
Hany K. Abd-Elhady
Department of Pesticides, Faculty of Agriculture, Menoufiya University, Egypt
LiveDNA: 20.1484
Ahmed A. Abd El-Aal
Department of Economic Entomology and Agricultural Zoology, Faculty of Agriculture, Menoufiya University, Egypt
American Journal of Biochemistry and Molecular Biology
Year: 2011 | Volume: 1 | Issue: 4 | Page No.: 337-348







ABSTRACT
In Egypt pink bollworm Pectinophora gossypiella (Lepidoptera: Gelechiidae) has recently developed resistance to various kinds of insecticide in the field. The resistance status of pink bollworm to several classes of insecticides currently used in cotton fields was evaluated in the Nile Delta (Kafr-Elsheikh and Menoufia) Egypt. Two field-collected P. gossypiella populations have developed resistance to several classes of insecticides. Toxicological bioassays were conducted in the laboratory, LC50’s estimated by probit analysis and Resistance Ratio (RR) were calculated at the LC50 level. Of the seven insecticides tested four insecticides (chlorpyrifos, profenofos, thiodicarb and carbaryl) showed the Resistance Ratio (RR) values above 20 folds, against two field-collected populations whereas the other three insecticides (fenpropathrin, spinosad and abamectin) had RR values less than 10 folds. Also, the patterns of selection pressure by abamectin at the LC30 level to Kafr-Elsheikh (KFR)-and Menoufia (MNF)-field strains were determined by continuous insecticide application under laboratory conditions for eight successive generations. On the other hand, biochemical assays indicated that both KFR-and MNF-field strains expressed higher levels of Acetylcholinesterase (AChE) activity by 4.1 and 2.8-times, respectively, compared to that of susceptible strain. There were significant differences in esterase activities among the field-collected populations and susceptible strain. Analysis of protein patterns by (SDS-PAGE) showed some differences between resistant strains to abamectin, MNF-and KFR-field strains and the bands differed in intensity and molecular weights between all strains. These findings indicate that rapid biochemical assays coupled with biological assays are potential tools for estimating both the intensity of resistance in the field.
PDF Abstract XML References Citation
Received: February 28, 2011;
Accepted: May 28, 2011;
Published: June 25, 2011
How to cite this article
Hany K. Abd-Elhady and Ahmed A. Abd El-Aal, 2011. Insecticides Resistance Detection in Field-collected Populations of Pectinophora gossypiella (Saunders). American Journal of Biochemistry and Molecular Biology, 1: 337-348.
DOI: 10.3923/ajbmb.2011.337.348
URL: https://scialert.net/abstract/?doi=ajbmb.2011.337.348
DOI: 10.3923/ajbmb.2011.337.348
URL: https://scialert.net/abstract/?doi=ajbmb.2011.337.348
INTRODUCTION
Cotton is attacked by various pests during the different stages of its development (Abd El-Mageed et al., 2007). The pink bollworm Pectinophora gossypiella (Saunders), is the key pest in cotton fields, particularly in middle and late season under dry conditions (Korejo et al., 2000; Unlu, 2004). At present, it has been recorded in nearly all cotton-growing countries of the world. Newly hatched larvae are exposed to insecticides for a very short time before they enter flower buds or bolls while fully grown larvae emerging from bolls for pupation are difficult to control with chemicals (Attique et al., 2001a, b). Traditionally, chemical control is the major method for protecting cotton from pink bollworm damage but the efficacy of these insecticides has declined quickly due to the evolution of resistance to insecticides in pink bollworm (Yang et al., 2000). Susceptibility to pesticides in target agricultural pest populations is a valuable natural resource in agriculture that must be managed carefully sustains efficacy of pesticides as long as possible, especially if the pesticide is environmentally friendly. Bioassays have been the basic used for documenting levels of insecticides resistance in field populations on which management decisions are based (Shah et al., 2002). Given the history of resistance to various insecticides of insect pests of cotton, a need continues to develop baseline data, to monitor for changes in tolerance, is essential for detecting shifts in susceptibility (Margaritopoulos et al., 2008). Therefore, resistance monitoring is fundamentally important to every resistance management program. Continuous monitoring of the populations for changes in resistance frequencies allows informed decision making decisions regarding effective management strategies (Prabhaker et al., 1996). Provided that a monitoring survey is sufficiently extensive, patterns of