
Reetesh Kumar
Molecular Biology Unit, Institute of Medical Sciences, Banaras Hindu University, India
Sanjay K. Jaiswal
Department of Genetics and Plant Breeding, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi, India
Manish K. Vishwakarma
Department of Genetics and Plant Breeding, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi, India
LiveDNA: 91.17474
Ved P. Rai
Department of Genetics and Plant Breeding, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi, India
Priti Upadhyay
Department of Genetics and Plant Breeding, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi, India
Arun K. Joshi
Department of Genetics and Plant Breeding, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi, India
Asian Journal of Biotechnology
Year: 2011 | Volume: 3 | Issue: 5 | Page No.: 460-469







ABSTRACT
Evaluation of genetic diversity was made using RAPD as a genetic marker system in allohexaploid wheat (T. aestivum). A set of 30 RAPD markers were used for 43 wheat cultivars to examine their utility for detecting DNA polymorphism in genotypes and estimating genetic diversity among them. A total of 228 different RAPD bands were scored for the presence of bands among all the varieties. Out of 30 oligonucleotide primers used, 5 primers gave reproducible DNA product (bands) suitable for the establishment of genetic diversity among the varieties. The Genetic Similarity (GS) coefficient for all pairs of genotypes was ranged from 0.66 to 1.0. V 35 (HD 2747) and V 39 (HI 8498) had high coefficient of similarity (0.982) while, V 4 (VL 804) and V 6 (VL 809) showed less coefficient of similarity (0.646). It indicated that V 35 (HD 2747) and V 39 (HI 8498) carry closely related ancestry while the cultivars V 4 (VL 804) and V 6 (VL 809) are not close to each other. Present results revealed the potential of RAPD technique in determining genetic diversity and make suitable fingerprints for wheat cultivars.
PDF Abstract XML References Citation
Received: March 16, 2011;
Accepted: May 24, 2011;
Published: July 09, 2011
How to cite this article
Reetesh Kumar, Sanjay K. Jaiswal, Manish K. Vishwakarma, Ved P. Rai, Priti Upadhyay and Arun K. Joshi, 2011. Assessment of Genetic Diversity and its Usefulness for Varietal Identification in Indian Elite Varieties of Wheat (T. aestivum L. em thell.) Using RAPD Markers. Asian Journal of Biotechnology, 3: 460-469.
DOI: 10.3923/ajbkr.2011.460.469
URL: https://scialert.net/abstract/?doi=ajbkr.2011.460.469
DOI: 10.3923/ajbkr.2011.460.469
URL: https://scialert.net/abstract/?doi=ajbkr.2011.460.469
INTRODUCTION
India is the second largest producer of wheat in the world with production hovering around 70-75 million tons in the past few years (Nagarajan, 2005). At present, Uttar Pradesh, Punjab and Haryana account for nearly 80% of the total wheat production. Around 312 wheat varieties were released during past 40 years in India’s wheat zones and played a major role in enhancing productivity (Chatrath et al., 2006). Wheat production increased from 6.5 million tons in 1950, to the 72 million tons produced in 2005, a tenfold increase (DWR, 2006). Resultantly, increased yields per hectare that went from around 660 kg ha-1 in 1950 to 2,710 kg ha-1 in 2004 (FAO, 2004). Alongside increased yields came an increase in area planted from nearly 9.8 million hectares in 1950 to 24 million hectares in 1990, to about 27 million hectares in 2004 (FAO, 2004).
Genetic variability is of prime momentous for the improvement of many crop species including wheat. Genetic improvement of any crops depends upon the existence, nature and range of genetic diversity available for manipulation (Waugh and Powell, 1992). This not only gives breeders an idea to know the dimension of genetic diversity being used for commercial cultivation but also enables them to find ways of finding suitable donors of different genes. The forecasted global demand for wheat in the year 2020 is approximately 87.5 million tons, or about 13 million tons more than the record production of 75 million tons harvested in crop season 1999-2000 (Joshi et al., 2007). To achieve this target global production will need to increase 1.6 to 2.6% annually from the present production level. In addition, knowledge of the genetic diversity through characterization of varieties would also helps in proper identification of genotypes under use. The growing acceptance of Intellectual Property Rights all over the world also demands proper identification of genotypes for deciding monetary claims and disputes. Nearly all crop improvement programmes depends on genetic diversity in the available germplasm (Graner et al., 1994; Sorrells and Wilson, 1997) and molecular markers can be broadly classified in hybridization based DNA markers, PCR-based markers and sequencing-based markers (Gupta et al., 1999).
