
Ponnuswamy Vijayaraghavan
Centre for Marine Science and Technology, Manonmaniam Sundaranar University, Rajakkamangalam-629 502, Kanyakumari District, Tamil Nadu, India
Surgen A. Bright
Centre for Marine Science and Technology, Manonmaniam Sundaranar University, Rajakkamangalam-629 502, Kanyakumari District, Tamil Nadu, India
Anuj Nishanth Lipton
Department of Biotechnology, Udaya School of Engineering, Vellamodi, Kanyakumari District, Tamilnadu, India
S.G. Prakash Vincent
International Centre for Nanobiotechnology, Centre for Marine Science and Technology, Manonmaniam Sundaranar University, Rajakkamangalam-629 502, Kanyakumari District, Tamil Nadu, India
Asian Journal of Biotechnology
Year: 2011 | Volume: 3 | Issue: 5 | Page No.: 478-485







ABSTRACT
The aim of this study was to purify and characterize citric acid cycle enzyme malate dehydrogenase (MDH; EC 1. 1. 1. 37) from Pseudomonas aeruginosa. The purification steps consisted of ammonium sulphate precipitation, ion-exchange chromatography and gel filtration. A typical procedure provided 638-fold purification with 23.0% yield. Single band was observed in both native gradient-and SDS-PAGE. The molecular weight estimated for the native enzyme was 70.5 kDa whereas subunit values of 36 kDa were determined. Hence, MDH is a dimer of identical subunits. The enzyme was highly active at pH 8.0 when NADH was used as the cofactor and was highly stable at pH 7.0. The optimum temperature for the enzyme activity was recorded to be 40°C. Oxaloacetate was determined as the specific substrate with an apparent km of 10 μM. The characteristics of thermo-stability and its high activity at alkaline pH suggest its potential diagnostic, therapeutic and beverage related applications. This MDH may be of value in developing a serological test for pneumonia which is caused by P. aeruginosa.
PDF Abstract XML References Citation
Received: March 20, 2011;
Accepted: May 24, 2011;
Published: July 09, 2011
How to cite this article
Ponnuswamy Vijayaraghavan, Surgen A. Bright, Anuj Nishanth Lipton and S.G. Prakash Vincent, 2011. Purification and Properties of Novel Malate Dehydrogenase Isolated from Pseudomonas aeruginosa. Asian Journal of Biotechnology, 3: 478-485.
DOI: 10.3923/ajbkr.2011.478.485
URL: https://scialert.net/abstract/?doi=ajbkr.2011.478.485
DOI: 10.3923/ajbkr.2011.478.485
URL: https://scialert.net/abstract/?doi=ajbkr.2011.478.485
INTRODUCTION
Malate Dehydrogenase (MDH; EC 1.1.1.37) catalyzes the conversion of oxaloacetate and malate utilizing the NAD/NADH coenzyme system (Minarik et al., 2002). MDH was isolated from diverse sources including eubacteria, archaea, fungi, plants, mammals and from subcellular organelles such as mitochondria (Goward and Nicholls, 1994). In most organisms, including all eukaryotes and many bacterial species MDH occur as homodimeric molecules (Minarik et al., 2002). MDH has been purified and extensively studied from several species such as: Chlorobium vibrioforme, C. tepidum and Helicobacterium gestii (Charnock et al., 1992), syntrophic propionate oxidizing bacterium strain MPOB (Van-Kuijk and Stams, 1996), Nitrosomonas sp. TK 794 and Nitrobacter agilis ATCC 14123 (Masuko et al., 1997), Acetobacter sp. (Shinagawa et al., 2002), Beggiatoa sp. (Eprintsev et al., 2003a), sulfur bacterium Beggiatoa leptomitiformis (Eprintsev et al., 2003b), Valcanithermus medioaltalicus (Eprintsev et al., 2005) and Pyrobaculum islandicum (Yennaco et al., 2007). Recently, the MDH was characterized from the genus Aeromonas (Farfan et al., 2010) and Streptomyces coelicolor A3 (2) (Ge et al., 2010). MDH is a conjugate for enzyme immunoassay of a wide variety of compounds, particularly drugs, including drugs of abuse, drugs used in repetitive therapeutic applications, hormones (Roderick and Banaszak, 1986). It is also used to analyze the L-malate content from beverages (Mori et al., 2007). Although MDH persists in all bacteria, very few studies have been done on purification and characterization from the bacterial isolates. Considering the various biotechnological applications of MDH, investigations were carried out to purify and characterize MDH from Pseudomonas aeruginosa and the salient results are presented.
