
S. Gurudeeban
Faculty of Marine Sciences, Annamalai University, Parangipettai 608502, India
T. Ramanathan
Faculty of Marine Sciences, Annamalai University, Parangipettai 608502, India
K. Satyavani
Faculty of Marine Sciences, Annamalai University, Parangipettai 608502, India
T. Dhinesh
Faculty of Marine Sciences, Annamalai University, Parangipettai 608502, India
Asian Journal of Biotechnology
Year: 2011 | Volume: 3 | Issue: 5 | Page No.: 486-492







ABSTRACT
The present study has evaluated the genetic variability among three salt marsh plant belongs to the family of Chenopodiaceae using random amplified polymorphic DNA markers. The method involves a modified Cetyl trimethylammonium bromide extraction, using polyvinyl pyrrolidone while grinding, successive long-term Chloroform: lsoamyalcohol extractions, an overnight RNase treatment with all steps carried out at room temperature. The yield of DNA ranged from 1.5-2.5 μg μL-1 per gram of the leaf tissue. The technique is ideal for isolation of DNA from different salt marsh plant species and used for Randomly Amplified Polymorphic DNA (RAPD) analysis. RAPD protocol was optimized based on the use of lower concentrations of primer (2 μM) and Taq polymerase (2 units), 50 ng of template DNA, higher concentration of MgCl2 (2 mM) and an annealing temperature of 37°C, resulted optimal amplification. The results suggest that the optimized protocol for DNA isolation and PCR was appropriate to diversity analysis of salt marsh plants.
PDF Abstract XML References Citation
Received: February 03, 2011;
Accepted: May 02, 2011;
Published: July 09, 2011
How to cite this article
S. Gurudeeban, T. Ramanathan, K. Satyavani and T. Dhinesh, 2011. Standardization of DNA Isolation and PCR Protocol for RAPD Analysis of Suaeda sp. Asian Journal of Biotechnology, 3: 486-492.
DOI: 10.3923/ajbkr.2011.486.492
URL: https://scialert.net/abstract/?doi=ajbkr.2011.486.492
DOI: 10.3923/ajbkr.2011.486.492
URL: https://scialert.net/abstract/?doi=ajbkr.2011.486.492
INTRODUCTION
Salt marshes are widely distributed in estuarine systems around the world (Nicol, 2007). Chenopodiaceae is one of the most interesting families with respect to having species with a large diversity in the structure of the carbon assimilating organs with different types of photosynthesis (Grigore and Toma, 2007). The high salinity of soils and the soil’s moisture had a major impact on halophytes anatomical structures and has formative effects. Their cumulated action had accompanied the halophytes evolution through time, as an active and dynamic component of the evolutionary “adventure” (Grigore and Toma, 2005). Suaeda monoica, Suaeda maritima and Suaeda nudiflora are salt marsh mangrove herbs, growing in hyper saline soils and distributed throughout the East west coast mangroves in India viz., Sunderbans (West Bengal), Bitharkanika and Mahnadhi (Orissa), Coringa, Godavari and Krishna (Andhra Pradesh), Pichavaram, Karangadu and Muthupet (Tamil Nadu) (Ramanathan et al., 2011).
Salt marsh plants contain exceptionally high amounts of polyphenols, tannins and secondary metabolites such as alkaloids, flavanoids and phenols which would interfere with the DNA isolation procedures (Bandaranayake, 2002). The problems encountered in the isolation and purification of DNA specially from halophytes include degradation of DNA due to endonucleases, inhibitor compounds like polyphenols and secondary metabolites which directly or indirectly interfere with the enzymatic reactions. Moreover, the contaminating RNA that precipitates along with DNA causes many problems in aromatic plants includes suppression of PCR amplification, interference with DNA amplification involving random primers and improper priming of DNA templates during thermal cycle sequencing (Padmalatha and Prasad, 2006). The surviving brown marsh (Spartina alterniflora) individuals are genetically different from those of healthy marshes and their survival could be due to favorable combination of genes responsible for tolerance to multiple abiotic stresses (Subudhi et al., 2008). Also the genetic variation affects population viability by providing the raw material for natural selection during periods of environmental change (Hamrick, 2004).
