
Abdullah-Al- Mahin
Laboratory of Microbial Technology, Division of Microbial Science and Technology, Department of Bioscience and Biotechnology, Faculty of Agriculture, Graduate School, Kyushu University, 6-10-1 Hakozaki, Higashi-ku, Fukuoka 812-8581, Japan
LiveDNA: 880.1542
ORCID: 0000-0001-6801-9910
Kenji Sonomoto
Laboratory of Microbial Technology, Division of Microbial Science and Technology, Department of Bioscience and Biotechnology, Faculty of Agriculture, Graduate School, Kyushu University, 6-10-1 Hakozaki, Higashi-ku, Fukuoka 812-8581, Japan
American Journal of Biochemistry and Molecular Biology
Year: 2012 | Volume: 2 | Issue: 3 | Page No.: 157-166







ABSTRACT
The effect of different concentrations of nisin on DnaK-overproducing Lactococcus lactis strains while growing at 40°C was examined. The plasmid pNZ-8048 which has a nisin promoter was used as a vector for transformation and expression of heterologous or homologous DnaK into L. lactis NZ9000. The transformants were then induced with different concentration of nisin and allowed to grow in presence of nisin. It was found that nisin, the antimicrobial peptide used for induction of DnaK-overexpression, itself conferred a lower but significant stress to the strains. Escherichia coli DnaK (DnaKEco)-overexpressing cells showed better tolerance to nisin than L. lactis DnaK (DnaKLla) and T. halophilus DnaK (DnaKTha)-overexpressing cells. However, all DnaK-overexpressing strains showed better tolerance than the cells without DnaK overexpression. These findings suggest the possibility for a relationship of DnaK protein and tolerance to different antimicrobial peptides.
PDF Abstract XML References Citation
Received: September 06, 2011;
Accepted: December 23, 2011;
Published: February 21, 2012
How to cite this article
Abdullah-Al- Mahin and Kenji Sonomoto, 2012. Nisin Tolerance of DnaK-overexpressing Lactococcus lactis Strains at 40°C. American Journal of Biochemistry and Molecular Biology, 2: 157-166.
DOI: 10.3923/ajbmb.2012.157.166
URL: https://scialert.net/abstract/?doi=ajbmb.2012.157.166
DOI: 10.3923/ajbmb.2012.157.166
URL: https://scialert.net/abstract/?doi=ajbmb.2012.157.166
INTRODUCTION
L. lactis, a spherical-shaped gram-positive lactic acid bacterium, has been used for long time for industrial production of fermented dairy products such as milk, cheese and yogurt. Microbial biomass of Lactic Acid Bacteria (LAB) are also widely used in the food and pharmaceutical industries (Lee et al., 2007). Because of the importance of lactic acid bacteria in food industry extensive research has been done on their metabolic pathway to increase its efficiency for fermentation. Recently, the bacterial group is getting much attention even in medical science (Anukam, 2007). Food supplemented with LAB have been shown to work against cancer development (Gursoy and Kinik, 2006). Some of their special characteristics such as faster growth, acidification and resistance to bacteriocin made them suitable for selection in the dairy industries (Van de Guchte et al., 2002; Yateem et al., 2008; Ali, 2011). On the other hands, these strains need to overcome the stress conditions encountered in the fermentation processes for better productivity. Many efforts have been made to overcome these problems. With the recent developments of molecular genetics and proteomics, some genes are figured out to be involved in stress responses of LAB, other bacteria and mammalian cells (Abdullah-Al-Mahin et al., 2010; Fiocco et al., 2007; Prasad et al., 2003; Walker et al., 1999; Walter et al., 2003; Kullen and Klaenhammer, 1999; Lentze and Narberhaus, 2005; Ahmed, 2006).
