
R. Singh
Laboratory of Microbiology, Department of Botany, Christ Church College, Kanpur, 208001, India
V. Kapoor
Laboratory of Microbiology, Department of Botany, Christ Church College, Kanpur, 208001, India
V. Kumar
Laboratory of Microbiology, Department of Botany, Christ Church College, Kanpur, 208001, India
Asian Journal of Biotechnology
Year: 2011 | Volume: 3 | Issue: 6 | Page No.: 540-553







ABSTRACT
The present study deals with the standardization of different cultural conditions for extracellular α-amylase production by thermophilic Streptomyces sp. MSC702 in submerged fermentation (SmF). The appropriate incubation period (48 h), temperature (50°C) and pH (7.0) were determined. The effects of derived and natural carbon sources, inorganic and organic nitrogen sources were also examined. Maximum α-amylase production i.e., 435.71 and 373.89 U mL-1 were achieved by employing derived (D-inositol) and natural (rice bran) carbon sources, respectively. Among the tested nitrogen sources, ammonium sulphate and peptone were found the best inorganic and organic sources, respectively. The C:N ratio found to be the optimum was 1:1. The highest α-amylase activity (807.64 U mL-1) was obtained by utilizing rice bran and wheat bran in 1:2 ratio as the substrate with supplements of D-inositol (1% w/v), ammonium sulphate (0.5% w/v) and peptone (1% w/v). By using the optimized cultural conditions with further characterization, this α-amylase may be utilized in wide spread applications like detergent, saccharification and pharmaceutical industry.
PDF Abstract XML References Citation
Received: May 15, 2011;
Accepted: September 16, 2011;
Published: December 07, 2011
How to cite this article
R. Singh, V. Kapoor and V. Kumar, 2011. Influence of Carbon and Nitrogen Sources on the α-amylase Production by a Newly Isolated Thermophilic Streptomyces sp. MSC702 (MTCC 10772). Asian Journal of Biotechnology, 3: 540-553.
DOI: 10.3923/ajbkr.2011.540.553
URL: https://scialert.net/abstract/?doi=ajbkr.2011.540.553
DOI: 10.3923/ajbkr.2011.540.553
URL: https://scialert.net/abstract/?doi=ajbkr.2011.540.553
INTRODUCTION
Programs to select new microorganisms for enzyme production are increasing around the world. Thermophilic actinomycetes are an integral part of the indigenous soil microflora involved in the decomposition of plant materials where temperature reaches up to 50-60°C such as hay, bagasse, compost etc. On the basis of taxonomical position these microorganisms is well defined in the prokaryote group; they are filamentous, gram-positive, aerobic and chemo-organotrophic bacteria (Stamford et al., 2001). They are one of the most investigated groups since they constitute a potential source of biotechnologically interesting substances (Lealem and Gashe, 1994; Arunachalam et al., 2010).
In the present day scenario, α-amylase (EC 3.2.1.1, 1,4-α-D-glucan glucanohydrolase) find application in all the industrial processes such as in food, detergents, textiles, paper industry, pharmaceutical and fine chemical industries for the hydrolysis of starch (Pandey et al., 2000; Gupta et al., 2003; Sivaramakrishnan et al., 2006). Microbial α-amylases have completely replaced chemical hydrolysis in the starch processing industry (Gupta et al., 2003). It is desirable that α-amylases should be active at the high temperatures of gelatinization (100-110°C) and liquefaction (80-90°C) to economize processes; therefore, there has been a need and continual search for more thermostable α-amylases (Rasooli et al., 2008).
Contemporary of this, microbial α-amylases can also find applications in the bioethanol production method to release low molecular weight sugars from stored starches. Bioethanol (most common biofuel worldwide) is an biologically produced alcohol by the action of microorganisms and enzymes through fermenting the sugar components of plant materials and it is made mostly from sugar and starch crops such as wheat, corn, sugar beets, sugar cane etc. (Hahn-Hagerdal et al., 2006). However, these raw materials base which also have to be used for animal feed and human needs, will not be sufficient to meet the increasing demand for fuel ethanol. Both these factors call for the exploitation of lignocellulose feedstocks, such as agricultural and forest residues as well as dedicated crops, for the production of ethanol (Farrell et al., 2006). Some starchy residues (agricultural residues i.e., rice/wheat straw and bran, corn straw and sugarcane bagasse etc.) are easily obtainable and appear to be the cheapest raw material for ethanol production (Chum and Overend, 2001; Kim and Dale, 2004; Demirbas, 2005; Uma et al., 2010; Roslan et al., 2011). A high degree of gelatinization and liquefaction processes are occurred in the initial step of production of ethanol from starch or starchy residues. So, there is a need of thermostable α-amylases to carry out these processes (Sodhi et al., 2005). Thermophilic actinomycetes including Thermomonospora and Thermoactinomyces are versatile producers of the thermostable α-amylases (Reddy et al., 2003).
