
Milad Mohkam
Department of Biology, Faculty of Sciences, University of Isfahan, Isfahan, Iran
Sayyed Hamid Zarkesh-Esfahani
Department of Biology, Faculty of Sciences, University of Isfahan, Isfahan, Iran
Mehdi Fazeli
Department of Basic Sciences, Faculty of Veterinary Medicine, Shiraz University, Shiraz, Iran
Asian Journal of Biotechnology
Year: 2011 | Volume: 3 | Issue: 5 | Page No.: 493-506







ABSTRACT
The aim of this study was to clone and optimize production of a recombinant antibody against human leptin receptor which showed inhibitory effects on leptin signalling. DNA sequence encoding Fab fragment of a mouse monoclonal antibody was cloned into pComb3 vector. E. coli was transformed with the plasmid and recombinant antibody was produced. In order to achieve high level of protein expression, recombinant antibody production was performed using different E. coli strains and culture conditions. Antibody production was detected using ELISA (Enzyme-linked immunosorbent assay), Western blotting and Dot blotting techniques. Recombinant Fab fragment of antibody was produced in E. coli successfully at reasonable quantities with comparable affinity to the antigen compared to antibody produced by original hybridoma cells. The results showed that among different culture media, SB medium was the best medium and among different temperatures 22°C was the best temperature for recombinant antibody production. From three different E. coli strains (JM109. XL1-Blue and BL21) tested, JM109 produced the highest levels and among different IPTG concentrations 1 mM induced the highest level of antibody production.
PDF Abstract XML References Citation
Received: April 19, 2011;
Accepted: June 25, 2011;
Published: July 27, 2011
How to cite this article
Milad Mohkam, Sayyed Hamid Zarkesh-Esfahani and Mehdi Fazeli, 2011. Construction of a Recombinant Fab Fragment of a Monoclonal Antibody against Leptin Receptor. Asian Journal of Biotechnology, 3: 493-506.
DOI: 10.3923/ajbkr.2011.493.506
URL: https://scialert.net/abstract/?doi=ajbkr.2011.493.506
DOI: 10.3923/ajbkr.2011.493.506
URL: https://scialert.net/abstract/?doi=ajbkr.2011.493.506
INTRODUCTION
Leptin, a non-glycosylated protein product of the ob (obese) gene, is a hormone/cytokine synthesized mainly by adipose cells to regulate weight control in a central manner, via its cognate receptor in the hypothalamus (Maffei et al., 1995; Zwick et al., 2001). The correlation of serum leptin levels with body fat mass increased with fat accumulation. Leptin is classically considered as a hormone because it regulates the balance between food intake and energy expenditure (Friedman and Halaas, 1998). On the other hand, leptin is a cytokine as well, as it is structurally similar to the cytokines of the type I cytokine family, characterized by a long-chain four-helical bundle such as growth hormone, prolactin, erythropoietin, interleukin (IL) 3, IL-11 and IL-12. Mutations of the ob gene are associated with immune deficiency and obesity (Farooqi et al., 2002). As a cytokine, leptin significantly increased the responses to acute infection and sepsis and has direct effects on some immunological functions such as CD4+ T lymphocyte propagation, macrophage phagocytosis and secretion of inflammatory cytokines (Loffreda et al., 1998). Leptin promotes a switch toward a Th1 responses, accelerates production of inflammatory cytokines from human monocytes (Zarkesh-Esfahani et al., 2004; Lord et al., 1998) and is permissive to autoimmune disease susceptibility in animal models (Matarese et al., 2005) such as autoimmune encephalomyelitis (Matarese et al., 2001; Sanna et al., 2003), arthritis (Busso et al., 2002), autoimmune diabetes (Matarese et al., 2002) and chronic inflammatory conditions such as atherosclerosis (Correia and Haynes, 2004).
A sexual dimorphism of leptin has been demonstrated in several species (Pardridge, 1986; Smith and Waddell, 2003). In humans, leptin is higher in females than males matched for age and the gender difference has been attributed to higher leptin production in subcutaneous adipose tissue, stimulation of leptin by oestrogen in females and suppression of leptin by testosterone in males. It is also known that autoimmune diseases are more prevalent in female than male (Saad et al., 1997). Leptin signals through a cell surface receptor (ObR) which is a member of the class I cytokine receptor family ( Li and Friedman, 1999; Tartaglia et al., 1995). ObR is expressed in both the nervous system and in peripheral tissues. The relative expression levels of different receptor isoforms vary among tissues, possibly to allow leptin’s biological activity to be more precisely regulated at various leptin target sites (Ahima et al., 2000; Ghilardi et al., 1996). To date, six splice variants of the ObR, “a” to “f”, have been identified but there are two main isoforms of the ObR: ObRb, the long form which has Jak/STAT and MAPK (Signal Transducers and Activators of Transcription - Mitogen-Activated Protein Kinase) signalling capabilities and is present in the hypothalamus and ObRa, the short form which lacks most of the cytoplasmic domain of the receptor but may be capable of signalling through MAPK ( Barr et al., 1999; Tartaglia, 1997). ObR isoforms have a similar extracellular ligand-binding domain at the amino terminus but differ at the intracellular carboxy-terminal domain. ObR is widely expressed in tissues and cells of the immune system, with ObRb expression detected at least in human monocytes and T cells (Zarkesh-Esfahani et al., 2004). Leptin antagonism has been proposed as an immunotherapeutic approach for the treatment of some autoimmune disorders (Farooqi et al., 1999; Landman et al., 2003).