Hsp70s play a very important role for folding of newly synthesized proteins, refolding of misfolded proteins and transportation of proteins through biological membranes both under normal and stressed conditions (Mayer et al., 2001). DnaK, a 70-kDa heat shock protein homolog in bacteria, transduces signals to other cellular factors when a shift of temperature increase occurs (Craig and Gross, 1991). The hsp70 genes are widely conserved in all organisms except some archaeal strains. However, the functional studies of this heat responsive gene were widely carried out with Saccharomyces cerevisiae, E. coli and Bacillus subtilis (Homuth et al., 1997; Schulz et al., 1995). DnaK functions in cooperation with DnaJ and GrpE and the complex of DnaK-DnaJ-GrpE plays a significant role in the refolding of thermally damaged proteins. However, the complex also assists in the folding of nascent protein chains under normal growth conditions (Bukau, 1993; Bukau and Walker, 1989, 1990; Cegielska and Georgopoulos, 1989). Liberek et al. (1991) reported that DnaK requires ATP for its activity in vitro and this ATPase activity is stimulated by its co-chaperone DnaJ and GrpE. Like other bacteria, L. lactis also exhibits a heat shock response in which molecular chaperones play key roles (Arnau et al., 1996; Kilstrup et al., 1997; Whitaker and Batt, 1991). Previously, overproductions of GroEL/ES and small Hsps have been reported to improve the stress tolerance of lactic acid bacteria (Desmond et al., 2002, 2004). We reported earlier that both heterologous and homologous expression of DnaK in L. lactis improves heat, salt, ethanol and acid (low pH)-stress. We also found that nisin, the antimicrobial peptide used for induction of DnaK-overexpression also conferred stress both at normal physiological conditions and high temperature (Abdullah-Al-Mahin et al., 2010). Therefore in this report, we aimed to investigate more in detail the effect of DnaK-overexpression in L. lactis strains to nisin when growing at 40°C.
MATERIALS AND METHODS
Bacterial strains and media: L. lactis subsp. cremoris NZ9000 and E. coli JM109 were used throughout the study. L. lactis NZ9000 was grown in GM17 medium (M17 broth supplemented with 0.5% glucose) at 30°C, unless stated otherwise. E. coli JM109 was grown aerobically in Luria-Bertani (LB) broth at 37°C, unless stated otherwise. T. halophilus JCM5888 was grown in MRS medium (Oxoid, Hampshire, UK) containing 1 M NaCl at 30°C. The medium was adjusted to pH 7.5 before sterilization. In order to facilitate clonal selection, 5 μg mL-1 chloramphenicol was added to the media.
Construction of DnaK-expressing L. lactis: Chromosomal DNA was isolated from E. coli JM109, L. lactis NZ9000 and T. halophilus JCM5888 using combination of two methods as described earlier (Berns and Thomas, 1965; Marmur, 1961). The DnaKEco gene was amplified from E. coli chromosome using primers 5’-CCCCTATTAGGATCCCACAACCACATGATGACCGAATATAT-3’ and 5’-GTCAGTATAATTACCCGTTTATAGAGCTCTTATTT-3’. The BamHI and SacI sites were simultaneously inserted into the amplified DnaKEco gene. A BamHI restriction endonuclease site was inserted into the plasmid pNZ8048 (De Ruyter et al., 1996) by inverse PCR using primers 5’-CTAGAGAGCTCAAGCTTTCTTTGAACCAAA-3’ and 5’-TTTTGTGGATCCTTTCGAACGAAATC-3’. The DnaKLla was amplified from L. lactis NZ9000 using primers 5’-ATATTGACCGCCATGGCTTTAAACTATTC-3’ and 5’- ACTGACGAAACGATGAGCTCTTTTTTAAA-3’, The NcoI and SacI sites were simultaneously inserted into the amplified DnaKLla gene. The DnaKTha gene was amplified from T. halophilus JCM5888 chromosome using primers 5’-AGATCAATATCATGAGTAAGATAATTGGTATTGACT-3’ and 5’-ATTTCCCAAATAGAGCTCTTATTGATTATCGTT-3’. The PagI and SacI sites were simultaneously inserted into the amplified DnaKTha gene. PCR was performed with KOD plus Dna polymerase (Toyobo, Osaka, Japan). The amplified PCR products were purified with the QIAquick PCR Purification Kit (Qiagenne West Sussex, United Kingdom). All the amplified DnaK genes and the plasmid were digested with their respective restriction enzymes. For insertion of DnaKLla and DnaKTha the plasmid pNZ8048 was enzymatically digested with NcoI and SacI restriction enzymes. Enzymatically digested products were then ligated using Ligation High ver. 2 (Toyobo), according to the manufacturer’s instructions. The resulting plasmids which contained DnaKEco DnaKLla and DnaKTha were named as pNZ-EDnaK, pNZ-LDnaK and pNZ-TDnaK. All these three plasmids were then transformed into L. lactis NZ9000, according to the method developed by Holo and Nes (Holo and Nes, 1989) and the transformants were designated as NZ-EDnaK, NZ-LDnaK and NZ-TDnaK, respectively. An empty plasmid pNZ8048 were also transformed into L. lactis NZ9000 and the transformant was named as NZ-Vector which is used as the control strain throughout the study.