Because of the industrial importance of thermostable α-amylases, there is ongoing interest in the isolation of new amylolytic strains of thermophilic actinomycetes. In order to develop an efficient production of α-amylases, knowledge regarding the nutritional factors affecting this process needs to be well identified. Carbon and nitrogen are essential components of the nutrient medium for the growth of the microorganisms (Narayana and Vijayalakshmi, 2008). The use of agricultural residues as carbon source in culture media leads to reduction in the cost of enzyme production (Tangerdy and Szakacs, 2003). In the present study, experimental selection for the screening of best carbon and nitrogen sources were done to achieve cost-effective optimal α-amylase production by newly isolated Streptomyces sp. MSC702.
MATERIALS AND METHODS
All the media ingredients and reagent chemicals were of analytical grade procured from E-Merck, Hi-Media and Qualigen Chemicals, India, Ltd. Natural carbon sources (sugarcane bagasse, wheat bran, rice bran, corn cob and wheat straw) used as sole carbon source were obtained from the local market, Kanpur, India and used in production medium without any pretreatment. All experiments were carried out in triplicate and average values are given in presented data. All the experiments were conducted in year 2010.
Microorganism and growth conditions: The microorganism used in this study was Streptomyces sp. MSC702 originally isolated from the mushroom compost collected from Mushroom Research and Development Centre, Plant Pathology Department, Chandra Shekhar Azad University of Agriculture and Technology, Kanpur, India. The stock culture of Streptomyces sp. MSC702 was maintained on M medium (Obi and Odibo, 1984) modified with 1% (v/v) trace element solution (Techapun et al., 2002) at 50°C.
Strain isolation: Thermophilic strains of actinomycetes were isolated from samples of mushroom compost on petri plates of modified M medium using dilution plate technique (Wakesman, 1927) at 45°C. Probable amylase producing strains were then selected by flooding the plates with 0.001 M iodine solution (Obi and Odibo, 1984). Subsequently, the amylase producing strain was cultivated in modified M medium agar slants for 2 days at 50°C and stored at 4°C.
Selection of the working strain: Each of 7 probable amylase producing strains was separately inoculated into 100 mL of M broth (Obi and Odibo, 1984) in 250 mL Erlenmeyer flask and allowed to ferment for 48 h at 50°C (pH 7.0). Contents were filtered with Whatman filter papers 1 (qualitative circles, 125 mm diameter) and centrifuged at 5000 g for 20 min at 4°C and the clear cell-free supernatant was used for α-amylase assay.
Preparation of inoculums: The inoculum was prepared by adding 10 mL sterile distilled water in to 2 days old culture slant and make a cell suspension with the help of sterile loop. The spore suspension was taken and agitated in a vortex-cyclomixer for 5 min in order to disperse the spores evenly. The cells (1.9-2.2x108 CFU mL-1) from these slants were inoculated into production media.
Biomass production: To determine the total amount of growth (biomass), the cells were harvested at the interval of 12 h for 96 h. The mycelial mass was removed by vacuum filtration and dried overnight in oven at 80°C to measure dry biomass weight, expressed in terms of mg dry mass 100 mL-1 of culture medium.
Production of α-amylase at different physiological conditions: Influence of incubation period (12 to 96 h), temperature (45 to 70°C) and pH (5.0 to 10.0) on α-amylase production by strain MSC702 was studied in 1% (w/v) starch supplemented production medium. The production medium (100 mL) was prepared according to Obi and Odibo (1984), were taken in 250 mL of Erlenmeyer flasks and autoclaved at 121°C (15 lbs) for 20 min and cooled. Production medium then inoculated with 1% (v/v) inoculum. The pH of the medium was adjusted using 1 N HCl or 1 N NaOH.
Production of α-amylase at different carbon and nitrogen sources: The influence of different carbon sources on the α-amylase production by strain MSC702 was studied by replacing 1% (w/v) starch with the other derived and natural carbon sources in production medium. The influence of the nitrogen sources on the α-amylase production was studied by replacing the 0.1% (w/v) (NH4)2SO4 with other nitrogen sources in production medium containing rice bran and wheat bran in 1:2 ratio as sole carbon source. The effect of C: N ratio on α-amylase biosynthesis was studied using starch as a fixed carbon source with varying nitrogen source (peptone) in different concentration. The initial pH of the production medium was 7.0. Production medium then inoculated with 1% (v/v) inoculum and incubated at 50°C for 48 h. The saccharolytic activity of α-amylase was determined by 3, 5-dinitrosalicylic acid method (Miller, 1959).
Analytical procedure: The reaction mixture contained 0.1 mL of appropriately diluted supernatant enzyme and 0.9 mL of 0.1 M phosphate buffer (pH 7.0) containing 1% (w/v) of soluble starch. The mixture was incubated at 50°C for 10 min. The amount of reducing sugar level released in the mixture was determined by the 3, 5-dinitrosalicylic acid method (Miller, 1959). The absorbance was measured at 550 nm using UV-visible spectrophotometer (UV-1700 PharmaSpec Shimadzu). One unit of α-amylase activity was defined as the amount of enzyme that released 1 μmol of reducing sugars (maltose) per minute under the standard assay conditions.