resistance can be detected within a region or between regions. Ideally, current resistance monitoring data should be communicated to growers and pest control advisors who are interested in participating in a resistance management program. With this information, they could alternate to a different insecticide class or alternative control strategy to avoid increasing selection pressure to populations (Prabhaker et al., 1996). Biochemical techniques are advantageous because they test for activity that is directly linked to a resistance mechanism (Byrne et al., 2000). On the other hand, biochemical assays may also be used to detect and monitor resistance. Biochemical assays measure changes in sensitivity at the pesticide's target site or measure metabolic activities of the pest that are responsible for, or at least linked to, resistance (Roditakis et al., 2009). Chemical insecticides are known to be able to act as inducers for the biosynthesis of cellular proteins or as inhibitors of enzymes; thus, the molecular biochemical explanation of variations among the tested field strains can be attributed to either variation or similarity in the background exposure to the chemical control programs. However, early resistance detection is very difficult using conventional bioassay-based monitoring methods, particularly when resistance is recessive (Clark, 2010). In addition, it is difficult collecting large numbers of some insects. A biochemical assay used by Soleno et al. (2008) showed different degrees of tolerance associated with increased esterase activities in codling moth populations. Bioassays on adult moths are successfully utilised for monitoring resistance in codling moth (Dunley and Welter, 2000). More recently, it has been demonstrated that larvae can also be used for detection of resistance in field collections (Soleno et al., 2008).
The objectives of this study were (1) to further describe the expression of resistance to different insecticides of field populations of the pink bollworm collected from cotton fields in the Nile Delata (2) to prolong the useful life of abamectin we selected field strains of the pink bollworm. We herein present the results of our characterization of resistance level of the selected strains and possible mechanisms of resistance and (3) to investigate the activities of esterases, acetylcholinesterase and analysis of protein patterns by (SDS-PAGE) in both resistant and susceptible strains.
MATERIALS AND METHODS
Strain collection and rearing procedures: A susceptible strain of pink bollworm was obtained from the Lab. of Bollworms Department at the Institute of Plant Protection, Dokki, Egypt and maintained in the laboratory under insecticide-free conditions. It was used as the baseline reference strain for the comparisons levels of resistance between the studied strains. Two P. gossypiella field strains were collected from different fields of cotton located in Menoufia (MNF) and Kafr-Elsheikh (KFR), Nile Delta area in season 2009. Pink bollworm larvae were reared on a modified artificial diet as described by El-Hafez et al. (1982). All bioassay trials were done on the newly hatched larvae of the pink bollworm.
Chemicals and insecticide bioassays: The commercial formulations of insecticides used in bioassays were: chlorpyrifos (Dursban 48% Emulsifiable Concentrate (EC), DowElanco Co.); profenofos (Curacron 72% EC, Ciba-Geigy AG); thiodicarb (Larvin 80% DF [dust flowable], Union Carbide Agricultural Co., Inc.); carbaryl (Sevin 85% WP [wettable powder], Rhone-Poulenc); fenpropathrin (Danitol 30% EC, Sumitomo Chemical Co. Ltd; spinosad (Success 12% SC [suspension concentrate], Dow AgroSciences); and abamectin (Vertemic 1.8% EC, Syngenta, Basel, Switzerland). All other chemicals used in biochemical assays were purchased from Sigma Chemical Company as analytical standards.
Contact bioassay: A stock solution of each insecticide formulation was prepared using the acetone and then serially diluted concentrations were prepared. One ml of each insecticide concentration was pipetted into a Petri dish (7.5 cm diameter). After accurately distribution of solutions, dishes were left until dryness then ten newly hatched larvae (12 to 24 h-old) were transferred to each dish containing control diet and held at the same conditions (27±1°C and 70-75% RH). Control Petri dishes were precoated with acetone only. Mortality was assessed 48 h after treatment. Control mortality was never >10%; data were corrected by Abbott (1925) formula. The data were analyzed by probit analysis PC (Finney, 1971). Resistance Ratio (RR) was calculated by dividing the LC50 of field-collected strain by the LC50 of lab-susceptible strain.
