
Roohi
Protein Research Laboratory, Department of Biotechnology and Microbiology, Integral University, Lucknow-226026, India
M. Kuddus
Protein Research Laboratory, Department of Biotechnology and Microbiology, Integral University, Lucknow-226026, India
I.Z. Ahmad
Protein Research Laboratory, Department of Biotechnology and Microbiology, Integral University, Lucknow-226026, India
J.M. Arif
Protein Research Laboratory, Department of Biotechnology and Microbiology, Integral University, Lucknow-226026, India
Asian Journal of Biotechnology
Year: 2011 | Volume: 3 | Issue: 5 | Page No.: 449-459







ABSTRACT
Cold-active α-amylases provided a large biotechnological potential and offers numerous economical and ecological benefits through energy saving process. It also minimizes undesirable chemical reactions that could occur at high temperatures. The objective of present study was to isolate novel bacterial strain for production of cold-active α-amylase. On the basis of primary screening thirty cold-active α-amylase producing bacteria were isolated from soil of Gangotri glacier, Western Himalaya, India. The isolated bacteria were subjected to enzyme production and one potential isolate (GA2) was selected for production optimization. The α-amylase production was found maximum (5870 units) at 20°C and pH 9 after 120 h incubation. Among the carbon and nitrogen sources, lactose (1%) and yeast extract (1%) was found best source, respectively. The isolate GA2 (Gangotri amylolytic) was resistant to penicillin (10 μg) among tested antibiotics and as per plasmid curing results, amylase production was a plasmid mediated characteristic. The phylogenetic analysis revealed that GA2 have highest homology with Microbacterium foliorum (99%). This was the first report on cold-active α-amylase production by M. foliorum GA2 and on the basis of 16S rRNA sequences also got an accession number HQ832574 from NCBI.
PDF Abstract XML References Citation
Received: April 28, 2011;
Accepted: June 03, 2011;
Published: July 09, 2011
How to cite this article
Roohi, M. Kuddus, I.Z. Ahmad and J.M. Arif, 2011. Production of Cold-active Extracellular α-Amylase by Newly Isolated Microbacterium foliorum GA2 from Gangotri glacier, Western Himalaya, India. Asian Journal of Biotechnology, 3: 449-459.
DOI: 10.3923/ajbkr.2011.449.459
URL: https://scialert.net/abstract/?doi=ajbkr.2011.449.459
DOI: 10.3923/ajbkr.2011.449.459
URL: https://scialert.net/abstract/?doi=ajbkr.2011.449.459
INTRODUCTION
Amylases constitute one of the most important groups of industrial enzymes and account for nearly 30% of the total sale of enzymes (Bordbar et al., 2005). Alpha amylases (EC 3.2.1.1) randomly cleaves the 1,4-α-D-glucosidic linkages between the adjacent glucose units in linear amylose chain of starch (Regulapati et al., 2007). Amylases are used in a wide range of starch industries such as baking, brewing, distillery, textile, paper and detergent industry. Amylases also have numerous biotechnological applications such as food processing, pharmaceutical and bioremediation (Burhan et al., 2003; Haki and Rakshit, 2003; Tonkova, 2006). From the past decade it has been found that cold adapted microorganisms and their enzymes provided a large biotechnological potential, offering numerous economical and ecological advantages over the use of organisms and their enzymes which operate at higher temperatures (Ohgiya et al., 1999; Cavicchioli et al., 2002; Margesin et al., 2002; Gomes and Steiner, 2004; Marchi et al., 2007). Cold-active α-amylases offer economic benefits through energy savings by overcoming the heating requirements and minimize undesirable chemical reactions that could occur at high temperatures (Gerday et al., 2000; Georlette et al., 2004; Ramteke and Bhatt, 2007). In food industry cold-active enzymes has particular significance because it prevent any modification of the original heat-sensitive substrates and products (Cavicchioli et al., 2002). The enzymes produced by cold-tolerant bacteria have commercial interest because they might be used in low temperature applications due to optimum activity at moderate to low temperatures. These enzymes are also of scientific interest to study the relationship between protein structure and thermal stability (Smith and Zahnley, 2005). Around 85% of earth is occupied by cold ecosystems including the ocean depths, polar and alpine regions (Feller and Gerday, 2003). Comparatively little information is available on enzymes from microorganisms isolated from glacier regions, which may be an ideal habitat for cold-adapted micro-organisms and their enzymes. In the present study novel bacterial strain has been isolated from Gangotri glacier and production of cold-active α-amylase is optimized through fermentation technology.