MATERIALS AND METHODS
Chemicals: Sephadex G-75 was purchased from Amersham Biosciences, Sweden. Electrophoresis chemicals, reagents and protein marker were purchased from GeneI (Presently Merck), Bangalore, India. Culture media and other salts were obtained from Himedia laboratories, Mumbai, India. All other chemicals used were of analytical grade.
Microorganism and culture: The bacterial isolate, Pseudomonas aeruginosa was obtained from the Microbiology Laboratory, Centre for Marine Science and Technology, M.S. University, Rajakkamangalam, Tamilnadu, India in the year 2009 for the isolation of MDH. Pseudomonas aeruginosa was cultured in the nutrient medium composed of in (g L-1): Peptic digest of animal tissue, 5.0; beef extract, 1.5; yeast extract, 1.5 and sodium chloride, 5.0 (pH 7.0) and sterilized at 121°C for 20 min. A loopful culture of 18 h P. aeruginosa was inoculated into the nutrient medium with rotary shaking at 150 rpm at 37°C for 24 h.
Preparation of cell-free extract: The P. aeruginosa cells were harvested (10000 rpmx10 min) (5 g as wet weight) and washed with 0.05 M Tris buffer (pH 7.4) (buffer A) . The pelleted cells were again suspended in 40 mL of buffer A and lysed with lysozyme. The homogenate was centrifuged at 10,000 rpm for 15 min and the supernatant was used as the crude extract.
MDH assay: MDH activity was assayed spectrophotometrically, usually in the direction of oxaloacetate reduction as previously described by Chaffai and El-Ferjani (2005). The standard assay mixture (3 mL) consisted 0.1 mL of 6 mM of Oxaloacetic Acid (OAA), 0.2 mL of 3 mM of NADH and 2.6 mL of 50 mM sodium phosphate buffer (pH 6.5). The reaction was started by the addition of an enzyme solution. The rate of NAD+ reduction was measured spectrophotometrically at 340 nm. One enzyme unit was defined as the amount of enzyme required for oxidation/reduction of 1 μmol of substrate/min. Specific enzyme activity was expressed in units/mg protein.
Evaluation of optimum condition for MDH activity: The pH optimum was studied using the following buffers at 3.0, 4.0, 5.0, 6.0, 7.0, 8.0, 9.0 and 10.0 at 0.1 M concentration in the reaction mixture: citrate buffer, pH 3.0 to 4.0; succinate buffer, pH 5.0; potassium phosphate buffer, pH 6.0 to 7.0; tris buffer, pH 8.0, glycine-NaOH buffer, pH 9.0 to 10.0. The effect of temperature on enzyme activity was examined at 25, 30, 35, 40, 45, 50 and 55°C (Yueh et al., 1989). The enzyme activity of MDH towards substrates (2.5 to 50 μM) such as: citrate, oxaloacetate, l-sorbose and D-fructose were evaluated (Walk and Hock, 1977). The MDH activity in response to various chemical substances was evaluated using the substances such as: EDTA, HgCl2, KCl, MgCl2 and NaCl at 1 mM concentration.
Purification of MDH: The purification steps consisted of ammonium sulphate precipitation, ion-exchange chromatography and gel filtration as suggested by Yueh et al. (1989) with little modifications. Briefly, the crude extract was fractionated by ammonium sulphate precipitation and centrifugation. The pellet obtained between 40 and 80% saturation of ammonium sulphate was dissolved in minimum volume of buffer A. The precipitate was dialyzed against 0.05 M Tris-HCl buffer, pH 7.4 containing 10 mM KCl, 1 mM MgCl2, 0.1 mM EDTA and 2 mM DTT (buffer B). The dialyzed sample was applied on pre-equilibrated DEAE cellulose column (0.6x20 cm) with buffer B. MDH was adsorbed on the column and the unbound proteins were washed off by 2 bed volume of buffer B. Washing was continued till the OD reduced to 0.003 at 280 nm. After the column had been washed with buffer to remove unbound proteins, the enzyme was eluted by an application of a 75 mL linear gradient of 0 to 0.3 M NaCl in buffer B. The fractions showing MDH activity were combined and dialyzed against buffer B. The dialyzed sample was rechromatographed on sephadex G-75 gel filtration column using buffer B. All fractions were evaluated for MDH activity.
Total protein content: Total protein content was estimated as per the method of Lowry et al. (1951), using Bovine Serum Albumin as standard.