Overexpression of DnaK and investigation of stress tolerance: All the transformants were grown in GM17 medium (10 mL) containing 5 μg mL-1 chloramphenicol at 30°C. When optical density at 600 nm (OD600) reached at 0.5-0.6, the expression of DnaK was induced with different concentrations of nisin (0, 0.05, 1.0, 10.0, 25.0 and 50 ng mL-1) for overnight. Nisin solution was prepared with commercial nisin of Streptococcus lactis (Sigma). Aliquots of the nisin-induced cultures were then transferred to fresh GM17 medium (30 mL) containing 5 μg mL-1 chloramphenicol and the same concentrations of nisin that was used for induction to obtain an OD600 of 0.04-0.05. Bacterial strains were then allowed to grow at 40°C and growths were measured at OD600. Bacterial strains that survived combined stresses of nisin and heat were detected using the same method. All the experiments were repeated at least three times to check the reproducibility of the results.
Confirmation of DnaK production by Western blotting: To check the production level of DnaK proteins after induction with different nisin concentration, overnight bacterial cultures (10 mL) were harvested by centrifuging at 6,000x g for 5 min at 4°C. Cell-free extracts were prepared from the pellets suspended in chilled 50 mM potassium phosphate buffer (pH 7.4) by using a Multi-Beads Shocker (Yasui Kikai, Osaka, Japan) at 2,500 rpm for 1 min at 4°C; this process was repeated 5 times with 1 min intervals. The cell-free extracts were obtained by centrifuging at 2,000x g for 15 min to remove the cell debris and the supernatant that contained the soluble proteins was collected. The protein concentration in the supernatant was determined by using the Bradford assay kit (Nacalai Tesque, Kyoto, Japan). The soluble proteins (10 μg) were then subjected to separation on a 12% (w/v) SDS-PAGE gel. The proteins were transferred onto a polyvinylidene difluoride membrane. Immunoblotting and detection of DnaK proteins were performed as previously described using anti-T. halophilus DnaK antibody (Sugimoto et al., 2008).
RESULTS
Overexpression of DnaK in L. lactis: To check the production of DnaK upon nisin induction, cell lysates of all four transformants induced with different concentrate of nisin (0-50 ng mL-1 nisin) was used for Western blot analysis (Fig. 1a-c). We could detect L. lactis DnaK (DnaKLla) in all the transformants including control strain (NZ-Vector). An additional band was detected in NZ-EDnaK only in presence of 10 ng mL-1 nisin which had an equal mobility of that of DnaK from E. coli cell extract used as a control (Fig. 1b). DnaKEco could be clearly distinguished from DnaKLla in NZ-EDnaK based on the difference in their mobilities. Between the two protein bands obtained from the cell lysate of NZ-EDnaK, the upper band was identified as DnaKEco. It was also observed that the amount of heterologously produced DnaKEco is very low compared to the NZ-LDnaK`s own DnaKLla. When NZ-LDnaK and NZ-TDnaK were induced with different nisin concentration overproduction of L. lactis DnaK (DnaKLla) and T. halophilus DnaK (DnaKTha) were detected in cell lysates of NZ-LDnaK and NZ-TDnaK, respectively (Fig. 1b).