The PCR (polymerase chain reaction) based methods have now been accepted as useful method to detect genetic variability due to their speed and low cost. In a RAPD reaction, a synthetic oligonucleotide of 9 to 12 bases is used as starting point for DNA amplification with thermostable DNA polymerases (Williams et al., 1990). For wheat, RAPD marker has been widely used for genetic diversity analysis (Vierling and Nguyen, 1992; Bibi et al., 2009; Khan et al., 2010; Ahmed et al., 2010).
With the aim to assess the genetic diversity and its characterization, the present investigation on wheat was initiated with forty three different varieties using thirty RAPD markers.
MATERIALS AND METHODS
Forty-three wheat cultivars released in India during last 50 years of breeding in which some of them were very promising varieties. Seeds of these were obtained from Institute of Agricultural Sciences, BHU, Varanasi, India, for investigating the genetic diversity. The seeds of collected accessions were sown in a plastic pot in January-February 2004 at Banaras Hindu University, India. Approximately 18-days leaves were collected for DNA isolation. The list of cultivars with their pedigree is given in Table 1. Plant genomic DNA was extracted by a CTAB (Cetyl trimethyl ammonium bromide) method (Saghai-Maroof et al., 1984). DNA quality was verified by gel electrophoresis on agarose gel 1%. DNA samples were diluted to a concentration of 50 μL for using in the polymerase chain reaction.
Total of 30 RAPD primers were used for the analysis of genetic diversity in the all wheat cultivars Table 2. The amplification conditions were rigorously tested in optimization experiments describe below in the results section. During the experiment we have used PCR volume 25 μL and contained 1 μL Primer, 0.8 μL DNTPs, 2 μL template DNA and 0.5 μL Taq DNA polymerase including 2.5 X PCR Buffer. The reaction mixtures were overloaded with mineral oil and subjected to PCR. Standard amplification were performed in a thermal cycler programmed for 60 cycle of 1 min at 94°C, 1 min at 36°C and 2 min at 72°C, using the fastest possible transition between each temperature. After the last cycle, the sample were kept at 72°C for an additional 5 min and then cooled at 4°C. Samples were analyzed by electrophoresis on a 1.5% agarose gel and amplified product was detected by staining with ethidium bromide.
Statistical analysis: RAPD bands were scored for the presence (1) or absence (0) of bands among all the varieties. Data were analyzed and similarity matrix was constructed from binary data with Jaccard’s coefficients (Jaccard, 1908) and cluster analysis was done using UPGMA (unweighted pair-group method with arithmetic average. The RAPD primer generated fragment length was estimated by AlphaVIEW SA software provided by Alpha Innotech USA.
| Table 1: | List of varieties and their pedigree used for RAPD fingerprinting |
 | |
| Table 2: | Lest of primers used |
 | |
RESULTS
In this investigation genetic diversity analysis was performed using RAPD primers with forty three cultivars of wheat. Bands with the same mobility were treated as identical fragments while those with different mobility were treated as non-identical bands. A high degree of variation was noticed among wheat cultivars and a total of 228 different RAPD bands were scored for the presence or absence of bands. The result of RAPD screening has been presented here in Table 3. Five RAPD primers namely OPG-07 (TTCGAGCCAG), OPG-08 (GAACGGACTC), OPG-09 (TGTCTGGGTG), OPG-11 (TGCCCGTCGT) and OPG-12 (CAGCTCACGA) gave reproducible bands out of 30 oligonucleotide primers and these five can be used for genetic diversity assessment of wheat varieties. The above said five primers showed polymorphism in all the wheat cultivars.