MATERIALS AND METHODS
Isolation of cold-active α-amylase producing bacteria: In order to isolate cold-tolerant bacteria, the soil samples were collected from vicinity of Gangotri glacier, Western Himalaya during winter season of 2010. The glacier is situated in between 30°44'-30°56'N and 79°04'-79°15' E having temperature 5-2°C in summer and subzero in winter. The samples were serially diluted in sterile cold saline solution and subjected to isolation of amylase producing microbes at low temperature (Mishra and Behera, 2008). Production of α-amylase was carried out by using the method of Abe et al. (1988). The assay was based on reduction in blue color intensity resulting from enzyme hydrolysis of starch (Swain et al., 2006).
Optimization of enzyme production
Incubation period and growth kinetics: Effect of incubation period on enzyme production and cell growth was determined by inoculating production media with bacterial isolate (1%) and incubated at 20±2°C for different time intervals (24-168 h) at 120 rpm. Amylase production was estimated after every 24 h for 8 days using the method described by Swain et al. (2006). Growth kinetics was obtained by measuring the cell density at 660 nm.
Effect of temperature, pH and agitation: To find out the optimum temperature for cold-active α-amylase production, the inoculated production media was incubated at different temperatures (4-50°C). Optimum pH was established by using production media having different pH (pH 5-12) (Kar and Ray, 2008). To optimize the best condition for fermentation, the inoculated production media was incubated in static condition and in a rotary shaker (120 rpm) at 20±2°C. Amylase activity was measured as per standard protocol (Swain et al., 2006).
Effect of carbon/nitrogen sources and metal ions: The effect of carbon and nitrogen sources as additional supplement in media was studied to maximize the enzyme production. Different carbon sources (1%) such as lactose, maltose, glucose, sucrose and glycerol and nitrogen sources viz. casein, glycine, yeast extract, ammonium acetate and ammonium sulfate (1%) were tested. To evaluate the impact of metal ions, media was supplemented with maximum tolerance level of different metal ions such as Ca2+, Cu2+, Zn2+, Fe2+, Mg2+ and Hg2+ and incubated under optimized conditions. Enzyme activity was measured as per standard protocol.
Characterization of bacterial isolate
Identification of potential isolates: The isolate was identified after studying morphological, physiological and biochemical characteristics by using Bergey’s Manual (Krieg and Holt, 1984). The identity was further confirmed by 16S rRNA analysis.
Isolation of genomic DNA for 16S rRNA and PCR amplification: Total genomic DNA was extracted from the cells by using phenol-chloroform method and checked for its quality and concentration (Ruzzante et al., 1996). 16S rRNA region was amplified with universal forward and reverse bacterial primers. The PCR amplification was performed using a PTC-150 Mini cycler (MJ Research), with a primary heating step for 2 min at 95°C, followed by 30 cycles of denaturation for 20 sec at 95°C, annealing for 60 sec at 55°C and extension for 2 min at 72°C, then followed by a final extension step for 7 min at 72°C. Each 25 μL reaction mixture contained 2 μL of genomic DNA, 14.25 μL of MilliQ water, 2.5 μL of 10x buffer (100 mM Tris-HCl, pH 8.3; 500 mM KCl), 1.5 μL of MgCl2 (25 mM), 2.5 μL of dNTP’s mixture (dATP, dCTP, dGTP, dTTP at 10 mM conc.), 1.0 μL of each primer (20 pmoles mL-1) and 0.25 μL of Taq DNA polymerase. The PCR-amplified product was analyzed on 1% Agarose gel containing ethidium bromide (0.5 mg mL-1) and 1 kb DNA molecular weight marker and documented using a gel documentation system. The PCR amplicon for the partial 16S rRNA gene was further processed for sequencing. Sequencing was carried out using the same set of primers in both the directions to check the validity of the sequence. Sequencing was done by Ocimum Biosolutions, India.
Antibiotic susceptibility and plasmid curing: The antibiotic susceptibility of the isolate was studied on Muller-Hinton agar media by disc diffusion methods (Bauer et al., 1966). Zone of inhibition was recorded after 24-72 h and interpretation of results was done based on manufacturer’s instructions (HiMedia, India). The antibiotics (mcg) used were Penicillin (10), Ampicillin (10), Chloramphenicol (30), Streptomycin (10), Tetracyclin (30) and Doxycycline (30). Plasmid curing was studied to identify the presence/absence of enzyme production gene on plasmid. It was performed by the method of Trevors (1986).