Molecular mass determination: The native gradient polyacrylamide gel (5 to 16%) was used to determine the molecular mass of the native MDH (Nishizawa et al., 1988). The molecular mass of the subunit of the enzyme was estimated by SDS-Polyacrylamide gel electrophoresis (11.0%) (Laemmli, 1970) with myosin rabbit muscle (205 kDa), phosphorylase b (97.4 kDa), bovine serum albumin (66 kDa), ovalbumin (43 kDa) and carbonic anhydrase (29 kDa) markers. The proteins were silver-stained (Blum and Beier, 1987) and documented using a gel documentation system (Syngene, UK).
RESULTS AND DISCUSSION
Properties of MDH: The dependence of the rates of oxaloacetate reduction catalysed by the MDH on the pH of the reaction mixture was determined in the range of pH 3.0 to 10.0. The pH optimum for oxaloacetate reduction was found to be 8.0 (Fig. 1). Similar result was reported by Genda et al. (2003) in Cornybacterium glutamicum. The optimum temperature for the MDH activity from P. aeruginosa isolate was 40°C (1.4 U mL-1). However this activity declined thereafter as could be noted from Fig. 2. In Flavobacterium frigidimaris KUG-1, through the MDH activity was high at 40°C, it declined when temperature was raised to while at 70°C (Oikawa et al., 2005). This result of thermal stability could be considered as advantages to utilize the MDH for evident biotechnological applications.
Results of enzyme activity of MDH towards substrates (2.5 to 50 μM) such as citrate, oxaloacetate, l-sorbose and D-fructose indicated that the MDH of P. aeruginosa was highly specific to oxaloacetate. The km of oxaloacetate was 10 μM. Earlier report by Oikawa et al. (2005) in Flavobacterium frigidimaris KUC-1 MDH also indicated the MDH specificity towards oxaloacetate. In syntrophic propionate-oxidizing bacterium strain MPOB, the km for oxaloacetate reduction was reported as 50 μM and it was 30 μM for NADH (Van-Kuijk and Stams, 1996).
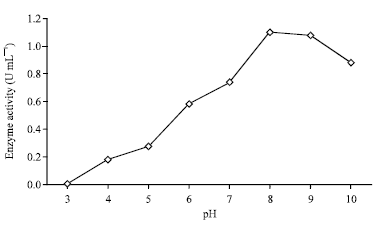 | |
| Fig. 1: | Effect of pH on enzyme activity. Optimum pH of the enzyme was determined by assaying MDH activity over the range 3.0 to 10.0 in various buffers at 0.1 M concentration |
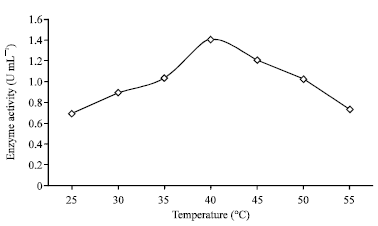 | |
| Fig. 2: | Activity of MDH at different temperatures. MDH activity was measured every 5°C at various temperatures ranging from 25 to 55°C with standard assay mixture |
| Table 1: | Effect of chemical substances on enzyme activity |
 | |
In response to various chemical substances, results indicated the moderate inhibition by EDTA and magnesium chloride and the remaining enzyme activity was 91.5 and 88.8%. Other chemical substances, such as mercuric chloride, potassium chloride and sodium chloride stimulated MDH activity and the related enzyme activity was 111, 126 and 137%, respectively (Table 1). Similar trend of activities of MDH from Flavobacterium frigidimaris KUC-1 was reported by Oikawa et al. (2005).
Purification of malate dehydrogenase: The MDH of the cell free culture of Pseudomonas aeruginosa was purified for homogeneity by a three-step procedure: ammonium sulphate precipitation, DEAE cellulose chromatography and sephadex G-75 gel filtration.