The RAPD patterns differentiating among 43 wheat cultivars amplified with two primer especially OPG-11 and OPG-12 that displayed maximum polymorphism have been presented in Fig. 1 and 2. Cultivars HW3082, VL804, RAJ6557 (D), GW1170 (D), HW3061, HW3064, HW2017, HW3067, UP2473, PDW274 (D), FLW3 produced unique band (~1375 bp), genomic DNA of cultivars HW3080, HD2755, MACS2846 (D) amplified unique band of ~1305 bp and ~1680 bp amplified unique band was produced with cultivars VL809, HW3033, TL2910, HW3024, HW3027, HP1731, PDW278 (D), PDW274 (D), HI8498 (D), HD4672 and FLW2 whereas single cultivar VL803 was made by itself due to the presence of single unique band of ~1584 bp by the primer OPG-11 (Fig. 2).
| Table 3: | Bands obtained in the RAPD analysis |
 | |
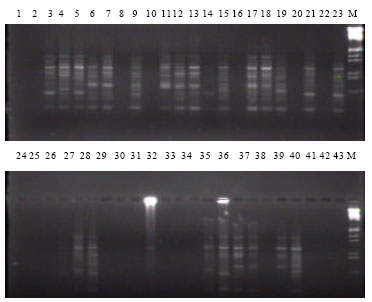 | |
| Fig. 1: | RAPD patterns differentiating among 43 isolates of wheat are amplified with arbitrary primer OPG-11. M is molecular marker DNA/Eco RI+Hind III |
Using above 5 RAPD primers, total number of amplified band were 228 and total number of polymorphic bands are 110. Average number of band per primer was observed 45.6 and average number of polymorphic bands per primer was recorded as 22 (Table 3).
The binary data generated after scoring of RAPD bands, it was analyzed using NTSYS pc 1.80 software (Rohlf, 1993). Genetic similarity among samples was estimated using a distance matrix based on RAPD bands scored and Jaccard (J) coefficient of similarity. Samples were divided into different clusters by their distribution on the dendrogram (Fig. 3). In generating dendrogram we got a good arrangement between the sample stocks of wheat cultivars. Therefore, it appears that the dendrogram obtained represent true genetic relationship between all wheat samples analyzed. In the dendrogram, genetic distance or coefficient of similarity among wheat cultivars ranged from 0.66 to 1.00. Those cultivars that displayed similar coefficient of similarity are genetically close to one another while those having variable coefficient of similarity and are dissimilar. For instance, cultivar 42 (FLW 2) and cultivar 43 (FLW 3) had similar coefficient of similarity (1.00). It suggested that these varieties are genetically too close to each other and are probably selection from the same pedigree/cross.
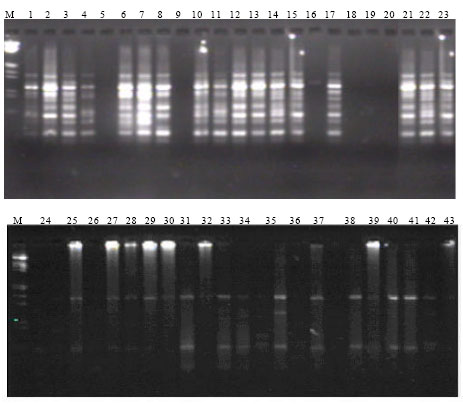 | |
| Fig. 2: | RAPD patterns differentiating among 43 isolates of wheat are amplified with arbitrary primer OPG-12. M is molecular marker DNA/Eco RI+Hind III |
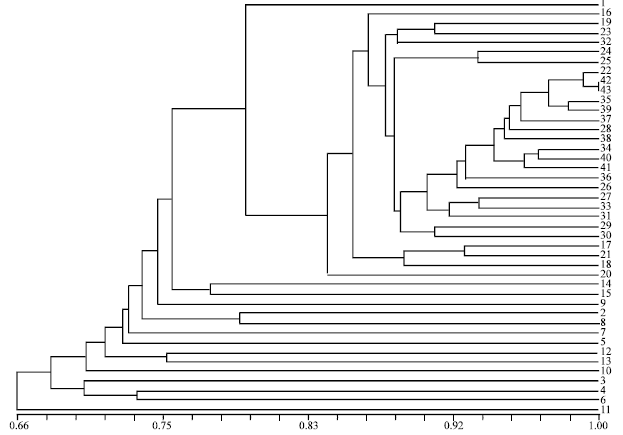 | |
| Fig. 3: | Dendrogram of RAPD analysis of 43 wheat cultivars |
Similarly, V 35 (HD 2747) and V 39 (HI 8498) had high coefficient of similarity (0.982) while, V 4 (VL 804) and V 6 (VL 809) showed less coefficient of similarity (0.646). It clearly showed that V 35 (HD 2747) and V 39 (HI 8498) have come closely related in their ancestry while the cultivars V 4 (VL 804) and V 6 (VL 809) are distant from each other. In the same way, cultivars V 35 (HD 2747) and V 37 [PDW 274 (D)] and V 6 (VL 809) and V 17(HW 3024) are genetically dissimilar with each other.