There are several ways to block cytokine receptors from which humanized antibodies have received high attention and some of them are approved by FDA (Food and Drug Administration) for clinical use (Iversen et al., 2002; Sanna et al., 2003).
A prokaryotic system such as E. coli is one of the earliest and most widely used hosts for the production of heterologous proteins including antibodies (Terpe, 2006). This system is excellent for functional expression of non-glycosylated proteins, although there were several reports that for prokaryotic systems which is suitable for recombinant antibody production (Donovan et al., 2000). E. coli genetics are far better understood than those of any other microorganism. Recent progress in the fundamental understanding of transcription, translation and protein folding in E. coli, together with the availability of improved genetic tools, is making this bacterium more valuable than ever for the expression of complex eukaryotic proteins (Swartz, 2001). The E. coli system has some drawbacks, however, which have to be overcome for efficient expression of proteins. Proteins which are produced as inclusion bodies, are often inactive, insoluble and require refolding. In addition, there is a problem producing proteins with many disulfide bonds and refolding these proteins are extremely difficult (Choi and Lee, 2004). To improve the E. coli process situation, the following measures have been taken: (i) use of different promoters to regulate expression; (ii) use of different host strains; (iii) co-expression of chaperones and/ or foldases; (iv) lowering of temperature; (v) secretion of proteins into the periplasmic space or into the medium; (vi) reducing the rate of protein synthesis; (vii) changing the growth medium; (viii) addition of a fusion partner; (ix) expression of a fragment of the protein (Mergulhao et al., 2005; Shiloach and Fass, 2005).
The aim of this study was to clone, express, purify and test binding of a recombinant mouse antibody from hybridoma cells raised against human leptin receptor. The recombinant antibody has the capacity for manipulation and humanization for further applications. As the level of functional protein expression in E. coli is highly variable, in order to develop an efficient bacterial expression system, the most important factors affecting recombinant antibody production in E. coli were also optimised.
MATERIALS AND METHODS
This study was performed during Feb 2007 till Sep 2008.
Cloning of the heavy and light chains of antibody: The heavy and light chains of antibody were cloned in the pComb3 vector which contains two different expression cassettes. The cDNA encoding heavy chain (VH) of antibody was amplified by PCR using the following set of primers: VHb, upstream, 5’-AG GTC CAG CTG CTC GAG TCT GG-3’ and CH1(γ1), downstream, 5’-AGG CTT ACT AGT ACA ATC CCT GGG CAC AAT-3’. The following set of primers was used to PCR amplify the light chain (VL) of antibody: VL (k), upstream, 5’-GT GCC AGA TGT GAG CTC GTG ATG ACC CAG TCT CCA-3’ and CL (k), downstream, 5’-T CCT TCT AGA TTA CTA ACA CTC TCC CCT GTT GAA-3’. The antibody fragments, VH and VL (Variable of the Heavy chain and Variable of the light chain), were inserted into the proper sites of pComb3 and the expression vector was constructed. The insertion of heavy and light chains in the pComb3 vector was proven by double digestion of construct with XhoI (MBI Fermentas Inc, Glen Burnie, MD) and XbaI (MBI Fermentas Inc, Glen Burnie, MD) restriction enzymes.
The digestion was performed overnight at 37°C in the water bath and the digested vector and undigested pComb3 plasmid were run on a 0.8% Agarose gel (Sigma Inc, Poole, UK).
Antibody affinity measurement: ELISA plates were coated with recombinant human leptin receptor (R and D Systems, UK) at 100 ng/well in carbonate buffer and incubated overnight at 4°C. From dot blot data, the concentration of recombinant Fab were estimated compared known cFab (chemically digested of monoclonal antibody by papain enzyme), then different dilutions of rFab and cFab were used in ELISA to optimize the ELISA condition.
Bacterial growth: The glycerol stock of E. coli strains ( XL1-Blue, JM109 and BL21) co-expressing the heavy and light chains of antibody were streaked out onto Luria-Bertani (LB) agar (Himedia Corp Mumbai, India) containing 100 μg mL-1 ampicillin (Sciencelab Inc, Houston, USA) and incubated at 37°C overnight. A single colony was used to inoculate 10 mL Superbroth (Himedia Corp Mumbai, India) (SB) medium that contained 100 μg mL-1 ampicillin.