Statistical analysis: All the experiments were carried out in triplicates and results are mean value of three independent findings. Appropriate controls were maintained in all the experiments. The graphs are prepared by using Microsoft excel.
RESULTS AND DISCUSSION
Psychrotrophic α-amylase producing bacteria: Microbial colonies isolated from soil of Gangotri glacier were screened for cold-active amylase production. Depending upon hydrolysis zone on starch agar media at 15°C, 5 isolates were subjected to enzyme production in broth media. On the basis of maximum enzyme production (3572 units) one potential isolate, designated as GA2, was taken for optimization of media and fermentation conditions (Table 1).
Optimization of cold-active α-amylase production
Effect of incubation time and temperature: The production of α-amylase from GA2 was gradually increases with incubation time and showed maximum yield (4090 units) at 120 h during stationary growth phase after that rapid decline has been observed. Gupta et al. (2008) also reported same time duration for maximum α-amylase production in case of A. niger. Cell mass was also increases up to 72 h after that it attained stationary phase. During stationary phase the enzyme production was increases gradually till 120 h after that it is decreases along with cell mass. The result suggested that enzyme production has direct relationship with cell growth (Fig. 1). In reference to growth phase of the cells, Wijbenga et al. (1991) also reported that Bacillus sp. produced maximum amylase into its late logarithmic growth phase and continued to secrete well into the stationary phase. Temperature has a profound influence on enzyme production and it was found that when cells were incubated at 4 to 50°C in production media, maximum amylase production (4200 units) was obtained at 20°C. There was continuous decline in enzyme production with increase in incubation temperature above 20°C and totally inhibited at 50°C (Fig. 2).
| Table 1: | Production of amylase at 15°C after 48 h incubation |
 | |
| * Values are mean of 3 replications after 2 days incubation at low temp. (15°C) | |
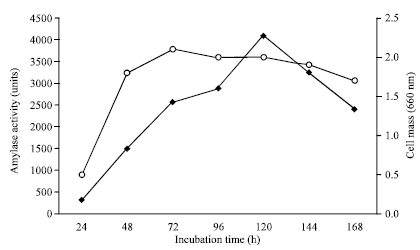 | |
| Fig. 1: | Effect of incubation period on growth (○) and enzyme production (♦) by GA2 |
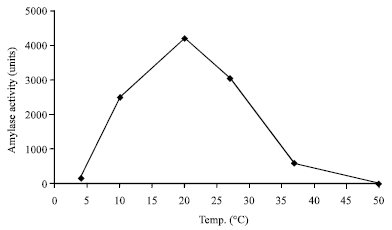 | |
| Fig. 2: | Effect of temperature on enzyme production by GA2 (120 h, pH 7.0) |
According to the classical definition given by Morita (1975) the isolate is psychrotrophic since they have an optimum growth temperature of ~15-20°C but are able to grow up to 30°C.
pH of media and agitation: The pH is known to affect the synthesis and secretion of α-amylase (Fogarty, 1983). The effect of pH on the production of amylase was studied in a pH range of 5-12 and it was found that GA2 produces maximum amylase (4600 units) at pH 9 (Fig. 3). Sudden decline in activity was observed ahead of pH 10. Poornima et al. (2008) also reported the optimum pH of 9.0 for amylase production from Actinomycete strain AE-19. The pH of the culture media strongly affects many enzymatic reactions and transport of compounds across the cell membrane as they are sensitive to the concentration of hydrogen ions present in the medium. pH tolerant α-amylase from microbes are commercially important for detergent industry and can be successfully used as an additive to remove starch based stains. Effective aeration is necessary to support high population of several aerobic bacteria. To check the necessity, isolate was subjected to agitation (120 rpm) and stagnant position at 20°C for 48 h and near about 1.5 fold boosts in enzyme production was noticed in shaking condition as compared to unperturbed condition.
Carbon and nitrogen sources: Carbon and nitrogen sources are necessary for the proper growth and metabolism of microorganisms. The use of cheap C and N sources are important for amylase production as these can significantly reduce the cost. Among the tested C source, GA2 gave maximum yield (5862 units) with lactose followed by maltose (4137 units) while glycerol has an inhibitory effect (Fig. 4). Hamilton et al. (1999) also reported lactose as a superior C-source for amylase production by Bacillus sp. Among the tested N source, GA2 gave maximum yield (5870 units) with yeast extract (Fig. 5). The result was in agreement with Dettori-Campus et al. (1992) and Narayana and Vijayalakshmi (2008) who reported yeast extract as a best N-source for B. stearothermophilus and S. albidoflavus, respectively. In contrast Pedersen and Nielson (2000) reported casein hydrolysate as a best nitrogen source for A. oryzae and Gurudeeban et al. (2011) reported maximum amylase production by B. megaterium in the presence of peptone as nitrogen source.