Production of α-amylase under optimum conditions: On the basis of the results obtained with all the optimum parameters, namely viz., rice bran and wheat bran in 1:2 ratio containing D-inositol (1% w/v) as supplementary carbon source and peptone (1% w/v) and ammonium sulphate (0.5% w/v) as supplementary nitrogen sources, pH of 7.0, incubation temperature of 50°C and incubation period of 48 h, the extent of improvement of α-amylase biosynthesis in the optimized medium was evaluated and compared with 1% (w/v) starch supplemented production medium as a control. The fermentation was carried out as described above.
RESULTS AND DISCUSSION
Selection and identification of working strain: During the screening process of seven amylolytic strains isolated from mushroom compost, strain MSC702 was chosen as working strain on the basis of the highest α-amylase production amongst all. The newly isolated strain was spore forming, Gram-positive, highly thermophilic, halophilic and alkaliphilic in nature (data not shown). The phenotypic and phylogenetic (data not shown) properties of strain MSC702 are consistent with its assignments to the genus Streptomyces. The data related to comparison of 16S rRNA sequence of strain MSC702 with other related Streptomyces spp. was described in Table 1.
Strain MSC702 was deposited in the Microbial Type Culture Collection and Gene Bank (MTCC), Chandigarh, India with accession number MTCC 10772. An almost complete 16S rRNA sequence of strain MSC702 (1474 nucleotides) has been deposited in the GenBank database with nucleotide sequence Accession No. JF325872.
Effect of incubation period, temperature and pH on α-amylase production: Strain MSC702 showed maximum α-amylase production (276.34 U mL-1, 48 h) in early log phase with drastic decrease in production of enzyme (232.56 U mL-1, 72 h) at late growth phase or early stationary phase (Fig. 1). The production of α-amylase by strain MSC702 began after 12 h and reached a peak of 276.34 U mL-1 at 48 h. Result showed that α-amylase production was independent of growth phase. Similar reports on α-amylase were reported on Streptomyces albidoflavus (Narayana and Vijayalakshmi, 2008) and S. erumpens MTCC 7317 (Kar and Ray, 2008).
The incubation period is governed by characteristics of the culture and is based on growth rate and enzyme production. In most cases, the period employed was 30 h, although an average of 48-50 h is common (Lonsane and Ramesh, 1990). The production of α-amylase by strain MSC702 began after 12 h and reached a peak of 276.34 U mL-1 at 48 h.
| Table 1: | Details of Streptomyces spp. closely related to Streptomyces sp. MSC702 on the basis of 16S rRNA gene sequence similarity |
 | |
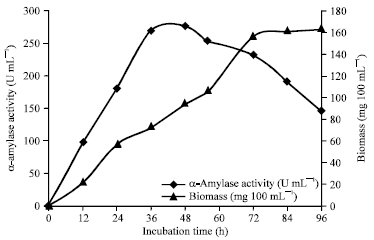 | |
| Fig. 1: | Growth characteristics with reference to α-amylase production at the interval of 12 h by the strain MSC702 (pH 7.0, at 50°C) |
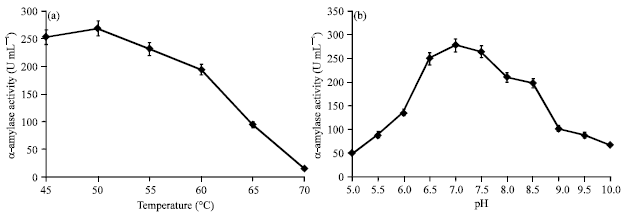 | |
| Fig. 2 (a-b): | α-Amylase production by Streptomyces sp. MSC702 at different physical parameters. (a) Effect of different temperature range (45-70°C) at pH 7.0 and 48 h incubation and (b) Effect of different pH range (5.0-10.0) for 48 h incubation at 50°C |
Similar results were obtained by Baysal et al. (2003) and Liu and Yan (2008) with Bacillus spp. Yang and Wang (1999) studied the influence of incubation period on α-amylase produced by Streptomyces rimosus. The production of α-amylase began after 24 h of cultivation and reached to peak levels after 48 h and declined thereafter. This result showed the good agreement with the present study.
Optimum amylase production at 45-55°C is characteristic of the moderately thermostable actinomycetes like Thermomonospora fusca (Busch and Stutzenberger, 1997), Streptomyces sp. IMD 267 (McMahon et al., 1997), Streptomyces sp. D1 (Chakraborty et al., 2009) and S. erumpens MTCC 7317 (Kar and Ray, 2008), beyond 55°C, there was sudden decrease in enzyme production. Streptomyces sp. MSC702 showed highest (269.78 U mL-1) α-amylase activity at 50°C (Fig. 2a) with retaining ~72% (195.16 U mL-1) and ~35% (95.29 U mL-1) α-amylase production at 60°C and 65°C, respectively. Many Bacillus sp. such as Bacillus amyloliquefaciens, B. subtilis and B. licheniformis reported for α-amylase production at same temperature range (37-60°C) (Syu and Chen, 1997; Mishra et al., 2005; Mendu et al., 2005).