Establishment of the resistant strain to abamectin: For selecting the resistant strain of pink bollworm P. gossypiella, abamectin (Vertemic 1.8% EC) was dissolved in acetone and applied using the contact bioassay technique. One mL of the abamectin concentration which kills 30% of first-instar larvae LC30 (after 48 h) was pipetted into a Petri dish. The surviving larvae were transferred into glass vials containing artificial diet. Toxicity tests were conducted every two generations to calculate its LC50.
Biochemical assays
Acetylcholinesterase (AChE) assay: AChE assays were measured using pink bollworm adult head homogenates following the method of Ellman et al. (1961). Data were corrected using a blank test without substrate in the mixture reaction. Protein content was determined using methods described by Bradford (1976). The AChE activity was determined using an extinction coefficient of 13.6 mM-1 cm-1 for thionitrobenzoic acid.
Esterase assays: Esterase activity towards α-NA was quantified using the assay method of Van Asperen (1962). An esterase assay was conducted to measure the production of α-naphthol from α-naphthyl acetate substrate. Esterase activity was presented relative to protein concentration which was measured using the assay of Bradford (1976). The esterase activity was expressed as μmol α-naphthol produced/min/mg protein.
Electrophoretic analysis (SDS-PAGE) of protein: SDS-PAGE was carried out according to methods described by Laemmli (1970). The intensities of the bands on each gel were then scanned with a computing densitometer at absorbance of 6J3 nm and results were graphically depicted with Image Phoretic (IDA) Quantifier software (Phoretix International, London). The peak heights, Rf, intensity and area were used as a relative comparison of band intensity between zygomorphs on the same gel.
Statistical analysis: The data was analyzed using analysis of variance (ANOVA) followed by the Student-Newman Keuls test to determine significance between different groups. These tests were performed using a computer software CoStat system for Windows, CoStat Program, 2006 Berkeley, CA, USA.
RESULTS AND DISCUSSION
Insecticides resistance in field populations: Present studies were carried out to evaluate the toxicity of some selected insecticides bearing novel modes of action against first instars of pink bollworm from the Lab-susceptible and field strains collected from cotton fields.
High level of resistance was detected for both field-collected strains to all of the organophosphate and carbamate insecticides tested (Table 1). Among the field-collected strains, KFR was more resistant to the organophosphates and carbamates than the MNF strain. Tests with pyrethroids revealed low resistance levels to fenpropathrin in both field-collected strains (Table 1).
| Table 1: | Responses of 1st instar of pink bollworm larvae from field strains to selected insecticides as assayed by contact technique bioassay |
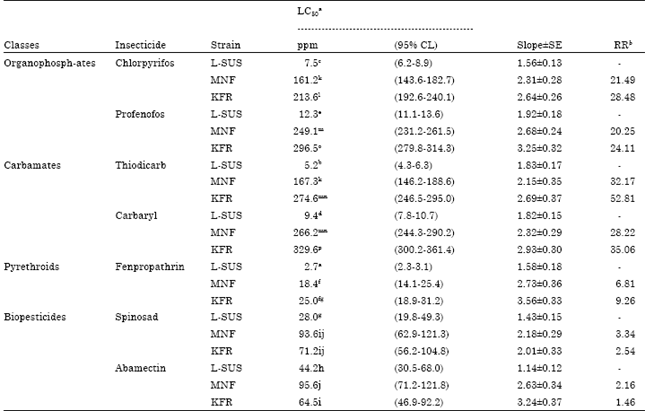 | |
aExpressed as ppm after 48 h of treatment and LC50 values followed by the same letter within each vertical column are not significantly different (95% CL do not overlap). bResistance ratio (RR) calculated by dividing the LC50 for the field strain by the LC50 for the Lab-susceptible strain | |
The field-collected strains did not differ from each other in susceptibility to fenpropathrin. Resistance to the biopesticides, spinosad and abamectin was low or insignificant in the case of KFR’s response to abamectin. The field strains showed less than 2.2-fold resistance to abamectin but exhibited variable levels (6.81 to 52.81-fold) of resistance to other covenantal insecticides used. Both field strains use in this study appear to not be homogenous for resistance to the insecticides we tested. Resistance of pink bollworms to biopesticides seems to be correlated with the pattern of insecticide-use prevalent in the localities studied. Therefore, our study is in agreement with the findings of Osman et al. (1991) who suggested that monitoring of pink bollworm resistance in field strains reared in the laboratory should be performed in the FI generation. The spinosad, abamectin and fenpropathrin resistance were generally low. It has been suggested that insects should not be considered resistant until a resistance ratio of 10 is exhibited (Torres-Vila et al., 2002; Kranthi et al., 2002; Bues et al., 2005). Current levels of resistance to selected insecticides, chlorpyrifos, profenofos, thiodicarb and carbaryl, in two field locations were high which is probably due to intensive use of these insecticides that results in high selection pressure for the development of resistance. These developments have had a dramatic effect on management plans for pink bollworm P. gossypiella control. For example, management strategies for pyrethroid resistance have relied on organophosphates and carbamates to reduce the frequency of pyrethroid use during recent years (Plapp et al., 1990), so exposure of insect to these compounds has increased. The cotton insect management plan recommended by Baldwin and Graves (1991) for Louisiana seems to be reliable to apply in Egypt. In essence, the window strategy, based on the use of a single class of insecticide on discrete generations of an insect pest is still advocated. However, the pyrethroids are recommended from 1 July to 15 August and organophosphates are then recommended during the late season. Based on our current study we are in agreement with recommendations of Baldwin and Graves (1991) which encourage the use of biological insecticides, especially during the early season. The susceptibility of pests to insecticides may change depending on the selection pressure of these compounds on population (Sayyed et al., 2005).