DISCUSSION
The availability of genetic variability is a pre requisite for any breeding programme (Sharma, 1995). Earlier, pedigree analysis of cultivars and lines was used to be the criteria to establish genetic diversity among them (Noiton and Alspach, 1996). However, this approach was confounded by inaccurate pedigree records, absence of pedigree records or misnamed and/or multiple named cultivars. Molecular characterization of germplasm is basic to the improvement of the species and can be done at the DNA level. Now a day, it is accepted that improved technology and methods are needed to reduce cost and simplify the task of mapping genes and for determining their relative allelic value in our cultivated crops and varieties (Sorrells and Wilson, 1997). For these reasons, as molecular marker technologies become available, they are being used widely for two objectives viz., creation of genetic variability and marker-assisted selection. Molecular tools are also being used for fingerprinting or genetic characterization of varieties of different crops. The RAPD procedure used in this study is simple well established and polymorphisms detected are repeatable. Due to the short length of the applied primers, any arbitrary sequence is expected to provide an equal chance of amplification under low stringency (Williams et al., 1990).
In this investigation, 30 primers were used to establish genetic diversity among all wheat genotypes. Among these, five primers OPG-07, OPG-08, OPG-09, OPG-11 and OPG-12 were found to be useful. However, two primers OPG-11 and OPG-12 primers produced the best result (Fig. 2a, b). The remaining 25 primers were either indistinct or monomorphic in nature. This low polymorphism is in agreement with some of the earlier reports which have also suggested low polymorphism by RAPD primers (Vidal et al., 1999; Ramshini et al., 2005; Prakash et al., 2006; Ahmed et al., 2010). RAPD technique is considered as a very sensitive method of screening for nucleotide sequence polymorphisms randomly distributed throughout the genome (Subudhi and Huang, 1999). Experimental evidences in Arabidopsis and Neurospora genomes (Williams et al., 1993) and maize (Beaumont et al., 1996) have indicated no clear bias in distribution of RAPD sites on the genome with the exception of a report of clustering in sugar beet (Nilsson et al., 1997) and in this study too RAPD followed the same.
The percentage of polymorphic bands displayed by 30 primers was 48.24% (110 bands).The Dendrogram generated by the analysis of RAPD data had one major cluster which contains twenty genotypes. Others genotypes failed to cluster as group. The reason behind this grouping nature may be that, there is no similarity in any trait. The RAPD primers amplified different alleles which would not at all relate to common characters. The highest genetic similarity was observed in the cultivars FLW2 and FLW3 (100%) and lowest genetic similarity was observed in VL 803 and GW 110 (57%). Both the genotypes (FLW2 and FLW3) have been reported for leaf rust resistance gene Lr26 derived from 1B/1R translocation reported in very cultivars that dominated in 1980’s (Schachermayr et al., 1994; Sudhakar and Joshi, 1996) and similarity observed between these two might have this basis. The role of physiological trait in grouping plays an important role (Upadhyay et al., 2011). The resistance trait playing here a role in this grouping is in consonance with the study of Upadhyay et al. (2011). It is also possible that there may be common parents in their ancestry at any stage. This study is in accordance with the study on bacterial population using eBURST software for assessing the population structure (Turner et al., 2007). The wide range of genetic similarity observed in the 43 wheat cultivars studied established that substantial genetic diversity were present among these cultivars. The genetic characterization obtained among these wheat cultivars can be used to plan crossing programme for obtaining desirable transgressive segregants as it is supported by the study performed by Tyagi and Khan (2010) in Lentil. By using more wheat genotypes for fingerprint, scope of such advantage in the ongoing wheat breeding programme of different centers can further be enhanced.