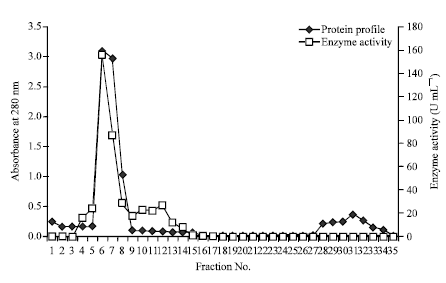 | |
| Fig. 3: | Elution profile of Pseudomonas aeruginosa MDH from a DEAE cellulose column. The column was equilibrated with Tris-HCl buffer, pH 7.4 containing 10 mM KCl, 1mM MgCl2, 0.1 mM EDTA and 2 mM DTT (buffer B) and the bounded enzyme was eluted from the column after the application of a linear gradient of 0 to 0.3 M of NaCl with buffer B |
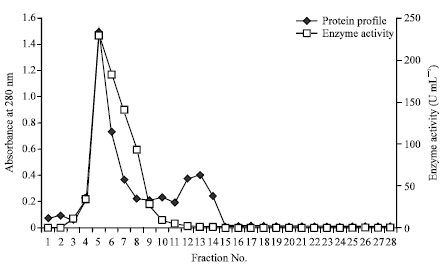 | |
| Fig. 4: | Elution profile of Pseudomonas aeruginosa MDH from sephadex G 75 gel filtration column. The column was pre-equilibrated with buffer B and eluted with the same |
The DEAE cellulose chromatography was still the major technique for purification of Pseudomonas aeruginosa MDH, because majority of contaminating proteins were removed at this step. One major peak was observed in this chromatography and 250 fold purification was achieved with 39% yield (Fig. 3). In sephadex G-75 gel filtration chromatography, the MDH could be purified about 638 fold purification with 23% yield (Fig. 4). The specific activity of MDH was 0.54 U mg-1 in crude sample, however after two chromatography steps of purification the specific activity was 255 U mg-1. Similar purification procedures have been used for the isolation of MDH with Pseudomonas testosteronei and over 450 fold of purification was obtained (You and Kaplan, 1975). A typical purification experiment is summarized in Table 2.
| Table 2: | Purification summary of malate dehydrogenase from Pseudomonas aeruginosa |
 | |
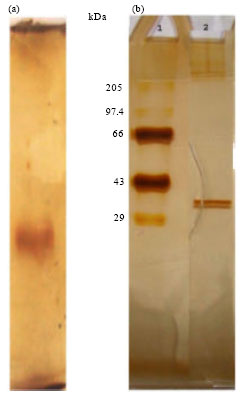 | |
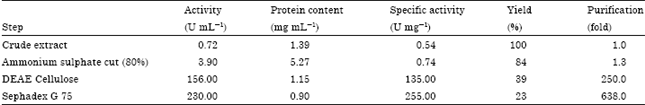
| Fig. 5 (a-b): | (a) Gradient non-denaturing polyacrylamide gel electrophoresis and (b) SDS-PAGE of purified MDH. (Lane 1: MW standards (in kDa), Lane 2: Purified MDH) |
Molecular weight and subunit structure: The purified enzyme was found to be homogenous on native gradient polyacrylamide gel electrophoresis and sodium dodecyl sulphate-polyacrylamide gel electrophoresis. From the native gradient gel, the molecular weight could be noted as 70.5 kDa (Fig. 5a). In the SDS-PAGE, the purified enzyme migrated as a single band with an apparent molecular weight of 36 kDa (Fig. 5b) suggesting it to be a homodimer. The presence of identical two subunits in Pseudomonas testosteronei with a molecular weight of 74 kDa was noted by You and Kaplan (1975). In P. stutzeri, Labrou and Clonis (1997) indicated the molecular weight of MDH as 66.5 kDa.
CONCLUSION
The citric acid cycle enzyme malate dehydrogenase was purified to homogeneity from Pseudomonas aeruginosa isolate. The MDH was purified over 600 fold using conventional methods. The characteristics of thermo-stability and its high activity at alkaline pH suggest its potential diagnostic, therapeutic and beverage related applications. This MDH may be of value in developing a serological test for pneumonia which is caused by P. aeruginosa.
ACKNOWLEDGMENTS
The authors are thankful to Dr. A.P. Lipton, Principal Scientist, Vizhinjam Research Centre of CMFRI, India for reviewing the manuscript.
REFERENCES
- Blum, H., H. Beier and H.J. Gross, 1987. Improved silver staining of plant proteins, RNA and DNA in polyacrylamide gels. Electrophoresis, 8: 93-99.
CrossRefDirect Link - Chaffai, R. and E.El-Ferjani, 2005. Alteration of the profile of organic acid content and exudation under aluminum stress in maize (Zea mays L.). Int. J. Bot., 1: 64-73.
CrossRefDirect Link - Charnock, C., U.H. Refseth and R. Sirevag, 1992. Malate dehydrogenase from Chlorobium vibrioforme, Chlorobium tepidum and Heliobacterium gestii: Purification, characterization and investigation of dinucleotide binding by dehydrogenases by use of empirical methods of protein sequence analysis. J. Bacteriol., 174: 1307-1313.