Selection of the abamectin-resistant strain from collected field populations: Resistance of the KFR-strain at F2 was estimated to be about 3.85-fold compared with the susceptible-strain (LC50 = 170.2) (Table 2).
Resistance in the selected KFR strain increased steadily from 3.85-fold at F2 to 21.01-fold at F8 (Table 2). The development of resistance in the MNF strain was minor until F4 after which it increased sharply to 15.90-fold at F8. Exponential increase of resistance was found from the F8. After 8 generations of selection, 48 h-LC50 value of abamectin against the 1st instar larvae increased significantly which was from 95.6 to 702.6 ppm. After 8 generations of selection the resistance level to abamectin in the MNF-selected strain increased more than 15-fold compared with the parental strain. Clarification of the resistant mechanism of P. gossypiella to abamectin is important to Integrated Pest Management (IPM) and Insect Resistance Management (IRM). The results showed that the level of resistance increased more than 20-fold compared with the parental strain after 8 generations of selection. Although biopesticides are effective on P. gossypiella larvae, its resistance has been documented in other bollworms of some areas (Gunning and Balfe, 2002; Wyss et al., 2003; Ahmad et al., 2003). In Pakistan, Spodoptera litura were investigated for spinosad, resistance ratio was in the range of 7 to 122-fold and 4 to 186-fold for abamectin (Ahmad et al., 2008). The selection pressure is responsible for the difference in resistance among regions is the positive correlation found between the number of insecticidal applications and the degree of resistance development (Kakani et al., 2010).
| Table 2: | Rate of development of resistance at 48 hr in first-instar of pink bollworm larvae field strains (Menoufiya and Kafr-Elsheikh) after exposure to selection at LC30 level with abamectin for eight successive generations |
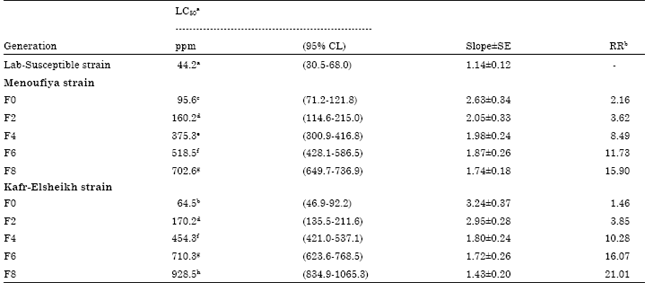 | |
aExpressed as ppm after 48 h of treatment and LC50 values followed by the same letter within each vertical column are not significantly different (95% CL do not overlap). bResistance ratio (RR) calculated by dividing the LC50 for the field strain by the LC50 for the Lab-susceptible strain | |
AchE and esterase activities: Insects from field populations collected either from MNF (F0, parents) or KFR (F0, parents) cotton fields, expressed higher levels of AChE activity than the reference (Lab-susceptible) strain, in which AChE activity was 2.80 and 4.10-fold higher than that from the Lab-susceptible strain, respectively (Table 3).