Present study showed the optimum pH range 6.5-7.5 for α-amylase biosynthesis by strain MSC702 with decrease in production thereafter (Fig. 2b). Similarly, Kuo and Hartman (1966) found that Thermoactinomyces vulgaris synthesized amylase most rapidly at pH values ranging from 6.5 and 7.5 and that amylase inactivation occurred rapidly if pH rose above 7.5. Amylase production by Streptomyces aureofaciens 77 (Shatta et al., 1990), S. erumpens MTCC 7317 (Kar and Ray, 2008) and S. rimosus (Yang and Wang, 1999) has been increased gradually as the initial pH values ascend from 5.0 to 7.0. These results agreed with our results where maximum α-amylase production (278.77 U mL-1) achieved at pH 7.0.
Effect of various carbon sources on α-amylase production: Growth and enzyme production of any organism are greatly influenced by both environmental conditions as well as the nutrients available in the growth medium. To investigate the effects of various carbon sources, Streptomyces sp. MSC702 was inoculated in the production medium containing 1% (w/v) derived [(monosaccharides = arabinose, xylose, glucose, fructose, galactose); (disaccharides = sucrose, maltose, lactose, cellobiose); (trisaccharides = raffinose); (polysaccharides = starch, cellulose); (deoxy sugar = L-rhamnose); (polyhydroxy alcohols = D-sorbitol, D-mannitol, D-inositol)] and natural carbon sources (sugarcane bagasse, wheat bran, rice bran, corn cob and wheat straw) separately for 48 h. Results showed that all the substrates supported α-amylase production by the strain, except arabinose, xylose and galactose. The combination of D-inositol with the mixture of rice bran and wheat bran proved superior to the other composition.
Derived carbon sources: All the tested carbon sources supported the growth of strain MSC702. In present investigation, there was no correlation between biomass yield and biosynthesis of α-amylase by the strain (Fig. 3). Glucose, fructose, sucrose, maltose, lactose, cellobiose, raffinose, starch, cellulose, D-sorbitol, D-mannitol and D-inositol were found to support α-amylase biosynthesis, whereas arabinose, xylose and galactose showed a repressive effect on α-amylase production (Fig. 3). L-rhamnose showed no significant effect on α-amylase biosynthesis. All the tested polyhydroxy alcohols favored α-amylase production. The maximum enzyme level was obtained in the medium containing D-inositol (435.71 U mL-1). The results obtained are in good agreement with the findings of Aiyer (2004) who also stated that amylase synthesis occurred in all the carbon sources except arabinose and galactose, both of them were least effective for amylase production. Srivastava and Baruah (1986) also found that inositol and sorbitol favors the growth and production of amylase by Bacillus stearothermophilus.
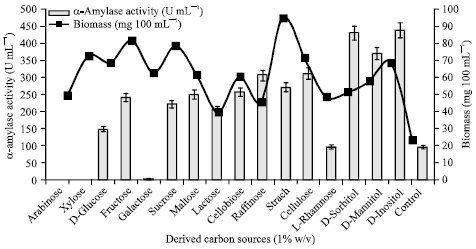 | |
| Fig. 3: | Effect of derived carbon sources (1% w/v) on cell growth and α-amylase production |
Natural carbon sources: To evaluate the effect of natural carbon sources on the production of α-amylase by Streptomyces sp. MSC702, fermentation was done with the addition of 1% (w/v) Sugarcane Bagasse (SB), Wheat Bran (WB), Rice Bran (RB), Corn Cob (CC) and Wheat Straw (WS) separately in the production medium. The order of substrate suitability was RB>CC>WB > WS>SB (Fig. 4). The α-amylase activities obtained with these substrates were: 373.89 U mL-1 with rice bran; 368.87 U mL-1 with corn cob; 261.48 U mL-1 with wheat bran; 234.54 U mL-1 with wheat straw and 201.14 U mL-1 with sugarcane bagasse. The utilization of agricultural residues for production of enzymes was also studied by Milala et al. (2009).
Therefore, the volumetric activities were quite different in the presence of the various derived and natural carbon sources. Carbohydrate present in the greatest proportion in the natural carbon sources are in the form of lignocellulosics and starch. Rice bran was the best carbon source for α-amylase production, contain 65% starch. On the other hand, corn cob, wheat bran, wheat straw and sugarcane bagasse has lesser percentage of starch. Therefore, better α-amylase production may be correlated with the higher content of starch in agro-residues. Due to the higher volumetric activity obtained with rice bran, it was maintained in the fermentation medium for the remaining experiments but it is important to note that combinations of the rice bran with other natural carbon sources, may be a good alternative for higher α-amylase production and due to the less expensive in nature, natural carbon source, could also be used for industrial scale production.