The genetic diversity and molecular identification by using RAPD markers had been performed in several plant species (Bautista et al., 2003). These molecular markers provide an opportunity for direct comparison and identification of different genetic material independent of any influences (Zhao and Pan, 2004). Therefore, we investigated the present work on total genomic DNA isolation protocol derived from a method originally developed for plants (Doyle and Doyle, 1987). The purpose of the present study is made to minimize and simplify the DNA isolation procedure and to assess genetic diversity within and among the species using RAPD markers.
MATERIALS AND METHODS
Plant material: Three plant species viz., Suaeda monoica, Suaeda maritima and Suaeda nudiflora were freshly collected from Southeast coast of India (Pichavaram Mangrove forest; Tamil Nadu) during the month of November 2010. The plants were taxonomically identified and authenticated in the Herbaria being maintained at the Centre of Advanced Study in Marine Biology, Annamalai University and (Voucher No. AUCASMB15, AUCASMB16, AUCASMB17).
Genomic DNA isolation protocol: Freshly collected leaf sample (1 g) was ground using a mortar and pestle along with 0.1 g of Poly Vinyl Pyrrolidone (PVP). The grinded leaves were quickly transferred to 3 mL of freshly prepared prewarmed (65°C) extraction buffer and shaken vigorously by in- version to form slurry. The tubes were incubated at 65°C in hot air oven with intermittent shaking and swirling for every 30 min. An equal volume of Chloroform: Isoamylalcohol (24:1) was added and mixed properly by inversion for 30 min and centrifuged at 12,000 rpm for 15 min at room temperature to separate the phases. The supernatant was carefully decanted and transferred to a new tube and was precipitated with equal volumes of cold lsopropanol and gently mixed to produce fibrous DNA and incubated at -20°C. The samples were centrifuged at 12000 rpm for 15 min. The pellet was washed with 70% ethanol, air dried and resuspended in 3 mL of TE buffer and 5 μL of RNase was added and incubated overnight at 37°C. The dissolved DNA was extracted with equal volumes of phenol: chloroform: lsoamylalcohol (25:24:1, v/v/v) at 8000 rpm for 15 min. The aqueous layer was transferred to a fresh 15 mL tube and re extracted with equal volume of chloroform and lsoamyalcohol (24:1) by centrifuging at 12,000 rpm for 15 min. The supernatant was transferred to a fresh tube and equal volumes of absolute alcohol and 1/10 volume of sodium acetate were added and incubated at -20°C for 30 min followed by centrifugation at 12,000 rpm for 15 min. The pellet was air dried and resuspended in TE buffer. All the centrifugation steps were carried out at RT to avoid precipitation with CTAB, DNA degradation and to obtain good quality DNA.
Concentration of isolated DNA: The yield of DNA per gram of leaf tissue extracted was measured using a UV Spectrophotometer (PerkinElmer, USA) at 260 nm. The purity of DNA was determined by calculating the ratio of absorbance at 260 nm to that of 280 nm. DNA concentration and purity was also determined by running the samples on 0.8% agarose gel based on the intensities of band when compared with the Lambda DNA marker.
Random amplified polymorphic DNA reaction: For the optimization of RAPD reaction using DNA extracted from Suaeda sp., oligonucleotide primers from A and C series (Operon Technologies Inc., USA) were used for amplification to standardize the PCR conditions. The polymerase chain reactions were carried out in a Thermo cycler (Lark, India). Reactions without DNA were used as negative controls. Each 15 μL reaction volume contained about 50 ng of template DNA, 1X PCR Buffer (10 mM Tris HCl pH 8; 50 mM KCl), 3 mM MgCl2 (Sigma, USA), 0.2 mM dNTP Mix, 0.5 μM of single primer, 0.2 U of Taq DNA polymerase (Helini Biomolecules, India). The thermo cycler was programmed for an initial denaturation step of 3 min at 94°C, followed by 30 cycles at 94°C, 1 min at 37°C, extension was carried out at 72°C for 1 min and final extension at 72°C for 5 min and a hold temperature of 4°C at the end. PCR products were electrophoresed on 2% (w/v) agarose gels, in 1X TBE Buffer at 50 V for 3 h and then stained with ethidiumbromide (0.5 μg mL-1). Gels with amplification fragments were visualized and photographed under UV light. Lambda DNA and Hind III digest was used as molecular marker (Helini Biomolecules, India) to know the size of the fragments.