The inoculums were grown overnight at 37°C with shaking at 250 rpm. These bacterial cultures were then used for the plasmid DNA extraction and production of recombinant antibody.
Transformation of E. coli strain with recombinant pComb3 plasmid: Recombinant pComb3 plasmid containing recombinant antibody sequences were transformed into competent E. coli (Cinnagen Inc, Tehran, Iran) cells by the heat-shock method (Donovan et al., 2000).
Recombinant antibody production: For production of recombinant antibody, four different parameters including: E. coli strains, various culture media, different temperatures and various IPTG (Isopropyl β-D-1-thiogalactopyranoside) concentrations were tested. To express recombinant Fab, a fresh colony of each E. coli cell (XL1-Blue, BL21 or JM109) containing the desired plasmid was inoculated into 5 mL of medium (LB, SB and 2YT) supplemented with 100 μg mL-1 ampicillin and 1% (w/v) glucose (Merk Inc, Whitehouse Station, NJ, USA) and grown at 37°C and shaking at 200 rpm. The overnight culture of bacterial cells was transferred to the large volume of the same medium and grown to mid-logarithmic phase (OD600 nm = 0.6-0.8) at 37°C. The bacterial culture was then centrifuged at 4,000 rpm (~4,700 g) for 15 min to remove the glucose and the cell pellet was resuspended in the equivalent fresh medium containing ampicillin. The protein expression was induced by addition of IPTG (Cinnagen Inc, Tehran, Iran) and incubation at different post-induction temperatures overnight.
Analysis of recombinant Fab (rFab) antibody by dot blotting: Dot blotting was performed in order to detect the protein in a quick way. Basically, the small amounts of culture media containing the rFab (1-2 μL) were applied on the nitrocellulose membrane (Porablot Inc, Germany) and left to dry. The membrane was then blocked by incubating for two hours at room temperature in PBS (Phosphate Buffered Saline) containing 0.05% Tween 20 (Sigma Inc, Poole, UK) (PBS-T) supplemented with 5% w/v skim milk powder (Merk Inc, Whitehouse Station, NJ, USA). After extensive wash with PBS-T, the membrane was probed first with primary antibody (Oncogen, UK) (rat anti-mouse κ light chain, 1:1000) for 1 h and washed again and incubated with secondary antibody (DakoCytomation Inc, Ely-UK), (rabbit anti-rat- HRP, 1:1000) for 1 h and detected by developing in ECL (Amersham Bio-Sciences Corp, NJ, USA) (Enhanced Chemi luminescence) reagents and exposing to X-ray film (Fujifilm, Essex, UK).
Analysis of rFab antibody by SDS-PAGE and Western blotting: The expression of rFab was analyzed using 12% sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) (Donovan et al., 2000).
Plasmid preparation and extraction: Plasmid extraction was performed using alkaline lysis method and BioNeer (Seoul, Korea) DNA plasmid extraction kit (Donovan et al., 2000).
Binding of recombinant Fab to human leptin receptor: The specificity and affinity of recombinant Fab for binding to human leptin receptor was investigated by plate binding assay (Tartaglia et al., 1995).
RESULTS
Confirmation of Fab fragment insertion in the pComb3 vector by double digestion of construct with XhoI and XbaI: The insertion of recombinant Fab in pComb3 plasmid was proven by double digestion of construct with XhoI and XbaI restriction enzymes (Fig. 1). The constructed plasmid DNA containing both heavy and light chains of antibody was transformed into various host strains. The transformed plasmid was then extracted and digested with XbaI and XhoI to confirm the presence of the insert prior to protein expression. The digestion was performed overnight at 37°C and run on a 0.8% agarose gel. Restriction analysis of the construct produced the expected two bands around 3 and 2 kb. The upper band demonstrated linearised pComb3 plasmid and the lower band supported the presence of both heavy chain and light chain fragments in pComb3 vector. The reading frame was restored within the clone when the stuffer fragment was replaced with the rFab gene.
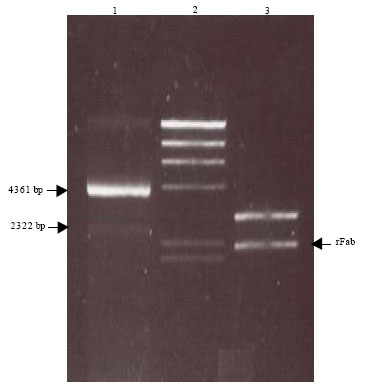 | |
| Fig. 1: | Digestion analysis of pComb3/Fab construct with XhoI and XbaI restriction enzymes: A representative gel photograph of undigested and digested pComb3 plasmid containing DNA sequence for recombinant Fab is shown. Double digestion of construct with XbaI and XhoI enzymes resulted in two fragments at expected size. Lane1: Undigested pComb3 vector, Lane2: Lambda DNA/Hind III marker and Lane3: Double digestion of extracted plasmid DNA from E.coli JM109 |
The cloned Fab fragment was later sequenced and blasted against Kabat system and proved to be a right sequence for mouse IgG1 (data are not shown).