Combinations of natural carbon sources: Figure 4 and 5, show the effect of different combination (1:1 w/v) of natural carbon sources on α-amylase production taking rice bran as fixed substrate in all combinations. In the present study, out of the different combinations (RB:SB; RB:WB; RB:CC; RB:WS) tested, α-amylase activity reached up to 398.1 U mL-1 in rice bran and wheat bran (1:1 w/v) containing medium. Ten different combinations (0.5:0.5; 1:1; 1:2; 1:3; 1:4; 1:5; 2:1; 2:1; 4:1 and 5:1) of rice bran and wheat bran were analyzed to maximize α-amylase production. The highest α-amylase activity was found at the level from mixed substrates of rice bran: wheat bran in 1:2 ratio. Yousaf et al. (2008) were also reported similar result (rice bran to wheat bran 1:2; amylase activity ~180 U mL-1). Jecu (2000) also reported the importance of mixed substrate (wheat bran and wheat straw) for endoglucanase production by Aspergillus niger.
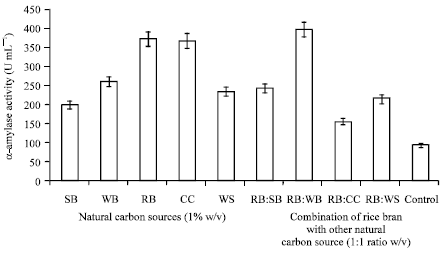 | |
| Fig. 4: | Effect of natural carbon sources (1% w/v) and combinations of rice bran with other natural carbon sources (1:1 ratio w/v) on α-amylase production |
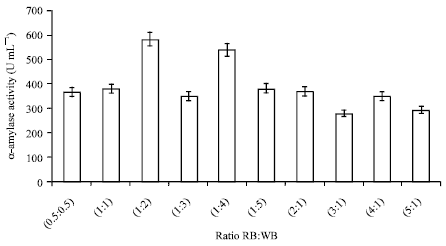 | |
| Fig. 5: | Effect of different ratios of rice bran and wheat bran on α-amylase production |
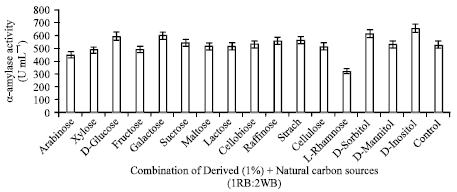 | |
| Fig. 6: | Effect of combination of derived and natural carbon sources (1RB: 2WB) on α-amylase |
Combination of derived and natural carbon sources: The combined effect of different derived carbon sources (1% w/v) with rice bran: wheat bran (1:2) was studied by their addition to production medium (Fig. 6). D-inositol gave significantly higher (665.37 U mL-1) yield of α-amylase than other carbon sources when added to production medium containing rice bran and wheat bran in 1:2 ratio. Similar results shown by Srivastava and Baruah (1986), they also observed the effect of inositol on amylase production by Bacillus stearothermophilus.
Effect of nitrogen sources on α-amylase production: A complex nitrogen source appears to be essential for the production of high amount of α-amylase by Streptomyces sp. MSC702. The requirement may be explained by organism’s preference for protein as a nitrogen source for growth as well as for extracellular enzyme production. The amylase synthesis by several microorganisms has been correlated to the presence or absence of various amino acids and complex nitrogen sources in the culture medium (Fogarty and Kelly, 1980; Hillier et al., 1997). In present study, the tested nitrogen sources of inorganic and organic nitrogen were NaNO3, (NH4)2SO4, NH4NO3, (NH4)2HPO4, urea, thiourea, peptone, yeast extract, tryptose, casein and amino acids. Among the tested nitrogen sources, ammonium sulphate was the best inorganic nitrogen source followed by ammonium nitrate (Fig. 7). While peptone was the best organic nitrogen source with 9.0% more yield than ammonium sulphate. α-Amylase production in presence of peptone as a nitrogen source was 584.38 U mL-1. The effects of different concentrations of peptone and ammonium sulphate on α-amylase production were also studied.
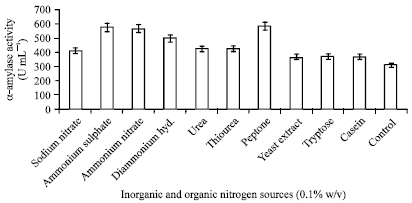 | |
| Fig. 7: | Effect of inorganic and organic nitrogen sources (0.1% w/v) on α-amylase production |
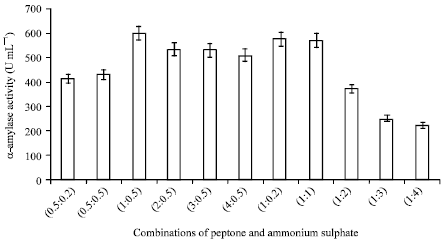 | |
| Fig. 8: | Effect of combinations of peptone and ammonium sulphate with different concentration on α-amylase production |
Results revealed that 1.0% of peptone with 0.5% ammonium sulphate gave highest α-amylase yield (Fig. 8). In the present study, ammonium sulphate stimulate α-amylase synthesis with certain amount, on increasing the amount, it reduced the α-amylase synthesis. Similar result was reported by Mai et al. (1992), who also observed stimulation of amylase activity by ammonium salts. Aiyer (2004) also reported the enhancement of amylase production by using the combination of peptone with ammonium hydrogen phosphate. In contrast, Saxena et al. (2007) reported inhibitory effect of ammonium salts on amylase production.