RESULTS AND DISCUSSION
DNA extraction was improved by modifying some of the steps in the original CTAB DNA isolation protocol. Presented procedure resulted in extracting, high quality, low-polysaccharide genomic DNA from three different salt marsh plant species belonging to Chenopodiaceae. We found following modified steps necessary to standardize and increase the quality and quantity of genomic DNA such as concentration of DNA 50 ng, Magnesium chloride (mM) dNTPs 2 mM, deoxynucleotide triphosphates 10 mM, primer concentration 2 μM, Taq polymerase 2 U, initial denaturation time 94°C for 3.0 min, annealing temperature 37°C for 60 sec and reaction volume 15 μL. The degree of purity and quantity varies between applications (Zidani et al., 2005). Similar observation had been made important Nigerian fiber plant Corchorus sp. (Ogunkanmi et al., 2010). The yield of extracted DNA with high quality was obtained ranged from 1.5 to 2.5 μg μL-1.
The study plants contain exceptionally high amounts of polyphenols, tannins and secondary metabolites such as alkaloids, flavanoids and phenols which would interfere with the DNA isolation procedures (Bandaranayake, 2002). The presence of polyphenols which are powerful oxidizing agents present in many plant species, can reduce the yield and purity by binding covalently with the extracted DNA making it useless for most research applications (Singh et al., 2010). Tannins, terpenes and resins considered as secondary metabolites are also difficult to separate from DNA (Mazid et al., 2011). Certain polysaccharides are known to inhibit RAPD reactions. They distort the results in many analytical applications and therefore lead to wrong interpretations (Kotchoni et al., 2003). The plants with higher secondary metabolites using CTAB/NaCl the polysaccharides are precipitated and elimination of proteins through chloroform: isoamyl alcohol (Dehestani and Kazemi Tabar, 2007). Polysaccharide co-precipitation is avoided by adding a selective precipitant of nucleic acids, to keep polysaccharides in solution though SDS (Dellaporta et al., 1983), sorbitol and glucose were added initially in the extraction buffer which did not exhibit any effect. Addition of PVP along with CTAB may bind to the polyphenolic compounds by forming a complex with hydrogen bonds and may help in removal of impurities to some extent. Also in addition of Poly Vinyl Pyrrolidone (PVP) in CTAB protocol, increased the solubility of polysaccharides present in woody plants (Sahasrabudhe and Deodhar, 2010). Long-term chloroform: isoamylalcohol treatment ensured removal of chlorophyll, pigments and dyes. Many DNA isolation procedures also yield large amounts of RNA, especially 18S and 25S rRNA (Joshi et al., 2010). Large amounts of RNA in the sample can chelate Mg2+ and reduce the yield of the PCR. A prolonged overnight RNase treatment degraded RNA into small ribonucleosides that do not contaminate the DNA preparation and yielded RNA-free pure DNA. Additional precipitation steps removed large amounts of precipitates by centrifugation and modified speed and time.