However, AChE activity increased following laboratory rearing under selection pressure of both MNF and KFR indicating that these field-collected strains appeared to increase their resistance to abamectin. The activity in F8 generation of MNF strain was similar to that in the KFR (F0, parents) strain whereas insects from MNF-F8 generation showed an increase in AChE activity by 3.90-fold higher than that in Lab-susceptible strain. Therefore, present data demonstrated that increase in resistance to abamectin in both field-collected strains seems to be associated with AChE activity. This also may be particularly important for understanding AChE insensitivity is an important insecticide resistance mechanism in a number of insect species (Soderlund and Blopmquist, 1990; Harold and Ottea, 1997). Esterase activities toward α-naphthyl acetate (α-NA) in pink bollworm larvae from both MNF-and KFR field strain were higher than those measured in Lab-susceptible strain (Table 3). Similarly, esterase activities in F0 of both field-collected strains were slightly elevated, 1.31 and 1.51-fold for the MNF and KFR strains, respectively, relative to the Lab-Sus strain. Additionally, esterase activity increased to 1.61 and 2.26-fold for these respective strains. Metabolic resistance to abamectin has also been implicated for the American bollworm, Helicoverpa armigera (Christie and Wright, 1990).
Protein levels: Differences in protein level among the various strains were observed in last instars (Table 4). The abamectin resistance selected strains had the highest protein levels followed by the field-collected strains. Similarly, protein level in F0 of both field-collected strains were slightly elevated, 1.07 and 1.10-fold for the MNF and KFR (F0) strains, respectively, relative to the Lab-Sus strain.
| Table 3: | Levels of acetylcholinesterase (AChE) and esterase activities in lab-susceptible strain, field-strains (F0) and F8-abamectin resistant strains of pink bollworm |
 | |
aAChE activity was measured in heads of pink bollworm moths toward acetylcholine iodide and expressed as nmol/min/mg protein. bEsterases activity were measured in midgut homogenate from pink bollworm larvae toward α-naphthyl acetate (α-NA) and expressed as nmol α-naphthol/min/mg protein. Values by the same letter within each vertical column are not significantly different (p = 0.05) | |
| Table 4: | Mean of protein levels in lab-susceptible strain, field-strains (F0) and F8-abamictin resistant strains of pink bollworm |
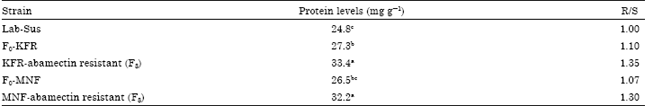 | |
Values by the same letter within each vertical column are not significantly different (Turkey's p = 0.05) | |
| Table 5: | Molecular weight and relative mobility (Rf) values of protein bands in the body tissues of pink bollworm larvae |
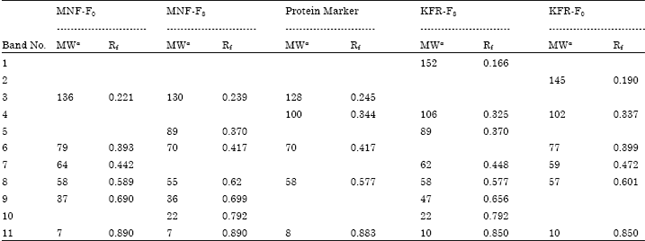 | |
aMolecular weight in kilodaltons. The four protein profile patterens (1,2,4 and 5) which presented by field-strains (F0) and F8-abamectin resistant strains of P. gossypiella | |
However, the F0-MNF strain did not significantly differ from the Lab-Sus strain. Additionally, protein level increased significantly to 1.30 and 1.35-fold for the MNF and KFR abamectin resistant (F8) strains, respectively.
Protein electrophoresis: Gel scanning with respective standard molecular weights revealed variation in protein bands number and molecular weight type among the four strains (Table 5). There were two new bands in the KFR-, MNF-abamectin resistant strains (F8) which had molecular weight (89 and 22), compared with both field-collected strains (F0).