Among the seventeen amino acids examined, only seven (Table 2) have stimulatory effect on α-amylase production. Glycine, DL-aspartic acid and L-glutamic acid were found to be indispensable for α-amylase production. Glycine (138.82 U mL-1) was found to be most suitable amino acid for the production of α-amylase. The lowest value (62.38 U mL-1) of α-amylase production was obtained in presence of threonine. Similar results have been obtained by other workers (Chandra et al., 1980; Srivastava and Baruah, 1986). α-Amylase production by Bacillus amyloliquefaciens ATCC 23350 increased by a factor of 300 in the presence of glycine (Zhang et al., 1983). The effect of glycine was not only as a nitrogen source rather it affected α-amylase production by controlling pH and subsequently α-amylase production increased. DL-Alanine, DL-valine and DL-methionine were effective for the production of alkaline amylase by Bacillus sp. However, the role of amino compounds was considered to be neither as nitrogen nor as a carbon source but as stimulators of amylase synthesis and excretion (Ikura and Horikoshi, 1987).
| Table 2: | Effect of different amino acids on α-amylase production |
 | |
| Control*-medium without amino acid | |
| Table 3: | Effect of C: N ratio on α-amylase production |
 | |
Kundu et al. (1973) reported that only asparagines gave good enzyme yields while the importance of arginine for α-amylase production from B. subtilis has also been well documented (Lee and Parulekar, 1993).
Effect of C:N ratio on α-amylase production: For C:N ratio study, soluble potato starch and peptone were used as carbon and nitrogen sources, respectively. 1% (w/v) of potato starch was used while the amount of peptone was varied (0.5, 1.0, 1.5, 2.0, 2.5 and 3.0% w/v) to attain desired C:N ratio. The activity of α-amylase in fermented broth was recorded after 48 h of incubation. The maximum α-amylase yield (605.80 U mL-1) was obtained when C:N ratio was 1.0 (Table 3). Aiyer (2004) worked on Bacillus licheniformis SPT 27 showed similar results and having optimum production of amylase in 1:1 C:N ratio where potato soluble starch and peptone were used as carbon and nitrogen sources, respectively. McTigue et al. (1995) reported C:N ratio of 1:1 with starch and yeast extract as carbon and nitrogen sources, respectively.
Production of α-amylase under optimized conditions: A high level of α-amylase production (807.64 U mL-1) was achieved by utilizing rice bran and wheat bran in 1:2 ratio as the substrate with supplements (w/v) of D-inositol (1%), peptone (1%) and ammonium sulphate (0.5%) for 48 h at 50°C, pH of 7.0.
CONCLUSIONS
In the present investigation, we have isolated thermostable α-amylase producing strain Streptomyces sp. MSC702 from mushroom compost. From the above mentioned results, we concluded that ~3 fold increase in α-amylase production was achieved under the optimized fermentation conditions with (w/v) 1% D-inositol, 1% peptone and 0.5% ammonium sulphate as supplements, as compared with the production medium with 1% starch. The enzyme in this research performed some differences in properties related to the production medium which are valuable for higher industrial α-amylase production. The study also concluded that the suitability of newly isolated strain of thermophilic actinomycete Streptomyces sp. MSC702 in the degradation of agro-residues for the release of sugars in bioethanol production. Application of agro-residues in bioprocesses not only provides alternative substrates but it also helps solving their disposal problem.
ACKNOWLEDGMENT
The authors are grateful to the University Grant Commission, Selection and Award Bureau, New Delhi, India for providing financial support to carry out this work and awarding a fellowship to Renu Singh under the scheme of ‘Rajiv Gandhi National Fellowship’.
REFERENCES
- Aiyer, P.V.D., 2004. Effect of C: N ratio on alpha amylase production by Bacillus licheniformis SPT 27. Afr. J. Biotechnol., 3: 519-522.
Direct Link - Baysal, Z., F. Uyar and C. Aytekin, 2003. Solid state fermentation for production of α-amylase by a thermotolerant Bacillus subtilis from hot-spring water. Process Biochem., 38: 1665-1668.
CrossRefDirect Link - Busch, J.E. and F.J. Stutzenberger, 1997. Amylolytic activity of Thermomonospora fusca. World J. Microbiol. Biotechnol., 13: 637-642.