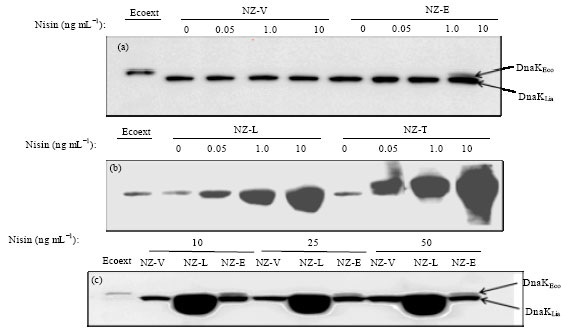 |
| Fig. 1(a-c): | DnaK production after nisn induction. Cell-free extracts of NZ-Vector (NZ-V), NZ-EDnaK (NZ-E), NZ-LDnaK (NZ-L) and NZ-TDnaK (NZ-T) prepared from cultures that were induced overnight with indicated concentration of nisin in GM17 liquid medium supplemented with 0.5% glucose at 30°C were separated using a 12% SDS gel for Western blotting. In cell free extracts production of DnaK was confirmed using DnaK polyclonal antibody raised against T. halophilus DnaK. (a) DnaK productions after induction with 0-10 ng mL-1 nisin were checked by western blot where a single blot was use for NZ-Vector and NZ-EDnaK. Here cell free extract of E. coli (Ecoext) was used as control for DnaKEco. (b) DnaKLla and DnaKTha production were checked in another blot in presence of 0-10 ng mL-1 nisin where same concentration of NZ-Vector cell free extract was used to compare the amount of these DnaKs. (c) DnaK productions in cell free extracts of NZ-Vector, NZ-EDnaK and NZ-LDnaK were compared in another blot after induction with 10-50 ng mL-1 nisin. Ecoext was used as control. Equal amounts of proteins were used in all lanes |
| Table 1: | Generation time and maximum growth yields of L. lactis strains in presence of different concentration of nisin GM17 media at 40°C |
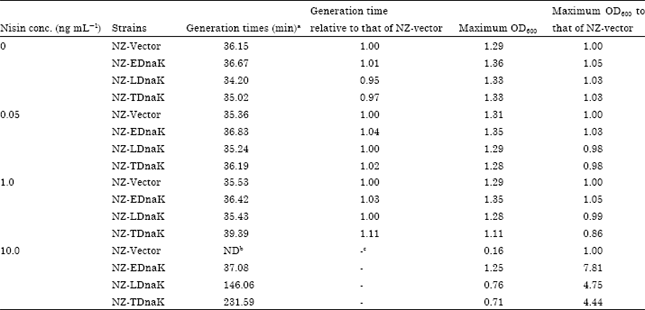 | |
aGeneration time was determined in the exponential growth phase. bND: Not detected (since the cells were not in exponential phase). c: Uncalculated | |
Although chromosomal- and plasmid-borne DnaK could not be separated in NZ-LDnaK and NZ-TDnaK the increased band intensity confirmed the overproduction of DnaKLla and DnaKTha in cell lysates of NZ-LDnaK and NZ-TDnaK, respectively. Figure 1 clearly shows that although 0.05 ng mL-1 nisin was quite enough to induce DnaK production in NZ-LDnaK and NZ-TDnaK (Fig. 1b) production of E. coli DnaK was induced only after induction with 10 ng mL-1 nisin (Fig. 1a). To check whether increased nisin induce further production of DnaK in the studied strains cell lysates of the strains after induction with 25 and 50 ng mL-1 nisin were compared with the cell lysates induced with 10 ng mL-1 nisin. It was then detected that nisin concentration more than 10 ng mL-1 did not induce more DnaK production in any of the strains of NZ-Vector, NZ-LDnaK, NZ-EDnaK (Fig. 1c) and NZ-TDnaK (Data not shown).
Tolerance to nisin stress by DnaK-overexpression at 40°C: While comparing the nisin stress tolerance of all heterologous or homologous DnaK-producing cells at 40°C, no growth difference of the strains in absence of nisin (Fig. 2a) and no visible growth of the strains in presence of 25 and 50 ng mL-1 nisin (Fig. 2e, f) clearly indicates a stress effect of this antimicrobial agent in presence of 40°C temperature. In presence of 0.05 ng mL-1 nisin, there were no significant difference in growth pattern (Fig. 2b), generation time and maximum OD600 (Table 1) although in this concentration of nisin NZ-LDnaK and NZ-TDnaK over-produced DnaKLla and DnaKTha (Fig. 1b). Again, in presence of 1 ng mL-1 nisin, the generation time and maximum OD600 of NZ-Vector, NZ-EDnaK and NZ-LDnaK were also not significantly different. However, DnaKTha producing cells showed a little higher generation time (1.11-fold) and lower maximum OD (0.86-fold) than the corresponding values of NZ-Vector (Table 1). A clear difference in stress tolerance by the studied strains was visible in presence of nisin concentration of 10 ng mL-1 only (Fig. 2c).