Effect of different culture media on recombinant antibody production: The effect of culture media on recombinant Fab production was investigated by culturing transformed bacteria in different media. Fresh colonies of E. coli JM109 and two other strains containing recombinant plasmid were cultured in either LB medium, 2YT medium or SB medium supplemented with 100 μg mL-1 ampicillin and rFab production was assessed using Dot blot. The highest antibody production was achieved using SB medium while the lowest amount was from LB medium (Fig. 2). Therefore, SB medium was selected for future experiments.
Effect of various E. coli strains on rFab production: As the bacterial strain greatly influences the protein production, three different E. coli strains were tested for their ability to produce rFab. Three strains of E. coli (XL1-Blue, BL21 and JM109) were transformed and the protein production was inducted. Subsequently, the supernatant were analyzed by Dot blot. The E. coli JM109 strain demonstrated the highest production rate of rFab compared to other two strains (Fig. 3). Therefore, E. coli JM109 was selected for further experiments.
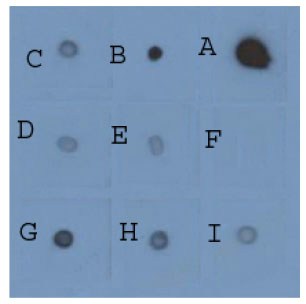 | |
| Fig. 2: | Dot blot analysis demonstrating the expression level of recombinant Fab in various media: Culture supernatant were applied onto nitrocellulose membrane, blocked by skim milk, probed with antibodies and detected by developing in ECL reagents and exposed to X-ray film. A to E: Positive control dilution series (Fab created by papain cleavage, 30 mg mL-1) from 1 to 0/0001 dilutions. F: Negative control (Growth hormone, 1 μg). G: SB medium (showing around 0.03 mg mL-1 of rFab compared to positive control). H: 2YT medium (showing around 0.003 mg mL-1 of rFab compared to positive control). I: LB medium (showing around 0.0003 mg mL-1 of rFab compared to positive control). All experiments were done at least three times and in triplicates |
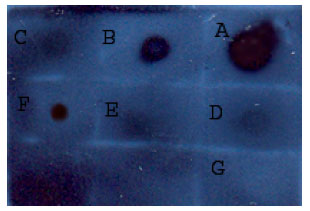 | |
| Fig. 3: | Dot blot analysis showing the rFab expression level by different E. coli strains: Supernatant (2 μL) from transformed bacterial culture were applied onto nitrocellulose membrane, blocked in skim milk, probed with antibodies and detected by developing in ECL reagents and exposing to X-ray film. A to C: Positive control dilution series (Fab created by papain cleavage, 30 mg mL-1 stock) dilutions from 0.01 to 0/0001. D: Supernatant of E. coli XL1-Blue. E: Supernatant of E. coli BL21, F: Supernatant of E. coli JM109.G: Negative control (Growth hormone, 1 μg) |
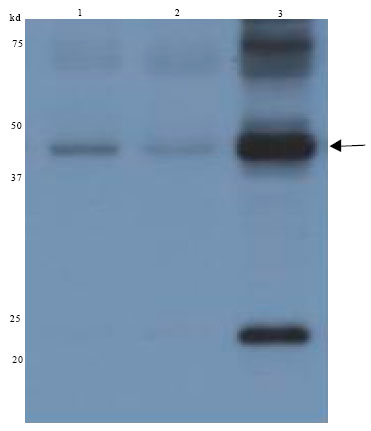 | |
| Fig. 4: | Western blot analysis showing the rFab expression level of different E. coli strains: Supernatant (10 μL) from transformed bacteria were run on a 12% SDS-PAGE and transferred onto nitrocellulose membrane, blocked by skim milk, probed with antibodies and detected by developing in ECL reagents and exposing to X-ray film. Lane 1: rFab expression by E. coli XL1-Blue strain, Lane 2: rFab expression by E.coli BL21 strain and Lane 3: rFab expression by E. coli JM109 strain. Arrow indicates the rFab |
To assess the size of recombinant Fab produced by bacteria and also to confirm the results of Dot blot, Western blot analysis were also performed (Fig. 4). As demonstrated, E. coli JM109 showed the highest rFab production compared to other strains. A band around 50Kd could be detected on Western blotting which was expected size for rFab.
Impact of various pos-induction temperatures on level of rFab antibody production: The effect of different culturing temperatures on the expression level of rFab was investigated. The transformed E. coli JM109 expressing the heavy and light chain of rFab was cultured in SB medium containing 100 μg mL-1 ampicillin in different temperatures (22 to 37°C) following IPTG induction. Dot blot data analysis revealed that the antibody was produced at both 22 and 30°C with higher production rate at 30°C but there was no detectable amount of antibody form bacteria incubated at 37°C (Fig. 5). Therefore, for further experiments the 22°C culture temperature was chosen.