CrossRef - Chakraborty, S., A. Khopade, C. Kokare, K. Mahadik and B. Chopade, 2009. Isolation and characterization of novel α-amylase from marine Streptomyces sp. D1. J. Mol. Cataly. B: Enzym., 58: 17-23.
CrossRef - Chandra, A.K., S. Medda and A.K. Bhadra, 1980. Production of extracellular thermostable α-amylase by Bacillus licheniformis. J. Ferment. Technol., 58: 1-10.
Direct Link - Chum, H.L. and R.P. Overend, 2001. Biomass and renewable fuels. Fuel Process. Technol., 71: 187-195.
CrossRefDirect Link - Demirbas, A., 2005. Bioethanol from cellulosic materials: A renewable motor fuel from biomass. Energy Sources, 27: 327-337.
CrossRefDirect Link - Farrell, A.E., R.J. Plevin, B.T. Turner, A.D. Jones, M. O'Hare and D.M. Kammen, 2006. Ethanol can contribute to energy and environmental goals. Science, 311: 506-508.
CrossRefDirect Link - Gupta, R., P. Gigras, H. Mohapatra, V.K. Goswami and B. Chauhan, 2003. Microbial α-amylases: A biotechnological perspective. Process Biochem., 38: 1599-1616.
CrossRefDirect Link - Hillier, P., D.A.J. Wase, A.N. Emery and G.L. Solomons, 1997. Instability of α-amylase production and morphological variation in continuous culture of Bacillus amyloliquefaciens is associated with plasmid loss. Process Biochem., 32: 51-59.
CrossRef - Ikura, Y. and K. Horikoshi, 1987. Effect of amino compounds on alkaline amylase production by alkalophilic Bacillus sp. J. Ferment. Technol., 65: 707-709.
CrossRef - Jecu, L., 2000. Solid state fermentation of agricultural wastes for endoglucanase production. Ind. Crops Prod., 11: 1-5.
Direct Link - Kim, B., A.M. Al-Tai, K.S. Bum, P. Somasundaram and M. Goodfellow, 2000. Streptomyces thermocoprophilus sp. nov., a cellulase-free endo-xylanase-producing streptomycete. Int. J. Syst. Evol. Microbiol., 50: 505-509.
Direct Link - Kim, S. and B.E. Dale, 2004. Global potential bioethanol production from wasted crops and crop residues. Biomass Bioenergy, 26: 361-375.
CrossRefDirect Link - Kuo, M.J. and P.A. Hartman, 1966. Isolation of amylolytic strains of Thermoactinomyces vulgaris and production of thermophilic actinomycete amylases. J. Bacteriol., 92: 723-726.
Direct Link - Lee, J. and S.J. Parulekar, 1993. Enhanced production of α-amylase in fed-batch cultures of Bacillus subtilis TN106[pAT5]. Biotechnol. Bioeng., 42: 1142-1150.
CrossRef - Liu, X.D. and Y. Xu, 2008. A novel raw starch digesting α-amylase from a newly isolated Bacillus sp. YX-1. Purification and characterization. Bioresour. Technol., 99: 4315-4320.
CrossRefPubMedDirect Link - Lonsane, B.K. and M.V. Ramesh, 1990. Production of bacterial thermostable α-amylase by solid state fermentation: A potential tool for achieving economy in enzyme production and starch hydrolysis. Adv. Applied Microbiol., 35: 1-56.
CrossRef - Mai, N.T., D.T. Giang, N.T.N. Minh and V.T. Thao, 1992. Thermophilic amylase-producing bacteria from Vietnamese soils. World J. Microbiol. Biotechnol., 8: 505-508.
CrossRef - Milala, M.A., B.B. Shehu, H. Zanna and V.O. Omosioda, 2009. Degradation of agro-waste by cellulase from Aspergillus candidus. Asian J. Biotechnol., 1: 51-56.
CrossRefDirect Link - McMahon, H.E.M., C.T. Kelly and W.M. Fogarty, 1997. Effect of growth rate on α-amylase production by Streptomyces sp. IMD 2679. Applied Microbiol. Biotechnol., 48: 504-509.
Direct Link - McTigue, M.A., C.T. Kelly, E.M. Doyle and W.M. Fogarty, 1995. The alkaline amylase of the alkalophilic Bacillus sp. IMD 370. Enzyme Microbial Technol., 17: 570-573.
CrossRef - Mendu, D.R., B.V.V. Ratnam, A. Purnima and C. Ayyanna, 2005. Affinity chromatography of α-amylase from Bacillus licheniform. Enzyme Microb. Technol., 37: 712-717.
CrossRef - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - Mishra, S., S.B. Noronha and G.K. Suraishkumar, 2005. Increase in enzyme productivity by induced oxidative stress in Bacillus subtilis cultures and analysis of its mechanism using microarray data. Process Biochem., 40: 1863-1870.
CrossRef - Narayana, K.J.P. and M. Vijayalakshmi, 2008. Production of extracellular α-amylase by Streptomyces albidoflavus. Asian J. Biochem., 3: 194-197.