Influence of heavy metal ions: Heavy metals present in surroundings play an important role in the growth of bacteria. Impact of heavy metals on production of cold-active amylase was evaluated with maximum tolerance level of metals to GA2 at 20±1°C. The enzyme production was enhanced (208%) by Mg2+ (6250 units) in comparison to control (3000 units), whereas Cu2+, Fe2+,Zn2+ and Hg2+ worse the production producing less than 3000 units of enzyme.
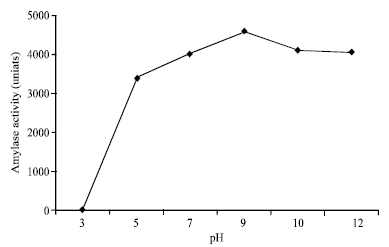 | |
| Fig. 3: | Effect of pH of meida on enzyme production by GA2 (120 h, 20°C) |
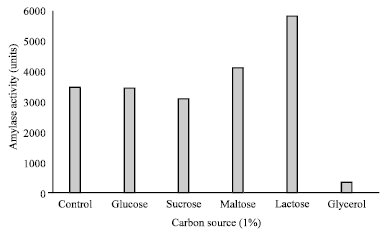 | |
| Fig. 4: | Effect of carbon source on amylase production by GA2 |
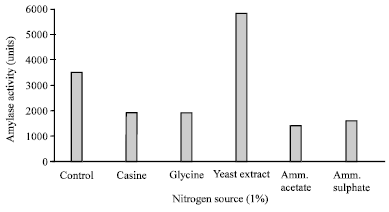 | |
| Fig. 5: | Effect of nitrogen source on amylase production by GA2 |
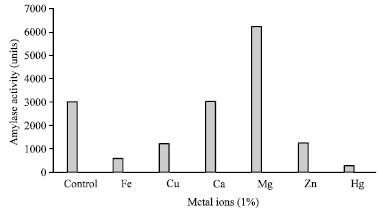 | |
| Fig. 6: | Effect of metal ions on enzyme production by GA2 |
However, Ca2+ (3000 units) have no significant effect (Fig. 6). The fluctuation in enzyme production may be due to either utilization of metals by organism or due to binding of metal ions to the enzyme that may increase or decrease enzyme activity. Vishwanathan and Surlikar (2001 ) also reported increased production of amylase with Ca2+. The result suggested that GA2 produces Ca+2 independent amylase, whose merits are in starch liquefaction, especially in manufacture of fructose syrup, where Ca+2 is a known inhibitor of glucose isomerase as reported by Tonkova (2006).
Characterization of bacterial strain
Identification of bacteria: The isolate GA2 was identified by 16S rRNA sequencing as Microbacterium foliorum and assigned an accession number HQ832574 by NCBI. The major morphological and biochemical characteristics of the isolates are given in Table 2.
Analysis of DNA sequences: Homology of the partial 16S rRNA gene sequence of the isolate was analyzed using the BLAST algorithm in GenBank (http://blast.ncbi.nlm.nih.gov/Blast.cgi). Phylogenetic and molecular evolutionary analyses were conducted using MEGA version 4.1 (Tamura et al., 2007). Only the highest-scored BLAST result was considered for phylotype identification. BLAST showed that the isolate GA2 which is of 1450 bp linear DNA has maximum homology (99%) with Microbacterium foliorum (Fig. 7) with nucleotide base count Adenine: 348, Cytosine: 345, Guanine: 472, Thymine: 285. The DNA sequence was aligned using Clustal W. The forward and reverse sequences of GA2 which we got after sequencing were aligned with the maximum homology sequence of Microbacterium foliorum. The phylogenetic tree was constructed using software MEGA 4.1, this software basically used to estimate the parameters of Genetic variation. A phylogenetic tree based on bacterial 16S rRNA sequence showed a close relationship between the strain GA2 and the genus Microbacterium. For the tree construction four different out groups were used which is mentioned in the Fig. 7 with their accession number. After alignment the final sequence of GA2 was submitted to Genbank at National Centre for Biotechnology Information (NCBI) through Bankit.