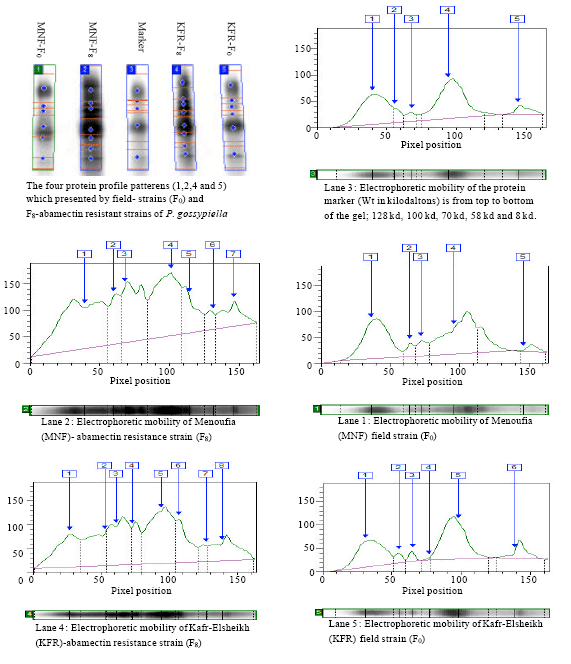 |
| Fig. 1: | Electrophoretic analysis of total proteins patterns by (SDS-PAGE) extracted from the last-larval instar of P. gossypiella belong to four different strains: Lane 1: Menoufia (MNF) field strain (F0), Lane 2: Menoufia (MNF) abamectin resistance strain (F8) Lane 3: Protein Marker, Lane 4: Kafr-Elsheikh (KFR),abamectin resistance strain (F8) Lane 5: Kafr-Elsheikh (KFR) field strain (F0) |
These specific protein bands were absent in field-collected strains, which may indicate a resistance mechanism present in these strains and that the SDS-PAGE technique may be used to screen and detect abamectin resistance. A rapid detection tool such as this may be valuable for determining in-season insecticide selection for resistance management Fig. 1.
Similarly, Abdeen (1985) found that exposure of cotton bollworm, Helicoverpa zea, to diflubenzuron resulted in an increase of some major proteins through the first two days post treatment. Also, Hewady (1990) indicated that when the haemolymph protein of normal 5th instar spiny bollworm Earias insulana was separated on acrylamide gel electrophoresis, 12 different proteins were recorded. Their mobility resolution (M.R.) values were 6.49 to 98.7. Twenty days after pyriproxyfen treatments caused appearance or disappearance of some bands with different mobility resolution. Jongsma et al. (1996) characterized the proteolytic activity present in the luminal contents of larval gut of Spodoptera exigua. Separation of gut proteins by SDS-PAGE and subsequent incubation with proteinase substrates visualized 6 separate activities with apparent molecular masses that ranged from 29 to 110 Kda. Eid et al. (1979) reported that no protein bands related to insecticide poisoning were observed in the larval tissue of S. littoralis, until 18 h post insecticide treatment when two different bands were detected in haemolymph protein as a result of profenofos and fenitrothion treatments.
CONCLUSION
Early detection of insecticide resistance provides a basis for the management of resistant pest populations (Osorio et al., 2008). It seems that little by little, the industrialized countries adapt new procedures for control of agricultural pests (Mirmoayedi et al., 2010). Rapid biochemical assays coupled with biological assays are potential tools for estimating both the intensity and the frequency of resistance in the field (Rauch and Nauen, 2003; Farghaly, 2010). For example, measurement of AChE sensitivity to inhibition by insecticides or resistance associated esterases with naphthyl acetate esters provides a rapid indication of resistance mechanisms expressed in insecticide-resistant pests (Brown et al., 1996). Additionally, the SDS-PAGE technique can be a biomarker for detection of susceptibility levels or resistance in the field strains which should lead to a change in the currently applied chemicals (Massoud et al., 2006).
REFERENCES
- Abbott, W.S., 1925. A method of computing the effectiveness of an insecticide. J. Econ. Entomol., 18: 265-267.
CrossRefDirect Link - Ahmad, M., A.H. Sayyed, M.A. Saleem and M. Ahmad, 2008. Evidence for field evolved resistance to newer insecticides in Spodoptera litura (Lepidoptera: Noctuidae) from Pakistan. Crop Prot., 27: 1367-1372.
CrossRefDirect Link - Ahmad, M., M.I. Arif and Z. Ahmad, 2003. Susceptibility of Helicoverpa armigera (Lepidoptera: Noctuidae) to new chemistries in Pakistan. Crop Prot., 22: 539-544.
CrossRefDirect Link - Attique, M.R., M.M. Ahmad, Z. Ahmad and M. Rafiq, 2001. Sources of carry-over and possibilities of cultural control of Pectinophora gossypiella (Saunders) in the Punjab, Pakistan. Crop Prot., 20: 421-426.