Effect of various IPTG concentrations on rFab production: The influence of IPTG concentration on recombinant Fab production was evaluated using different concentrations (0.75 to 1.5 mM) of IPTG.
 | |
| Fig. 5: | Dot blot analysis demonstrating the expression level of recombinant Fab at various post-induction temperatures: Supernatant (2 μL) from media were applied onto nitrocellulose membrane, blocked by skim milk, probed with antibodies and detected by developing in ECL reagents and exposing to X-ray film. A to E: Positive control dilution series (Fab created by papain cleavage, 30 mg mL-1 stock) from 1 to 0/0001 dilutions respectively. F: Negative control (Growth hormone, 1 μg). G: Medium from E. coli JM109 cultured at either 22°C (G), 30°C (H) or 37°C (I) |
 | |
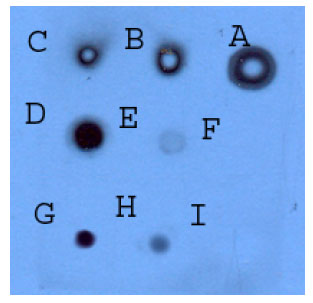
| Fig. 6: | Dot blot analysis illustrating the expression level of recombinant Fab during various post induction temperatures: Supernatant (2 μL) from media were applied onto nitrocellulose membrane, blocked by skim milk, probed with antibodies and detected by developing in ECL reagents and exposing to X-ray film. Lane A to D: Positive control dilution series (Fab created by papain cleavage, 30 mg mL-1) from 0.1 to 0/0001 dilution. Lane E: rFab production in 0.75 mM of IPTG. Lane F: rFab production in 1 mM of IPTG. Lane G: rFab production in 1.5 mM of IPTG |
The competent E. coli JM109 expressing the heavy and light chain of rFab were cultured in SB medium containing 100 μg mL-1 ampicillin followed by our standard protein production procedure in which the protein expression was induced by addition of three different IPTG concentrations (0.75, 1 and 1.5 mM). Recombinant Fab production was assessed by Dot blot technique. The level of rFab production was higher in high concentrations of IPTG without any significant difference between 1 and 1.5 mM. The level of protein production was low at 0.75 mM IPTG and therefore, 1 mM concentration of IPTG was used for further experiments (Fig. 6).
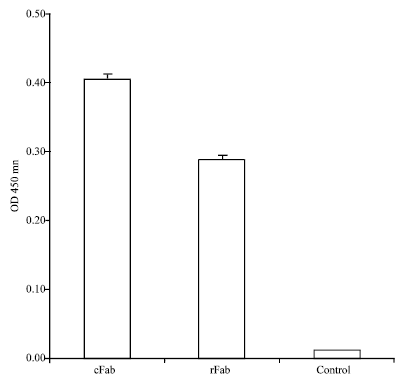 | |
| Fig. 7: | Antigen binding of recombinant Fab: The antigen binding of rFab was assessed by plate binding assay. The ninety six-well plate was coated with human leptin receptor, blocked with PBS-BSA, incubated with different samples and antigen-binding was detected using secondary antibody and TMB substrate. cFab: papain made Fab from original mouse monoclonal antibody, rFab: Recombinant Fab and Control: Supernatant from pComb3 empty vector-transformed JM109 |
Binding of recombinant 9F8 Fab to human leptin receptor: The binding of recombinant Fab to human leptin receptor was investigated by ELISA assay. The recombinant Fab molecule recognised human leptin receptor as demonstrated by plate binding assay (Fig. 7). The results showed that both the recombinant immunoglobulin fragment and the enzymatically made Fab by papain bound to human leptin receptor while no reaction could be observed with the supernatant from negative control (pComb3 empty vector transformed bacteria). Hence, the recombinant Fab was specific to human leptin receptor and showed an affinity comparable to that of enzymatically Fab produced from original hybridoma antibody (Fig. 7).
DISCUSSION
The Fab fragment is a small molecule compared to the whole molecule of monoclonal antibody which usually has the same affinity for antigen binding and bioactivity (Burton et al., 1991). The Fab fragment exerts many advantages to intact antibody including low immunogenicity, easy manufacturing, cost effectiveness and improved pharmacokinetics properties (Donovan et al., 2000). Although Fab fragment can be produced by enzymatic cleavage, recombinant Fab is preferred as it is homogenous and can be subjected to humanization and manipulation which facilitate minimizing its immunogenicity and improving its biological activities. More importantly, the large-scale bacterial expression of rFab enables this protein to be produced in quantities that might be difficult to obtain from natural sources (Swartz, 2001). Our initial attempts to express rFab in E. coli XL1-Blue resulted in a very low level of protein expression. Therefore, to obtain higher yield of antibody, bacterial expression level of rFab was optimised by examining a variety of media, different strains of E. coli, various post-induction temperature and varied IPTG concentrations. The maximal expression level of rFab was achieved with the IPTG-induced JM109 strain of E. coli (at 1 mM IPTG) and the optimal expression conditions were identified as pre-induction glucose addition and growing culture in the Super Broth (SB) medium at 22°C for overnight.