In this study, typical banding pattern was observed for RAPD which confirms that the PCR conditions were optimized properly and it can be positively used for genetic diversity characterization and SCAR development. SCARs developed from these RAPD markers would be helpful for identification varieties under question (Hernandez et al., 1999). This in turn can also be used for determining the genetic purity of a seed lot or even for identifying the pedigree of an unknown line. The results obtained in the study further confirm the usefulness of RAPD as a potent tool in the assessment of genetic diversity and creating suitable fingerprints of Indian wheat cultivars.
Based on the findings of this study, following conclusions can be drawn. Substantial genetic diversity was present among the 43 wheat cultivars investigated. The primers OPG-12 (CAGCTCACGA) and OPG-11 (TGCCCGTCGT) displayed maximum polymorphism among wheat varieties. The genetic similarity value was highest between cultivars 42 and 43. Lowest genetic similarity was observed between cultivar 4 and 6. RAPD proved as a potent tool in determining genetic diversity and designing suitable fingerprints of wheat cultivars. The reproducibility of the RAPD technique can be influenced by variable factors, such as primer sequence, template quality and quantity, the type of thermocycler and polymerase concentration (Hernandez et al., 1999). However, the use of a standardized RAPD protocol can ensure a reproducible RAPD pattern.
CONCLUSION
RAPD technique was found to be quite effective in determining the genetic variation among wheat genotypes and in combination with other PCR based marker systems it can also be utilized as DNA fingerprinting for variety identification and for the establishment of plant breeder rights.
REFERENCES
- Ahmed, M.F., M. Iqbal, M.S. Masood, M.A. Rabbani and M. Munir, 2010. Assessment of genetic diversity among Pakistani wheat (Triticum aestivum L.) advanced breeding lines using RAPD and SDS-PAGE. Electronic J. Biotechnol., 13: 1-10.
Direct Link - Beaumont, V.H., J. Mantet, T.R. Rocheford and J.M. Widholm, 1996. Comparision of RAPD and RFLP markers for F2 generation in maize (Zea mays L.). Theor. Applied Genet., 93: 606-612.
CrossRef - Bibi, S., M.U. Dahot, I.A. Khan, A. Khatri and M.H. Naqvi, 2009. Study of genetic diversity in wheat (Triticum aestivum) using Random Amplified Polymorphic DNA (RAPD) markers. Pak. J. Bot., 41: 1023-1027.
Direct Link - Graner, A., W.F. Ludwig and A.E. Melchinger, 1994. Relationship among European barley germplasm. II. Comparison of RFLP and pedigree data. Crop Sci., 34: 1199-1205.
Direct Link - Gupta, P.K., R.K. Varshney, P.C. Sharma and B. Ramesh, 1999. Molecular markers and their applications in wheat breeding. Plant Breed., 118: 369-390.
CrossRefDirect Link - Hernandez, P., A. Martin and G. Dorado, 1999. Development of SCARs by direct sequencing of RAPD products: A practical tool for the introgression and marker-assisted selection of wheat. Mol. Breed., 5: 245-253.
CrossRefDirect Link - Prakash, S.P.J., K.R. Biji, S.M. Gomez, K.G. Murthy and R.C. Babu, 2006. Genetic diversity analysis of sorghum (Sorghum bicolour L. Moench) accessions using RAPD markers. Indian J. Crop Sci., 1: 109-112.
Direct Link - Joshi, A.K., B. Mishra, R. Chatrath, G.O. Ferrara and R.P. Singh, 2007. Wheat improvement in India: Present status, emerging challenges and future prospects. Euphytica, 157: 431-446.