PubMed - Eprintsev, A.T., M.I. Falaleeva, I.Y. Stepanova, N.V. Parfenova and M. Zuzu, 2003. Isolation, purification, and properties of malate dehydrogenase from sulfur bacterium Beggiatoa leptomitiformis. Biol. Bull., 30: 243-247.
CrossRef - Eprintsev, A.T., M.I Falaleeva and N.V. Parfenova, 2005. Malate dehydrogenase from the thermophilic bacterium Vulcanithermus medioatlanticus. Biochemistry, 70: 1027-1030.
CrossRef - Farfan, M., D. Minana-Galbis, A. Garreta, J.G. Loren and M.C. Fuste, 2010. Malate dehydrogenase: A useful phylogenetic marker for the genus Aeromonas. Syst. Applied Microbiol., 33: 427-435.
PubMed - Ge, Y.D., Z.Y. Cao, Z.D. Wang, L.L. Chen, Y.M. Zhu and G.P. Zhu, 2010. Identification and biochemical characterization of a thermostable malate dehydrogenase from mesophile Streptomyces coelicolor. Biosci. Biotechnol. Biochem., 74: 2194-2201.
CrossRef - Genda, T., T. Nakamatsu and H. Ozak, 2003. Purification and characterization of malate dehydrogenase from Corynebacterium glutamicum. J. Biosci. Bioeng., 95: 562-566.
CrossRef - Goward, C.R. and D.J. Nicholls, 1994. Malate dehydrogenase: A model for structure, evolution and catalysis. Protein Sci., 3: 1883-1888.
Direct Link - Labrou, N.E. and Y.D. Clonis, 1997. L-Malate dehydrogenase from Pseudomonas stutzeri: Purification and characterization. Arch. Biochem. Biophys., 337: 103-114.
CrossRef - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Masuko, T., R. Takahashi and T. Tokuyama, 1997. Malate dehydrogenases from nitrifying bacteria: Purification and properties. Soil Microorganisms, 50: 51-59.
Direct Link - Minarik, P., N. Tomaskova, M. Kollarova and M. Antalik, 2002. Malate dehydrogenases-structure and function. Gen. Physiol. Biophys., 21: 257-265.
PubMed - Mori, H., A. Yamashila and S. Maitani, 2007. Determination of L-malate using immobilized malate dehydrogenase and aspartate aminotransferase in a flow system and its application to analyze the L-malate content of beverages. J. Health Sci., 53: 128-131.
Direct Link - Nishizawa, H., N. Kita, S. Okimura, E. Takao and Y. Abe, 1988. Determination of molecular weight of native proteins by polyacrylamide gradient gel electrophoresis. Electrophoresis, 9: 803-806.
PubMed - Oikawa, T., N. Yamamoto, K. Shimoke, S. Uesato, T. Ikeuchi and T. Fujioka, 2005. Purification, characterization and overexpresssion of psychrophilic and thermolabile malate dehydrogenase of a novel antarctic psychrotolerant Flavobacterium frigidimaris KUC-1. Biosci. Biotechnol. Biochem., 69: 2146-2154.
PubMed - Roderick, S.L. and L.J. Banaszak, 1986. The three-dimensional structure of porcine heart mitochondrial malate dehydrogenase at 3.0 A resolution. J. Biol. Chem., 261: 9461-9464.
PubMed - Shinagawa, E., T. Fujishima, D. Moonmangmee, G. Theeragool, H. Toyama, K. Matsushita and O. Adachi, 2002. Purification and characterization of membrane-bound malate dehydrogenase from Acetobacter sp. SKU 14. Biosci. Biotechnol. Biochem., 66: 298-306.
PubMed - Walk, R.A. and B. Hock, 1977. Glyoxysomal and mitochondrial malate dehydrogenase of watermelon (Citrullus vulgaris) cotyledons. II Kinetic properties of the purified isoenzymes. Planta, 136: 221-228.
CrossRef - You, K.S. and N.O. Kaplan, 1975. Purification and properties of malate dehydrogenase from Pseudomonas testosteroni. J. Bacteriol., 123: 704-716.
PubMed - Yueh, A.Y., C.S. Chung and Y.K. Lai, 1989. Purification and molecular properties of malate dehydrogenase from the marine diatom Nitzschia alba. Biochem. J., 258: 221-228.
PubMed - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link