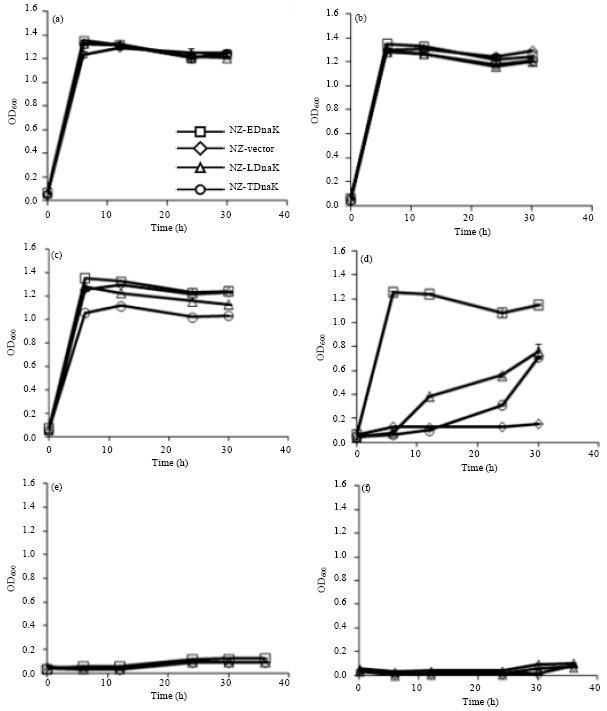 |
| Fig. 2(a-f): | Effect of nisin on growth at 40°C. NZ-Vector, NZ-EDnaK, NZ-LDnaK and NZ-TDnaK were grown in GM17 medium containing 0.5% glucose as the sole carbon source containing without pH control. Nisin concentrations of (a) 0 ng mL-1, (b) 0.05 ng mL-1, (c) 1 ng mL-1, (d) 10 ng mL-1, (e) 25 ng mL-1 and (f) 50 ng mL-1 were used in the media and the periodic growth was measured at 600 nm. The values shown are the Means± standard errors (error bars) for three independent experiments |
At 10 ng mL-1 nisin concentration, the concentration which is sufficient for the DnaK-overproduction in all the strains, NZ-EDnaK showed maximum growth yields among the strains indicating highest tolerance to the combined effect of nisin and heat. Since NZ-Vector was not in log phase generation time could not be compared. However, the maximum OD of NZ-EDnaK, NZ-LDnaK and NZ-TDnaK were 7.81, 4.75 and 4.44 fold higher than that of NZ-Vector (Table 1) suggesting the potency of overproduced DnaK to make the strains capable to grow even the conditions when NZ-Vector showed a very negligible growth.
DISCUSSION
Although recent developments in fermentation technology are capable of minimizing the stress conditions during fermentation, scientists are conducting research on stress-stable fermentation microorganisms and overexpression of molecular chaperone genes is now considered to be one of the most promising approaches to achieve that goal. We successfully overexpressed DnaKEco in L. lactis which showed multiple stress tolerance and higher lactic acid production at high temperature (Abdullah-Al-Mahin et al., 2010). However, the problem that was found is that nisin, the antimicrobial peptide used for DnaK overproduction, itself conferred stress to L. lactis strains (Abdullah-Al-Mahin et al., 2010). Antimicrobial agents were also reported to effect on the production of lactic acid and other fermentation products by LAB (Abou Ayana et al., 2011). Therefore, this study was aimed at evaluating the stress tolerance to nisin after expression of DnaK in lactic acid bacterium L. lactis NZ9000. Our study concluded with the finding that the expression of DnaK conferred increased tolerance to combined stress effect of heat and nisin to L. lactis NZ9000. The role of molecular chaperone in stress tolerance has already been reported by many researchers. Susin et al. (2006) reported the importance of DnaK/DnaJ for the survival of Caulobacter crescentus when exposed to heat stress. Previously, GroESL-overproducing L. lactis was reported to be more tolerant to heat (54°C for 30 min), salt (5 M NaCl for 1 h), or solvent (0.5% butanol) stress (Desmond et al., 2004). Sugimoto et al. (2003) earlier reported the remarkable suppression of the 5% (0.86 M) NaCl-induced protein aggregates by the overproduction of T. halophilus DnaK in E. coli. Tomas et al. (2003) reported that the overexpression of groESL in Clostridium acetobutylicum resulted in a 38 and 30% increase in acetone and butanol production, respectively, relative to the plasmid control strain during pH-controlled glucose-fed batch acetone-butanol fermentation. The groESL-overexpressing strain also showed increased tolerance against butanol than plasmid-controlled strain. Involvement of heat shock proteins to protect plants against heat and salt stresses has also been reported (Essemine et al., 2010; Mudgal et al., 2010; Joseph and Jini, 2010). These findings with regard to improved stress tolerance efficiency due to the expression of chaperone genes were consistent with the findings of present study. Further, the most important finding of this study was the tolerance of all the DnaK-overproducing strains to combined effects of heat and nisin.