CrossRef - Turner, K.M., W.P. Hanage, C. Fraser, T.R. Connor and B.G. Spratt, 2007. Assessing the reliability of eBURST using simulated populations with known ancestry. BMC Microbiol., 7: 30-30.
CrossRef - Khan, A.A., A. Iqbal, F.S. Awan and I.A. Khan, 2010. Genetic diversity in wheat germplasm collections from Balochistan province of Pakistan. Pak. J. Bot., 42: 89-96.
Direct Link - Nagarajan, S., 2005. Can India produce enough wheat even by 2020. Curr. Sci., 89: 1467-1471.
Direct Link - Nilsson, N.O., C. Hallden, M. Hanson, A. Hjerdin and T. Sall, 1997. Comparing the distribution of RAPD and RFLP markers in a high density linkage map of sugar beet. Genome, 40: 644-651.
PubMed - Noiton, D.A.M. and P.A. Alspach, 1996. Founding clones, inbreeding, ancestral and status number of modern apple cultivars. J. Am. Soc. Hort. Sci., 121: 773-782.
Direct Link - Ramshini, H., M.R. Nagavi and H. Alizadeh, 2005. Comparision of genetic diversity based on total and sharp bands of RAPD data in wheat. Asian J. Plant Sci., 4: 123-127.
Direct Link - Vierling, R.A. and H.T. Nguyen, 1992. Use of RAPD markers to determine the genetic diversity of diploid, wheat genotypes. Theor. Applied Genet., 84: 835-838.
CrossRefDirect Link - Saghai-Maroof, M.A., K.M. Soliman, R.A. Jorgensen and R.W. Allard, 1984. Ribosomal DNA spacer-length polymorphisms in barly: Mendelian inheritance, chromosomal location, and population dynamics. Proc. Natl. Acad. Sci., 81: 8014-8018.
CrossRefDirect Link - Schachermayr, G., H. Siedler, M.D. Gale, H. Winzeler, M. Winzeler and B. Keller, 1994. Identification and localization of molecular markers linked to the Lr9 leaf rust resistance gene of wheat. Theor. Applied Genet., 88: 110-115.
CrossRef - Sorrells, M.E. and W.A. Wilson, 1997. Direct classification and selection of superior alleles for crop improvement. Crop Sci., 37: 691-697.
Direct Link - Subudhi, P.K. and N. Huang, 1999. RAPD mapping in a doubled haploid population of rice (Oryza sativa L.). Hereditas, 130: 41-49.
CrossRefPubMedDirect Link - Sudhakar, Y. and A.K. Joshi, 1996. Genetics of resistance to Puccinia Recondita in two wheat crosses. Plant Breed., 115: 85-88.
CrossRef - Tyagi, S.D. and M.H. Khan, 2010. Genetic divergence in Lentil. Afr. Crop Sci. J., 18: 69-74.
Direct Link - Upadhyay, P., V.K. Singh and C.N. Neeraja, 2011. Identification of specific alleles and molecular diversity assessment of popular rice (Oryza sativa L.) varieties of India. Int. J. Plant Breed. Genet., 5: 130-140.
Direct Link - Vidal, J.R., M. Coarer and A. Defontaine, 1999. Genetic relationships among grapevine grown in different French and Spanish regions based on RAPD markers. Euphytica, 109: 161-172.
CrossRef - Waugh, R. and W. Powell, 1992. Using RAPD markers for crop improvement. TIBTECH, 10: 186-191.
Direct Link - Williams, J.G.K., M.K. Hanafey, J.A. Rafalski and S.V. Tingey, 1993. Genetic analysis using random amplified polymorphic DNA markers. Methods Enzymol., 218: 704-740.
Direct Link - Williams, J.G.K., A.R. Kubelik, K.J. Livak, J.A. Rafalski and S.V. Tingey, 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res., 18: 6531-6535.
CrossRefPubMedDirect Link - Jaccard, P., 1908. Nouvelles recherches sur la distribution florale. Bull. Soc. Vaud. Sci. Nat., 44: 223-270.
CrossRefDirect Link