In this study, the pComb3 expression vector was used to clone the heavy and light chains of antibody. The pComb3 vector is a common expression plasmid for Fab cloning since it allows co-expression of both chains and is appropriate for bacterial protein expression. After deletion of pIII fragment, it is possible to produce recombinant antibody in a soluble form. This plasmid has two pelB sequences for direction of antibody to periplasmic space and two lacZ sequences for lac promoter providing (Barbas et al., 1991). This allows each heavy and light chain of recombinant antibody to be produced separately, then assembled in periplasmic space to form the Fab antibody. Many scientists used this vector for antibody production with good results which were comparable to present results. The expression of the genes cloned downstream of the lac promoter can be conveniently induced by the addition of IPTG (De Bellis and Schwartz, 1990). All of the host cells that were used have the lac Iq genotype to limit the background expression from the lac promoter. This also ensures that the expression only occurs upon the induction by IPTG as constitutive expression which would almost certainly be detrimental and debilitating to the host (Carrier et al., 1993). Overproduction of foreign proteins that are encoded within the DNA that is introduced into the host cell will trigger the metabolic burden phenomenon and cause the cells to enter the stationary phase and finally leads to cell death; therefore, the IPTG concentration that was used in this study was limited to 1 mM which is comparable to other findings by several researchers (Zwick et al., 2001). It was reported that IPTG could increase the permeability of outer membrane; hence, it leads to realising about 90% of periplasmic proteins to extracellular medium (Markland, 2006). Many factors can affect the efficiency of the expression (e.g., choice of nutrients and environmental parameters, such as temperature, dissolved oxygen tension, etc.). The expression of foreign proteins in a recombinant host cell often utilizes significant amounts of host cell resources which places a metabolic burden on the host. It leads to a decrease in growth and a reduction of the protein expression (Maldonado et al., 2007). Therefore, to enhance the protein expression in this study, a concentration of 1% glucose (w/v) was added to the growth medium. This strategy was widely used among researchers and also was used in this study which was lead to good result as compared to other researchers (Donovan et al., 2000). In the absence of the inducing agent (IPTG), the Fab proteins were detected only in the culture medium of all three E. coli strains cells that carried recombinant pComb3 (data not shown). This implied that in the absence of an inducer and in the presence of the lac repressor, there was a basal level expression from the lac promoter (Dubendorff and Studier, 1991). The Fab protein in the extracellular medium was detected in all three strains, as a band of around 50 kDa (Fig. 4). This was a result of the secretion of soluble Fab from the periplasm into the extracellular growth medium which contained pelB sequences that directed each heavy and light chain to periplasmic space. The Fab expression of cloned anti-leptin receptor in all of the host strains was optimal in the growth medium that contained an additional carbon source (i.e., glucose or glycerol). Therefore, there is a need to supply an additional carbon source for the enhanced protein expression. The addition of 1% glucose before induction dramatically increased the yield and the solubility of the rFab leading to the presence of protein in the conditioned medium by leaking from periplasmic space to the medium. The presence of glucose in the growth medium suppresses expression from lacZ promoter, thereby eliminating any deleterious background protein expression. The addition of IPTG following glucose removal will result in induction of expression from lacZ promoter. However, in the absence of inducer, there was a basal level of expression which could be due to insufficiency of glucose to repress the promoter (data not shown). The preinduction glucose addition suppresses the lac promoter of vector and allows the bacterial cells to be proliferated without Fab expression. It was reported that when the glucose concentrations increased above 1%, the optimal expression was reduced. This could be due to the over-feeding of glucose to the host cells which enhanced glucose metabolism and consequently led to the fast accumulation of such metabolites which are detrimental to the host cell (Robbins and Taylor, 1989).
One of the most important factors that affect protein production by bacteria is culture medium. Preliminary tests clearly showed that the yield of protein was higher in SB than in LB medium. LB is not buffered and lacks enough carbon sources. Consequently, the pH of the medium decreases quickly and causes slow growth rate and poor expression level. MOPS is one of the most effective ingredient in SB medium which causes the secretion of periplasmic protein into the extracellular culture. Several investigators applied SB medium rather than LB or 2YT medium, therefore, their results are comparable to these findings (Markland, 2006; Zwick et al., 2001). The one known technique for preventing of inclusion body is culturing at the low temperature. This strategy effectively improves the soluble antibody expression. Low temperature also facilitates proper folding by preventing overproduction of protein and results in expression of more soluble protein (Schumann and Ferreira, 2004). In contrast, the formation of inclusion body at the high temperature is more favourable. In addition, high temperature increases the growth of bacteria and the yield of protein but reduces proper folding and the solubility of produced protein leads to protein aggregation and formation of insoluble inclusion bodies. Another benefit of using low temperature is partial deletion of heat shock proteases which are induced at the extreme conditions and degrade the proteins (Schumann and Ferreira, 2004).