Nisin is known to effect on cell membrane of gram-positive bacteria having the target site of lipid II (Guder et al., 2000; Sahl and Bierbaum, 1998). Increased sensitivity of heat stressed B. cereus and L. lactis to nisin was reported earlier (Beuchat et al., 1997; Kalchayanand et al., 1992, 1994). It was suggested that nisin prevented the repair of heat damaged membrane. In this study, combined action of heat and nisin (10 ng mL-1) effected on the cell growth of NZ-Vector. Homologously or heterologously overproduced DnaK, especially DnaKEco helped the strains to overcome the combined stresses.
Although DnaKLla and DnaKTha were produced higher than DnaKEco, more potentiality for combined stress tolerance was found by DnaKEco. These differences in DnaK overproduction ability can be explained by differences in codon usase (Abdullah-Al-Mahin et al., 2010). Our findings also suggested that DnaKEco was more efficient than DnaKLla and DnaKTha to overcome the stress at early growth period (Fig. 1d). Difference in stress tolerance ability by different DnaK has already been reported earlier (Abdullah-Al-Mahin et al., 2010; Sugimoto et al., 2003). However, despite higher DnaK production the failure to confer better stress tolerance by NZ-LDnaK and NZ-TDnaK suggested the importance of functional potency rather than the level of production.
Finally, it can be conclude that both homologously or heterologously overproduced DnaK had the effect to rescue the growth inhibition due to nisin at high temperature. This finding could have important implication for exploring the scope for further study to find the relationship of DnaK/chaperones and tolerance to different antimicrobial peptides. Since, the target site of nisin is cell membrane, the tolerance to nisin thereby opens a scope to research about the role of DnaK in membrane stability against bacteriocins.
REFERENCES
- Ahmed, R.G., 2006. The relation between biological consequences and temperature on some non-mammalian species. Int. J. Zool. Res., 2: 136-149.
CrossRefDirect Link - Ali, A.A., 2011. Isolation and identification of lactic acid bacteria isolated from traditional drinking yoghurt in Khartoum State, Sudan. Curr. Res. Bacteriol., 4: 16-22.
CrossRefDirect Link - Anukam, K.C., 2007. Probiotic toxicity, any evidence?. J. Pharmacol. Toxicol., 2: 590-598.
CrossRefDirect Link - Arnau, J., K.I. Sorensen, K.E. Appel, F.K. Vogensen and K. Hammer, 1996. Analysis of heat shock gene expression in Lactococcus lactis MG1363. Microbiology, 142: 1685-1691.
CrossRefDirect Link - Abou Ayana, I.A.A., A.A. Gamal El-Deen and M.A. El-Metwally, 2011. Behavior of certain lactic acid bacteria in the presence of pesticides residues. Int. J. Dairy Sci., 6: 44-57.
CrossRefDirect Link - Berns, K.I. and C.A. Thomas, 1965. Isolation of high molecular weight DNA from Haemophilus influenzae. J. Mol. Biol., 11: 476-490.
CrossRefDirect Link - Beuchat, L.R., M.R. Clavero and C.B. Jaquette. 1997. Effects of nisin and temperature on survival, growth, and enterotoxin production characteristics of psychrotrophic Bacillus cereus in beef gravy. Appl. Environ. Microbiol., 63: 1953-1958.
CrossRefDirect Link - Bukau, B., 1993. Regulation of the Escherichia coli heat-shock response. Mol. Microbiol., 9: 671-680.
CrossRefPubMedDirect Link - Bukau, B. and G.C. Walker, 1989. Cellular defects caused by deletion of the Escherichia coli dnaK gene indicate roles for heat shock protein in normal metabolism. J. Bacteriol., 171: 2337-2346.
CrossRefPubMedDirect Link - Bukau, B. and G. Walker, 1990. Mutations altering heat shock specific subunit of RNA polymerase suppress major cellular defects of E. coli mutants lacking the DnaK chaperone. EMBO J., 9: 4027-4036.
CrossRefDirect Link - Cegielska, A. and C. Georgopoulos, 1989. Functional domains of the Escherichia coli DnaK heat shock protein as revealed by mutational analysis. J. Biol. Chem., 264: 21122-21130.
CrossRefPubMedDirect Link - Craig, E.A. and C.A. Gross, 1991. Is hsp70 the cellular thermometer? Trends Biochem. Sci., 16: 135-140.