DNA isolated by this method yielded strong and reliable amplification products showing its compatibility for RAPD-PCR using random decamer primers (Fig. 1). In all 20 random primers were tested for amplification of which 12 random primers showed amplification in all the species and 8 primers did not give any amplification. Out of selected 12 primers only 4 primers produced polymorphic bands. Total numbers of polymorphic bands were 20. The size of amplicons was in the range of 4500 to 1800 bp. The highest number of polymorphic bands (11) was obtained with OPD-18 while the lowest number (2) was obtained with primer OPA-01. Different primers showed variation in their ability to detect polymorphism. The RAPD banding pattern varied with salt marsh plants. In OPD 18 (GAGAGCCAAC), a total of 4 alleles were observed. The alleles sized ranging from 1799 to 4513 bp (1799, 3328, 3925 and 4513 bp, respectively); In OPD 20 (ACCCGGTCAC), a total of 3 alleles were observed. The alleles sized from 3227, 3847 and 4782 bp, respectively. In OPA 04 (AATCGGGCTG), a total of 3 alleles were observed. The alleles sized from 1740, 3135 and 4905 bp. In OPA 01(CAGGCCCTTC), a total of 2 alleles were observed. The alleles sized ranges between 2000 and 4615 bp. The electrophoretic pattern of the entire four loci is shown (Fig. 2). Almost all the tested parameters for RAPDs like the concentration of template DNA, primer, magnesium chloride, Taq polymerase, dNTPs and temperature and time intervals during denaturation, annealing and elongation were also optimized which also had an effect on amplification, banding patterns and reproducibility. The optimized conditions for RAPD protocol are given in Table 1. The size of the amplified fragments ranged from 5000-2000 bp.
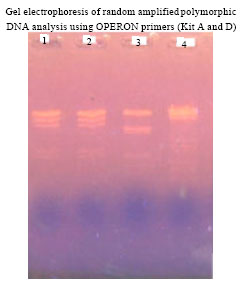 | |
| Fig. 1: | Gel fragmentation of RAPD-PCR amplification product of plant genomic DNA 50 ng. Lane 1-3 represents RAPD-PCR products amplified from genomic DNA of study plants. Lane 4 (1kb) Molecular Marker |
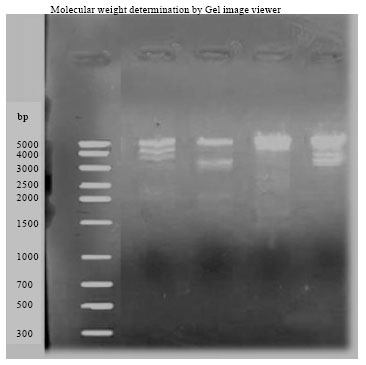 | |
| Fig. 2: | RAPD banding pattern varied with salt marsh plants using OPERON primers. The size of amplicons was in the range of 4500 to 1800 bp |
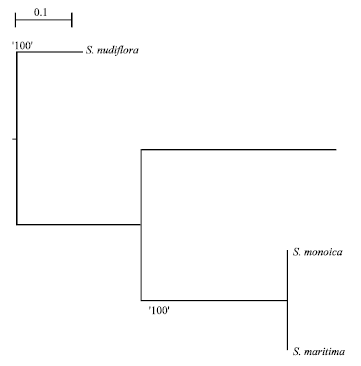 | |
| Fig. 3: | Dendogram analysis showing phylogenetic relationship of study plants identified by RAPD markers using Freetree soft ware |
| Table 1: | Genetic distance between three Suaeda species |
 | |
| Note: *Indicates no genetic variation | |
RAPD markers are used to determine which genetic resources should be retained in a collection in order to conserve maximum genetic diversity in the gene bank (Al-Saghir and Abdel Salam, 2011).
Genetic variability: Based on the 11 random primers, the genetic distance was calculated and displayed in (Table 1). The genetic distance between Suaeda maritima, Suaeda monoica and Suaeda nudiflora was calculated by using the software Freetree (URL:www.natur.cuni.cz/~flegr/programs/freetree.htm).The genetic variation between Suaeda maritima and Suaeda monoica was 0.42, whereas the genetic distance was much more 0.66 between Suaeda maritima and Suaeda nudiflora. The dendogram of phylogenetic relationship of salt marsh plants identified by RAPD markers showed in Fig. 3.
CONCLUSION
The optimized DNA isolation protocol and RAPD markers provided an efficient tool for molecular studies of salt marsh plants.
ACKNOWLEDGMENT
The authors are gratefully acknowledge to the Dean, Faculty of Marine Sciences, Annamalai University, Parangipettai, Tamil Nadu, India for providing all support during the study period.