Therefore, among the several temperatures (22 to 37°C) used for post-induction incubation, 22°C was found to be the best for soluble antibody production in extracellular medium, hence these findings are strongly supported by other researchers findings, except for those who desired the high level of antibody fragment as an inclusion body (Burton et al., 1991).
CONCLUSION
In conclusion, we reported cloning and optimized conditions for production of a recombinant antibody against human leptin receptor which binds to its ligand with comparable efficiency to original monoclonal antibody produced by mouse hybridoma cells. This recombinant antibody will be subjected to genetic manipulation for further studies and possible clinical pharmaceutical applications.
ACKNOWLEDGMENT
This study was supported by Research Office and Graduate Studies Office of University of Isfahan, Isfahan, Iran.
REFERENCES
- Barbas, 3rd, C.F., A.S. Kang, R.A. Lerner and S.J. Benkovic, 1991. Assembly of combinatorial antibody libraries on phage surfaces: The gene III site. Proc. Natl. Acad. Sci., 88: 7978-7988.
Direct Link - Barr, V.A., K. Lane and S.I. Taylor, 1999. Subcellular localization and internalization of the four human leptin receptor isoforms. J. Biol. Chem., 274: 21416-21424.
CrossRef - Burton, D.R., C.F. Barbas 3rd, M.A. Persson, S. Koenig, R.M. Chanock and R.A Lerner, 1991. A large array of human monoclonal antibodies to type 1 human immunodeficiency virus from combinatorial libraries of asymptomatic seropositive individuals. Proc. Natl. Acad. Sci., 88: 10134-10137.
Direct Link - Busso, N., A. So, V. Chobaz-Peclat, C. Morard, E. Martinez-Soria, D. Talabot-Ayer and C. Gabay, 2002. Leptin signaling deficiency impairs humoral and cellular immune responses and attenuates experimental arthritis. J. Immunol., 168: 875-882.
Direct Link - Carrier, M.J., M.E. Nugent, W.C.A. Tacon and S.B. Primrose, 1993. High expression of cloned genes in E. coli and its consequences. Trends Biotechnol., 1: 109-113.
CrossRef - Choi, J.H. and S.Y. Lee, 2004. Secretory and extracellular production of recombinant proteins using Escherichia coli. Applied Microbiol. Biotechnol., 64: 625-635.
CrossRef - Correia, M.L. and W.G. Haynes, 2004. Leptin, obesity and cardiovascular disease. Curr. Opin. Nephrol. Hypertens., 13: 215-223.
PubMed - De Bellis, D. and I. Schwartz, 1990. Regulated expression of foreign genes fused to lac: Control by glucose levels in growth medium. Nucleic Acids Res., 18: 1311-1316.
Direct Link - Donovan, R.S., C.W. Robinson and B.R. Glick, 2000. Optimizing the expression of a monoclonal antibody fragment under the transcriptional control of the Escherichia coli lac promoter. Can. J. Microbiol., 46: 532-541.
CrossRef - Dubendorff, J.W. and F.W. Studier, 1991. Controlling basal expression in an inducible T7 expression system by blocking the target T7 promoter with lac repressor. J. Mol. Biol., 219: 45-59.
CrossRef - Farooqi, I.S., S.A. Jebb, G. Langmack, E. Lawrence and C.H. Cheetham et al., 1999. Effects of recombinant leptin therapy in a child with congenital leptin deficiency. N. Engl. J. Med., 12: 879-884.
PubMed - Farooqi, I.S., G. Matarese, G.M. Lord, J.M. Keogh and E. Lawrence et al., 2002. Beneficial effects of leptin on obesity, T cell hyporesponsiveness and neuroendocrine/metabolic dysfunction of human congenital leptin deficiency. J. Clin. Invest., 110: 1093-1103.
PubMedDirect Link - Friedman, J.M. and J.L. Halaas, 1998. Leptin and the regulation of body weight in mammals. Nature, 395: 763-770.
CrossRefDirect Link - Ghilardi, N., S. Ziegler, A. Wiestner, R. Stoffel, M.H. Heim and R.C. Skoda, 1996. Defective STAT signaling by the leptin receptor in diabetic mice. Proc. Natl. Acad. Sci., 93: 6231-6235.
Direct Link - Iversen, P.O., C.A. Drevon and J.E. Reseland, 2002. Prevention of leptin binding to its receptor suppresses rat leukemic cell growth by inhibiting angiogenesis. Blood, 100: 4132-4138.
Direct Link - Landman, R.E., J.J. Puder, E. Xiao, P.U. Freda, M. Ferin and S.L. Wardlaw, 2003. Endotoxin stimulates leptin in the human and nonhuman primate. J. Clin. Endocrinol. Methods., 88: 1285-1291.
CrossRef - Li, C. and J.M. Friedman, 1999. Leptin receptor activation of SH2 domain containing protein tyrosine phosphatase 2 modulates Ob receptor signal transduction. Proc. Natl. Acad. Sci., 96: 9677-9682.