CrossRefPubMedDirect Link - de Ruyter, P.G., O.P. Kuipers and W.M. de Vos, 1996. Controlled gene expression systems for Lactococcus lactis with the food-grade inducer nisin. Appl. Environ. Microbiol., 62: 3662-3667.
CrossRefDirect Link - Desmond, C., R.P. Ross, E. O'Callagahan, G. Fitzegerald and C. Stanton, 2002. Improved survival of Lactobacillus paracasei NFBC 338 in spray dried powders containing gum acacia. J. Appl. Microbiol., 93: 1003-1011.
CrossRefPubMedDirect Link - Desmond, C., G.F. Fitzgerald, C. Stanton and R.P. Ross, 2004. Improved stress tolerance of GroESL-overproducing Lactococcus lactis and probiotic Lactobacillus paracasei NFBC338. Appl. Environ. Microbiol., 70: 5929-5936.
CrossRefDirect Link - Essemine, J., S. Ammar and S. Bouzid, 2010. Impact of heat stress on germination and growth in higher plants: Physiological, biochemical and molecular repercussions and mechanisms of defence. J. Biol. Sci., 10: 565-572.
CrossRefDirect Link - Fiocco, D., V. Capozzi, P. Goffin, P. Hols and G. Spano, 2007. Improved adaptation to heat, cold, and solvent tolerance in Lactobacillus plantarum. Appl. Microbiol. Biotechnol., 77: 909-915.
CrossRefDirect Link - Guder, A., I. Wiedemann and H.G. Sahl, 2000. Posttranslationally modified bacteriocins-the lantibiotics. Biopolymers, 55: 62-73.
CrossRefPubMedDirect Link - Gursoy, O. and O. Kinik, 2006. Probiotics: A new popular option for cancer inhibition. Int. J. Dairy Sci., 1: 100-103.
CrossRefDirect Link - Holo, H. and I.F. Nes, 1989. High-frequency transformation, by electroporation, of Lactococcus lactis subsp. cremoris grown with glycine in osmotically stabilized media. Appl. Environ. Microbiol., 55: 3119-3123.
CrossRefDirect Link - Homuth, G., S. Masuda, A. Mogk, Y. Kobayashi and W. Schumann, 1997. The dnaK operon of Bacillus subtilis is heptacistronic. J. Bacteriol., 179: 1153-1164.
CrossRefDirect Link - Joseph, B. and D. Jini, 2010. Proteomic analysis of salinity stress-responsive proteins in plants. Asian J. Plant Sci., 9: 307-313.
CrossRefDirect Link - Kalchayanand, N., T. Sikes, C.P. Dunne and B. Ray, 1994. Hydrostatic pressure and electroporation have increased bactericidal efficiency in combination with bacteriocins. Appl. Environ. Microbiol., 60: 4174-4177.
CrossRefDirect Link - Kilstrup, M., S. Jacobsen, K. Hammer and F.K. Vogensen, 1997. Induction of heat shock proteins DnaK, GroEL, and GroES by salt stress in Lactococcus lactis. Appl. Environ. Microbiol., 63: 1826-1837.
CrossRefDirect Link - Kullen, M.J. and T.R., Klaenhammer, 1999. Identification of the pH-inducible, proton-translocating F1Fo-ATPase (atpBEFHAGDC) operon of Lactobacillus acidophilus by differential display: Gene structure, cloning and characterization. Mol. Microbiol., 33: 1152-1161.
CrossRefPubMedDirect Link - Lee, B.B., H.J. Tham and E.S. Chan, 2007. Fed-batch fermentation of lactic acid bacteria to improve biomass production: A theoretical approach. J. Applied Sci., 7: 2211-2215.
CrossRefDirect Link - Lentze, N. and F. Narberhaus, 2005. Small heat shock proteins OR: A subgroup of molecular chaperones. J. Biol. Sci., 5: 1-9.
CrossRefDirect Link - Liberek, K., J. Marszalek, D. Ang, C. Georgopoulos and M. Zylicz, 1991. Escherichia coli DnaJ and GrpE heat shock proteins jointly stimulate ATPase activity of DnaK. Proc. Natl. Acad. Sci. USA., 88: 2874-2878.
CrossRefPubMedDirect Link - Abdullah-Al-Mahin, S. Sugimoto, C. Higashi, S. Matsumoto and K. Sonomoto, 2010. Improvement of multiple-stress tolerance and lactic acid production in Lactococcus lactis NZ9000 under conditions of thermal stress by heterologous expression of Escherichia coli dnaK. Applied Environ. Microbiol., 76: 4277-4285.
