. Pfeifer, 1990. Mechanisms by which mitochondria transport calcium. Am. J. Physiol. Cell Physiol., 258: C755-C786.
CrossRefDirect Link - Hietala, S.F., A. Martensson, B. Ngasala, S. Dahlstrom and N. Lindegardh et al., 2010. Population pharmacokinetics and pharmacodynamics of artemether and lumefantrine during combination treatment in children with uncomplicated falciparum malaria in Tanzania. Antimicrob. Agents Chemother., 54: 4780-4788.
Direct Link - Krishna, S., A.C. Uhlemann and R.K. Haynes, 2004. Artemisinins: Mechanism of action and potential for resistance. Drug Resist. Updates, 7: 233-244.
CrossRefPubMedDirect Link - Lapidus, R.G. and P.M. Sokolove, 1993. Spermine inhibition of the permeability transition of isolated rat liver mitochondria: An investigation of mechanism. Arch Biochem. Biophys., 306: 246-253.
CrossRefPubMedDirect Link - Lardy, H.A. and H. Wellman, 1953. The catalyst effect of 2,4 dinitrophenol on adeno-sinetriphosphate hydrolysis by cell particles and soluble enzymes. J. Biol. Chem., 201: 357-370.
Direct Link - Li, W., W. Mo, D. Shen, L. Sun and J. Wang et al., 2005. Yeast model uncovers dual roles of mitochondria in action of artemisinin. PLoS Genet., Vol. 1.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Moore, J.C., H. Lai, J.R. Li, R.L. Ren, J.A. McDougall, N.P. Singh and C.K. Chou, 1995. Oral administration of dihydroartemisinin and ferrous sulfate retarded implanted fibrosarcoma growth in the rat. Cancer Lett., 98: 83-87.
PubMed - Mpiana, P.T., B.K. Mavakala and Y. Zhi-Wu, 2007. Interaction of artemisinin based antimalarial drugs with hemin in water-DMSO mixture. Int. J. Pharmacol., 3: 302-310.
CrossRefDirect Link - Mwesigwa, J., S. Parikh, B. McGee, P. German and T. Drysdale et al., 2010. Pharmacokinetics of artemether-lumefantrine and artesunate and amodiaquine in Children in Kampala, Uganda. Antimicrob. Agents Chemother., 54: 52-59.
CrossRefDirect Link - Noori, S., G.A. Naderi, Z.M. Hassan, Z. Habibi, S.Z. Bathaie and S.M.M. Hashemi, 2004. Immunosuppresive activity of a molecule isolated from Artemisia annua on DTH responses compared with cyclosporine A. Int. Immunopharmacol., 4: 1301-1306.
Direct Link - Oyemitan, I.A., E.O. Iwalewa, O.E. Ukponmwan, R.O. Akomolafe and O.M. Daniyan, 2007. The involvement of serotonin in artemether-induced behavioural activities. J. Biol. Sci., 7: 575-578.
CrossRefDirect Link - Onyesom, I. and J.E. Agho, 2011. Changes in serum glucose and triacylglycerol levels induced by the co-administration of two different types of antimalarial drugs among some Plasmodium falciparum malarial patients in Edo-delta Region of Nigeria. Asian J. Sci. Res., 4: 78-83.
CrossRefDirect Link - del Pilar Crespo, M., T.D. Avery, E. Hanssen, E. Fox and T.V. Robinson et al., 2008. Artemisinin and a series of novel endoperoxide antimalarials exert early effects on digestive vacuole morphology. Antimicrob. Agents Chemother., 52: 98-109.
CrossRefDirect Link - Raji, Y., I.O. Osonuga, O.S. Akinsomisoye, O.A. Osonuga and O.O. Mewoyeka, 2005. Gonadotoxicity evaluation of oral artemisinin derivative in male rats. J. Med. Sci., 5: 303-306.
CrossRefDirect Link - Schneider, W.C. and G.H. Hogeboom, 1950. Intracellular distribution of enzymes. V. Further studies on the distribution of cytochrome c in rat liver homogenate. J. Biol. Chem., 183: 123-128.
CrossRefDirect Link - Skulachev, V.P., 1996. Role of uncoupled and non-coupled oxidations in maintenance of safely low levels of oxygen and its one-electron reductants. Q. Rev. Biophys., 29: 169-202.
CrossRefPubMedDirect Link - Zoratti, M. and I. Szabo, 1995. The mitochondrial permeability transition. Biochim. Biophys. Acta (BBA)-Rev. Biomembr., 1241: 139-176.
CrossRefDirect Link - Wang, J., L. Huang, J. Li, Q. Fan, Y. Long, Y. Li and B. Zhou, 2010. Artemisinin directly targets malarial mitochondria through its specific mitochondrial activation. PLoS ONE, Vol. 5.
CrossRefDirect Link - Zablockaite, D., V. Gendviliene, I. Martisiene and J. Jurevicius, 2007. Effect of oxidative phosphorylation uncoupler FCCP and F1F0-ATPase inhibitor oligomycin on the electromechanical activity of human myocardium. Adv. Med. Sci., 52: 89-93.
Direct Link - Zamzami, N., P. Marchetti, M. Castedo, C. Zanin, J.L. Vayssiere, P.X. Petit and G. Kroemer, 1995. Reduction in mitochondrial potential constitutes an early irreversible step of programmed lymphocyte death in vivo. J. Exp. Med., 181: 1661-1672.
CrossRefDirect Link - Zorov, D.B., K.W. Kinnally and H. Tedeschi, 1992. Voltage activation of heart inner mitochondrial membrane channels. J. Bioenerg. Biomembr., 24: 119-124.
Direct Link - Cousin, M., S. Kummerer, G. Lefevre, A.C. Marrast, D. Stein and M. Weaver, 2008. Coartem (artemether-lumefantrine) tablets for the treatment of malaria in patients with acute uncomplicated infections due to plasmodium falciparum or mixed infections including ip. Falciparum. Novartis, Africa, pp: 14-20.