CrossRefDirect Link - Obi, S.K.C. and F.J.C. Odibo, 1984. Partial purification and characterization of a thermostable actinomycete β-amylase. Applied Environ. Microbiol., 47: 571-575.
Direct Link - Pandey, A., P. Nigam, C.R. Soccol, V.T. Soccol, D. Singh and R. Mohan, 2000. Advances in microbial amylases. Biotechnol. Applied Biochem., 31: 135-152.
PubMedDirect Link - Petrosyan, P., M. Garcia-Varela, A. Luz-Madrigal, C. Huitron and M.E. Flores, 2003. Streptomyces mexicanus sp. nov., a xylanolytic micro-organism isolated from soil. Int. J. Syst. Evol. Microbiol., 53: 269-273.
CrossRefDirect Link - Pridham, T.G., C.W. Hesseltine and R.G. Benedict, 1958. A guide for the classification of streptomycetes according to selected groups; placement of strains in morphological sections. Applied Microbiol., 6: 52-79.
PubMedDirect Link - Rasooli, I., S.D.A. Astaneh, H. Borna and K.A. Barchini, 2008. A thermostable α-amylase producing natural variant of Bacillus spp. isolated from soil in Iran. Am. J. Agric. Biol. Sci., 3: 591-596.
Direct Link - Reddy, N.S., A. Nimmagadda and K.R.S.S. Rao, 2003. An overview of the microbial α-amylase family. Afr. J. Biotechnol., 2: 645-648.
Direct Link - Roslan, A.M., P.L. Yee, U.K.M. Shah, S.A. Aziz and M.A. Hassan, 2011. Production of bioethanol from rice straw using cellulase by local Aspergillus sp. Int. J. Agric. Res., 6: 188-193.
CrossRefDirect Link - Saxena, R.K., K. Dutt, L. Agarwal and P. Nayyar, 2007. A highly thermostable and alkaline amylase from a Bacillus sp. PN5. Bioresour. Technol., 98: 260-265.
CrossRefDirect Link - Srivastava, R.A.K. and J.N. Baruah, 1986. Culture conditions for production of thermostable amylase by Bacillus stearothermophilus. Applied Environ. Microbiol., 52: 179-184.
Direct Link - Stamford, T.L.M., N.P. Stamford, L.C.B.B. Coelho and J.M. Araujo, 2001. Production and characterization of a thermostable α-amylase from Nocardiopsis sp. endophyte of yam bean. Bioresource Technol., 76: 137-141.
CrossRef - Syu, M.J. and Y.H. Chen, 1997. A study on the α-amylase fermentation performed by Bacillus amyloliquefaciens. Chem. Eng. J., 65: 237-247.
Direct Link - Techapun, C., T. Charoenrat, N. Poosaran, M. Watanabe and K. Sasak, 2002. Thermostable and alkaline-tolerant cellulase-free xylanase produced by thermotolerant Streptomyces sp. Ab106. J. Biosci. Bioeng., 93: 431-433.
CrossRef - Tengerdy, R.P. and G. Szakacs, 2003. Bioconversion of lignocellulose in solid substrate fermentation. Biochem. Eng. J., 13: 169-179.
CrossRef - Trejo, W.H., L.D. Dean, J. Pluscec, E. Meyers and W.E. Brown, 1977. Streptomyces laurentii, a new species producing thiostrepton. J. Antibiot., 30: 639-643.
PubMed - Uma, C., C. Muthulakshmi, D. Gomathi and V.K. Gopalakrishnan, 2010. Fungal invertase as aid for production of ethanol from sugarcane bagasse. Res. J. Microbiol., 5: 980-985.
CrossRefDirect Link - Yang, S.S. and J.Y. Wang, 1999. Protease and amylase production of Streptomyces rimosus in submerged and solid state cultivations. Bot. Bull. Acad. Sin., 40: 259-265.
Direct Link - Zhang, Q., N. Tsukagoshi, S. Miyashiro and S. Udaka, 1983. Increased production of α-amylase by Bacillus amyloliquefaciens in the presence of glycine. Applied Environ. Microbiol., 46: 293-295.
PubMed - Syu, M.J. and Y.H. Chen, 1997. A study on the α-amylase fermentation performed by Bacillus amyloliquefaciens. Chem. Eng. J., 65: 237-247.
Direct Link - Arunachalam, R., E.G. Wesely, J. George and G. Annadurai, 2010. Novel approaches for identification of Streptomyces noboritoensis TBG-V20 with cellulase production. Curr. Res. Bacteriol., 3: 15-26.
CrossRefDirect Link - Sodhi, H.K., K. Sharma, J.K. Gupta and S.K. Soni, 2005. Production of a thermostable α-amylase from Bacillus sp. PS-7 by solid state fermentation and its synergistic use in the hydrolysis of malt starch for alcohol production. Process Biochem., 40: 525-534.
CrossRefDirect Link