Direct Link - Loffreda, S., S.Q. Yang, H.Z. Lin, C.L. Karp and M.L. Brengman et al., 1998. Leptin regulates proinflammatory immune responses. FASEB J., 12: 57-65.
PubMedDirect Link - Lord, G.M., G. Matarese, J.K. Howard, R.J. Baker, S.R. Bloom and R.I. Lechler, 1998. Leptin modulates the T-cell immune response and reverses starvation-induced immunosuppression. Nature, 394: 897-901.
PubMed - Maffei, M., H. Fei, G.H. Lee, C. Dani and P. Leroy et al., 1995. Increased expression in adipocytes of ob RNA in mice with lesions of the hypothalamus and with mutations at the db locus. Proc. Natl. Acad. Sci., 92: 6957-6960.
Direct Link - Maldonado, L.M.T.P., V.E.B. Hernandez, E.M. Rivero, A.P. Barba de la Rosa, J.L.F. Flores, L.G.O. Acevedo and A. De Leon Rodriguez, 2007. Optimization of culture conditions for a synthetic gene expression in Escherichia coli using response surface methodology: The case of human interferon beta. Biomol. Eng., 24: 217-222.
CrossRef - Matarese, G., S. Moschos and C.S. Mantzoros, 2005. Leptin in immunology. J. Immunol., 174: 3137-3142.
Direct Link - Matarese, G., V. Sanna, A. Di Giacomo, G.M. Lord and J.K. Howard et al., 2001. Leptin potentiates experimental autoimmune encephalomyelitis in SJL female mice and confers susceptibility to males. Eur. J. Immunol., 31: 1324-1332.
PubMed - Matarese, G., V. Sanna, R.I. Lechler, N. Sarvetnick, S. Fontana, S. Zappacosta and A. La Cava, 2002. Leptin accelerates autoimmune diabetes in female NOD mice. Diabetes, 51: 1356-1361.
CrossRef - Mergulhao, F.J.M., D.K. Summers and G.A. Monteiro, 2005. Recombinant protein secretion in Escherichia coli. Biotechnol. Adv., 23: 177-202.
CrossRef - Pardridge, W.M., 1986. Receptor-mediated peptide transport through the blood-brain barrier. Endocrinol. Rev., 7: 314-330.
CrossRef - Robbins, Jr. W.J. and K.B. Taylor, 1989. Optimization of Escherichia coli growth by controlled addition of glucose. Biotechnol. Bioeng., 34: 1289-1294.
CrossRef - Saad, M.F., S. Damani, R.L. Gingerich, M.G. Riad-Gabriel and A. Khan et al., 1997. Sexual dimorphism in plasma leptin concentration. J. Clin. Endocrinol. Metab., 82: 579-584.
CrossRefDirect Link - Sanna, V., A. Di Giacomo, A. La Cava, R.I. Lechler, S. Fontana, S. Zappacosta and G. Matarese, 2003. Leptin surge precedes onset of autoimmune encephalomyelitis and correlates with development of pathogenic T cell responses. J. Clin. Invest., 111: 241-250.
CrossRef - Schumann, W. and L.C.S. Ferreira, 2004. Production of recombinant proteins in Escherichia coli. Genet. Mol. Biol., 27: 442-453.
CrossRef - Shiloach, J. and R. Fass, 2005. Growing E. coli to a high cell density-a historical perspective on method development. Biotechnol. Adv., 23: 345-357.
CrossRef - Smith, J.T. and B.J. Waddell, 2003. Developmental changes in plasma leptin and hypothalamic leptin receptor expression in the rat: Peripubertal changes and the emergence of sex differences. J. Endocrinol., 176: 313-319.
CrossRef - Swartz, J.R., 2001. Advances in Escherichia coli production of therapeutic proteins. Curr. Opin. Biotechnol., 12: 195-201.
CrossRef - Tartaglia, L.A., M. Dembski, X. Weng, N. Deng and J. Culpepper et al., 1995. Identification and expression cloning of a leptin receptor, OB-R. Cell, 83: 1263-1271.
CrossRefPubMedDirect Link - Terpe, K., 2006. Overview of bacterial expression systems for heterologous protein production: From molecular and biochemical fundamentals to commercial systems. Appl. Microbiol. Biotechnol., 72: 211-222.
PubMedDirect Link - Zarkesh-Esfahani, H., A.G. Pockley, Z. Wu, P.G. Hellewell, A.P. Weetman and R.J. Ross, 2004. Leptin indirectly activates human neutrophils via induction of TNF-alpha. J. Immunol., 172: 1809-1814.
Direct Link - Zwick, M.B., A.F. Labrijn, M. Wang, C. Spenlehauer and E.O. Saphire et al., 2001. Broadly neutralizing antibodies targeted to the membrane-proximal external region of human immunodeficiency virus type 1 glycoprotein gp41. J. Virol., 75: 10892-10905.
PubMedDirect Link