CrossRefPubMedDirect Link - Marmur, J., 1961. A procedure for the isolation of deoxyribonucleic acid from micro-organisms. J. Mol. Biol., 3: 208-218.
CrossRefDirect Link - Mayer, M.P., D. Brehmer, C.S. Gassler and B. Bukau, 2001. Hsp70 chaperone machines. Adv. Protein. Chem., 59: 1-44.
CrossRefPubMedDirect Link - Mudgal, V., N. Madaan and A. Mudgal, 2010. Biochemical mechanisms of salt tolerance in plants: A review. Int. J. Bot., 6: 136-143.
CrossRefDirect Link - Prasad, J., P. McJarrow and P. Gopal, 2003. Heat and osmotic stress responses of probiotic Lactobacillus rhamnosus HN001 (DR20) in relation to viability after drying. Appl. Environ. Microbiol., 69: 917-925.
CrossRefDirect Link - Sahl, H.G. and G. Bierbaum, 1998. Lantibiotics: Biosynthesis and biological activities of uniquely modified peptides from Gram-positive bacteria. Ann. Rev. Microbiol., 52: 41-79.
PubMedDirect Link - Schulz, A., B. Tzschaschel and W. Schumann, 1995. Isolation and analysis of mutants of the dnaK operon of Bacillus subtilis. Mol. Microbiol., 15: 421-429.
CrossRefDirect Link - Sugimoto, S., J. Nakayama, D. Fukuda, S. Sonezaki, M. Watanabe, A. Tosukhowong and K. Sonomoto, 2003. Effect of heterologous expression of molecular chaperone DnaK from Tetragenococcus halophilus on salinity adaptation of Escherichia coli. J. Biosci. Bioeng., 96: 129-133.
CrossRefPubMedDirect Link - Sugimoto, S., K. Saruwatari, C. Higashi, K. Tsuruno, S. Matsumoto, J. Nakayama and K. Sonomoto, 2008. In vivo and in vitro complementation study comparing the function of DnaK chaperone systems from halophilic lactic acid bacterium Tetragenococcus halophilus and Escherichia coli. Biosci. Biotechnol. Biochem., 72: 811-822.
CrossRefDirect Link - Susin, M.F., R.L. Baldini, F. Gueiros and S.L. Gomes, 2006. GroES/GroEL and DnaK/DnaJ have distinct roles in stress responses and during cell cycle progression in Caulobacter crescentus. J. Bacteriol., 188: 8044-8053.
CrossRefDirect Link - Tomas, C.A., N.E. Welker and E.T. Papotasakis, 2003. Overexpression of groESL in Clostridium acetobutylicum resulted in increased solvent production and tolerance, prolonged metabolism, and changes in cell's transcriptional program. Appl. Environ. Microbiol., 69: 4951-4965.
CrossRefDirect Link - Van de Guchte, M., P. Serror, C. Chervaux, T. Smokvina, S.D. Ehrlich and E. Maguin, 2002. Stress responses in lactic acid bacteria. Antonie Leeuwenhoek, 82: 187-216.
CrossRefPubMedDirect Link - Walker, D.C., H.S. Girgis and T.R. Klaenhammer, 1999. The groESL chaperone operon of Lactobacillus johnsonii. Appl. Environ. Microbiol., 65: 3033-3041.
CrossRefDirect Link - Walter, J., N.C.K. Heng, W.P. Hammes, D.M. Loach, G.W. Tannock and C. Hertel, 2003. Identification of Lactobacillus reuteri genes specifically induced in the mouse gastrointestinal tract. Appl. Environ. Microbiol., 69: 2044-2051.
CrossRefDirect Link - Whitaker, R.D. and C.A. Batt, 1991. Characterization of the heat shock response in Lactococcus lactis subsp. lactis. Appl. Environ. Microbiol., 57: 1408-1412.
CrossRefDirect Link - Yateem, A., M.T. Balba, T. Al-Surrayai, B. Al-Mutairi and R. Al-Daher, 2008. Isolation of lactic acid bacteria with probiotic potential from camel milk. Int. J. Dairy Sci., 3: 194-199.
CrossRefDirect Link - Kalchayanand, N.R., M.B. Hanilin and B. Ray, 1992. Sublethal injury makes Gram-negative and resistant Gram-positive bacteria sensitive to the bacteriocins, pediocin AcH and nisin. Lett. Applied Microbiol., 15: 239-243.
CrossRef