CrossRefDirect Link - Attique, M.R., Z. Ahmad, A.I. Mohyuddin and M.M. Ahmad, 2001. Studies on Pectinophora gossypiella (Saunders) and its control strategy in the Punjab, Pakistan. Pak. J. Zool., 33: 115-123.
Direct Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Brown, T.M., P.K. Bryson, F. Arnette, M. Roof, I.L. Mallett, J.B. Graves and S.R. Nemec, 1996. Surveillance of Resistant Acetylcholinesterase in Heliothis virescens. In: Molecular Genetics and Evolution of Pesticide Resistance, Brown, T.M. (Ed.). American Chemical Society Books, Washington, D.C., ISBN-13: 9780841234536, pp: 149-157.
- Bues, R., J.C. Bouvier and L. Boudinhon, 2005. Insecticide resistance and mechanisms of resistance to selected strains of Helicoverpa armigera (Lepidoptera: Noctuidae) in the South of France. Crop Prot., 24: 814-820.
CrossRefDirect Link - Byrne, F.J., K.L. Gorman, M. Cahill, I. Denholm and A.L. Devonshire, 2000. The role of B-type esterases in conferring insecticide resistance in the tobacco whitefly, Bemisia tabaci (Genn). Pest. Manag. Sci., 56: 867-874.
CrossRefDirect Link - Christie, P.T. and D.J. Wright, 1990. Activity of abamectin against larval stages of Spodoptera littoralis boisduval and Heliothis armigera hubner (Lepidoptera: Noctuidae) and possible mechanisms determining differential toxicity. Pestic. Sci., 29: 29-38.
CrossRefDirect Link - Clark, J. M., 2010. Permethrin resistance due to knockdown gene mutations is prevalent in human head louse populations. J. Open Dermatol., 4: 63-68.
CrossRefDirect Link - Dunley, J.E. and S.C. Welter, 2000. Correlated insecticide cross-resistance in azinphosmethyl resistant codling moth (Lepidoptera: Tortricidae). J. Econ. Entomol., 93: 955-962.
CrossRefPubMedDirect Link - Ellman, G.L., K.D. Courtney, V. Andres Jr. and R.M. Featherstone, 1961. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol., 7: 88-90.
CrossRefDirect Link - Abd El-Mageed, A.E.M., E.M. Anwar, L.R.A. Elgohary and H.F. Dahi, 2007. Evaluation of several programs of sequences pesticides application on cotton bollworms and some other sucking pests in cotton field. J. Entomol., 4: 93-103.
CrossRefDirect Link - Finney, D.J., 1971. Probit Analysis. 3rd Edn., Cambridge University Press, London, UK., pp: 76-80.
CrossRefDirect Link - Harold, J.A. and J.A. Ottea, 1997. Toxicological significance of enzyme activities in profenofos-resistant tobacco budworms, Heliothis virescens (F.). Pestic. Biochem. Physiol., 58: 23-33.
CrossRefDirect Link - Jongsma, M.A., J. Peters, W.J. Stiekema and D. Bosh, 1996. Characterization and partial purification of gut proteinases of Spodoptera exigua Hubner (Lepidoptera: Noctuidae). Insect Biochem. Mol. Biol., 26: 185-193.
CrossRefDirect Link - Kakani, E.G., N.E. Zygouridis, K.T. Tsoumani, N. Seraphides, F.G. Zalom and K.D. Mathiopoulos, 2010. Spinosad resistance development in wild olive fruit fly Bactrocera oleae (Diptera: Tephritidae) populations in California. Pest Manage. Sci., 66: 447-453.
CrossRefPubMedDirect Link - Korejo, A.K., A.W. Soomro, G.H. Mallah, A.A. Memon and M.Y. Solangi, 2000. Monitoring pink bollworm seasonal population through the use of gossyplure. Pak. J. Biol. Sci., 3: 2119-2121.
CrossRefDirect Link - Kranthi, K.R., D.R. Jadhav, S. Kranthi, R.R. Wanjari, S.S. Ali and D.A. Russell, 2002. Insecticide resistance in five major insect pests of cotton in India. Crop Prot., 21: 449-460.
CrossRefDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Margaritopoulos, J.T., G. Skavdis, N. Kalogiannis, D. Nikou and E. Morou et al., 2008. Efficacy of the pyrethroid alpha-cypermethrin against Bactrocera oleae populations from Greece and improved diagnostic for an iAChE mutation. Pest Manage. Sci., 64: 900-908.
CrossRefDirect Link - Massoud, M.A., A.K. Mourad, B.A. El and S.M. El, 2006. Biochemical studies on different field strains of the pink bollworm Pectinophora gossypiella (Saund.) (Lepidoptera: Gelechiidae) in Egypt. Commun. Agric. Applied Biol. Sci., 71: 517-535.
PubMedDirect Link - Mirmoayedi, A., M. Maniee and A. Yaghutipoor, 2010. Control of cotton spiny bollworm, Earias insulana boisduval, using three Bio-Insecticides, Bt, Spinosad and Neem-Azal. J. Entomol., 7: 89-94.
CrossRefDirect Link - Osman, A.A., T.F. Watson and S. Sivasupramanium, 1991. Reversion of permethrin resistance in field strains and selection for azinphosmethyl and permethrin resistance in pink bollworm (Lepidoptera: Gelechiidae). J. Econ. Entomol., 84: 353-357.
CrossRefDirect Link - Osorio, A., A.M. Martınez, M.I. Schneider, O. Diaz and J.L. Corrales et al., 2008. Monitoring of beet armyworm resistance to spinosad and methoxyfenozide in Mexico. Pest Manage. Sci., 64: 1001-1007.
CrossRefPubMedDirect Link - Prabhaker, N., N.C. Toscano, T.J. Henneberry, S.J. Castle and D. Weddle, 1996. Assessment of two bioassay techniques for resistance monitoring of silverleaf whitefly (Homoptera: Aleyrodidae) in California. J. Econ. Entomol., 89: 805-815.
CrossRefDirect Link - Rauch, N. and R. Nauen, 2003. Identification of biochemical markers linked to neonicotinoid cross resistance in Bemisia tabaci (Hemiptera: Aleyrodidae). Arch. Insect Biochem. Physiol., 54: 165-176.
CrossRefPubMedDirect Link - Roditakis, E., M. Grispou, E. Morou, J.B. Kristoffersen and N. Roditakis et al., 2009. Current status of insecticide resistance in Q biotype Bemisia tabaci populations from Crete. Pest. Manage. Sci., 65: 313-322.
CrossRefPubMedDirect Link - Sayyed, A.H., M.N.R. Attique, A. Khaliq and D.J. Wright, 2005. Inheritance of resistance and cross-resistance to deltamethrin in Plutella xylostella (Lepidoptera: Plutellidae) from Pakistan. Pest Manage. Sci., 61: 636-642.
CrossRefPubMedDirect Link - Shah, R., K. Armstrong, S.P. Worner and R.B. Chapman, 2002. Investigation of a PCR-based method for insecticide resistance monitoring. Pak. J. Biol. Sci., 5: 1070-1073.
CrossRefDirect Link - Soleno, J., L. Anguiano, A.P. de D'Angelo, L. Cichon, D. Fernandez and C. Montagna, 2008. Toxicological and biochemical response to azinphos-methyl in Cydia pomonella L. (Lepidoptera: Tortricidae) among orchards from the Argentinian Patagonia. Pest. Manage. Sci., 64: 964-970.
CrossRefPubMedDirect Link - Torres-Vila, L.M., M.C. Rodriguez-Molina, A. Lacasa-Plasencia and P. Bielza-Lino, 2002. Insecticide resistance of Helicoverpa armigera to endosulfan, carbamates and organophosphates: The Spanish case. Crop Prot., 21: 1003-1013.
CrossRefDirect Link - Unlu, L., 2004. The prediction for the infestation ratio of pink bollworm and spiny bollworm through the examination of blind bolls. Pak. J. Biol. Sci., 7: 2031-2033.
CrossRefDirect Link - Van Asperen, K., 1962. A study of housefly esterases by means of a sensitive colorimetric method. J. Insect Physiol., 8: 401-416.
CrossRefDirect Link - Wyss, C.F., H.P. Young, J. Shukla and R.M. Roe, 2003. Biology and genetics of a laboratory strain of the tobacco budworm, Heliothis virescens (Lepidoptera: Noctuidae), highly resistant to spinosad. Crop Prot., 22: 307-314.
CrossRefDirect Link