REFERENCES
- Al-Saghir, M.G. and A.G. Abdel Salam, 2011. Genetic diversity of North American Soybean (Glycine max L.) cultivars as revealed by RAPD markers. J. Plant Sci., 6: 36-42.
CrossRef - Bandaranayake, W.M., 2002. Bioactivities, bioactive compounds and chemical constituents of mangrove plants. Wetlands Ecol. Manage., 10: 421-452.
CrossRefDirect Link - Bautista, R., F.M. Canovas and M.G. Claros, 2003. Genomic evidence for a repetitive nature of the RAPD polymorphisms in Olea europaea (olive-tree). Euphytica, 130: 185-190.
CrossRefDirect Link - Dellaporta, S.L., J. Wood and J.B. Hicks, 1983. A plant DNA minipreparation: Version II. Plant Mol. Biol. Rep., 1: 19-21.
CrossRefDirect Link - Dehestani, A. and S.K.K. Tabar, 2007. A rapid efficient method for DNA isolation from plants with high levels of secondary metabolites. Asian J. Plant Sci., 6: 977-981.
CrossRefDirect Link - Doyle, J.J. and J.L. Doyle, 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull., 19: 11-15.
Direct Link - Grigore, M.N. and C. Toma, 2007. Histo-anatomical strategies of Chenopodiaceae halophytes: adaptive, ecological and evolutionary implications. WSEAS Trans. Biol. Biomed., 12: 204-218.
Direct Link - Hamrick, J.L., 2004. Response of forest trees to global environmental change. For. Ecol. Manage., 197: 323-335.
CrossRef - Joshi, N., R.A. Rawat, B. Subramanian and K.S. Rao, 2010. A method for small scale genomic DNA isolation from chickpea (Cicer arietinum L.) suitable for molecular marker analysis. Indian J. Sci. Technol., 3: 1214-1217.
Direct Link - Kotchoni, S.O., E.W. Gachomo, E. Betiku and O.O. Shonukan, 2003. A homemade kit for plasmid DNA mini-preparation. Afr. J. Biotechnol., 2: 88-90.
Direct Link - Mazid, M., T.A. Khan and F. Mohammad, 2011. Role of secondary metabolites in defense mechanisms of plants. Biol. Med., 3: 232-249.
Direct Link - Ogunkanmi, L.A., W.O. Okunowo, O.O. Oyelakin, B.O. Oboh, O.O. Adesina, K.O. Adekoya and O.T. Ogundipe, 2010. Assessment of genetic relationships between two species of jute plants using phenotypic and RAPD markers. Int. J. Bot., 6: 107-111.
CrossRefDirect Link - Padmalatha, K. and M.N.V. Prasad, 2006. Optimization of DNA isolation and PCR protocol for RAPD analysis of selected medicinal and aromatic plants of conservation concern from Peninsular India. Afr. J. Biotechnol., 5: 230-234.
Direct Link - Singh, B., R. Yadav, H. Singh, G. Singh and A. Punia, 2010. Studies on effect of PCR-RAPD conditions for molecular analysis in asparagus (Satawari) and Aloe Vera medicinal plants. Aust. J. Basic Applied Sci., 4: 6570-6574.
Direct Link - Subudhi, P.K., N.P. Parami, M.D. Materne and S.A. Harrison, 2008. Genetic diversity in smooth cordgrass from brown marsh areas of Louisiana. J. Aquat. Plant Manage., 46: 60-67.
Direct Link - Zidani, S., A. Ferchichi and M. Chaieb, 2005. Genomic DNA extraction method from pearl millet (Pennisetum glaucum) leaves. Afr. J. Biotechnol., 4: 862-866.
Direct Link - Zhao, W. and Y. Pan, 2004. Genetic diversity of genus morus revealed by RAPD markers. Int. J. Agric. Biol., 6: 950-956.
Direct Link - Sahasrabudhe, A. and M. Deodhar, 2010. Standardization of DNA extraction and optimization of RAPD-PCR conditions in Garcinia indica. Int. J. Bot., 6: 293-298.
CrossRefDirect Link