| Table 2: | Biochemical characterization of bacteria isolated from Gangotri glacier |
 | |
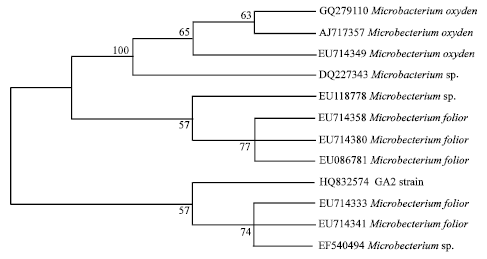 | |
| Fig. 7: | Phylogenetic tree of strain GA2 showing the similarity with M. foliorum |
The sequence in FASTA format is:
Strain GA2 16S ribosomal RNA gene partial sequence (1450 bp) |
Antibiotic susceptibility and plasmid curing: The results of antibiotic susceptibility suggested that Microbacterium foliorum was resistant to penicillin and to be susceptible for all other tested antibiotics. Curing of plasmid was done using curing agent ethidium bromide (20-100 μg mL-1), which gets intercalated between the bases of DNA and inhibits replication of plasmid without inhibiting chromosomal DNA replication. Such inhibition can lead to loss of plasmid. Out of 60 colonies tested on starch agar media, 100% of plasmid curing was obtained as no colony has clear zone of hydrolysis, indicating that amylase production is plasmid mediated characteristic.
CONCLUSION
The results concluded that Microbacterium foliorum GA2 (HQ832574) is a novel psychrotrophic bacterial strain producing cold active α-amylase in alkaline medium, so the enzyme could be successfully applied to remove starchy stains from clothes and used in detergents industry for cold washing that protect the color of fabrics and will be beneficial to save energy as they work at lower temperatures. The strain may also be used in bioremediation of polluted soils and waste waters in cold regions as it is stable over a broad pH range and resistant to various metal ions. The starch-digesting property of this strain at low temperature and its enzyme may replace the conventional synthetic processes in biotechnological industries.
ACKNOWLEDGMENTS
Financial assistance from Council of Science and Technology, Uttar Pradesh, India (Project No. CST/3221) and Technical assistance from Ms. Saima, Department of Biotechnology, Integral University, Lucknow is gratefully acknowledged.
REFERENCES
- Bauer, A.W., W.M.M. Kirby, J.C. Sherris and M. Turck, 1966. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol., 45: 493-496.
CrossRefPubMedDirect Link - Bordbar, A.K., K. Omidiyan and R. Hosseinzadeh, 2005. Study on interaction of α-amylase from Bacillus subtilis with cetyl trimethylammonium bromide. Colloids Surf. B: Biointerfaces, 40: 67-71.
CrossRef - Burhan, A., U. Nisa, C. Gokhan, C. Omer, A. Ashabil and G. Osman, 2003. Enzymatic properties of a novel thermostable, thermophilic, alkaline and chelator resistant amylase from an alkaliphilic Bacillus sp. Isolate ANT-6. Process Biochem., 38: 1397-1403.
CrossRefDirect Link - Cavicchioli, R., K.S. Siddiqui, D. Andrews and K.R. Sowers, 2002. Low-temperature extremophiles and their applications. Curr. Opin. Biotechnol., 13: 253-261.
CrossRefPubMedDirect Link - Dettori-Campus, B.G., F.G. Priest and J.R. Stark, 1992. Hydrolysis of starch granules by the amylase from bacillus stearothermophilus NCA 26. Process Biochem., 27: 17-21.
CrossRef - Georlette, D., V. Blaise, T. Collins, S. D'Amico and E. Gratia et al., 2004. Some like it cold: Biocatalysis at low temperatures. FEMS Microbiol. Rev., 28: 25-42.
CrossRef - Gerday, C., M. Aittaleb, M. Bentahir, J.P. Chessa and P. Claverie et al., 2000. Cold-adapted enzymes: From fundamentals to biotechnology. Trends Biotechnol., 18: 103-107.
CrossRefPubMedDirect Link - Gomes, J. and W. Steiner, 2004. The biocatalytic potential of extremophiles and extremozymes. Food Technol. Biotechnol., 42: 223-235.
Direct Link - Gupta, A., V.K. Gupta, D.R. Modi and L.P. Yadava, 2008. Production and characterization of α-amylase from Aspergillus niger. Biotechnology, 7: 551-556.
CrossRefDirect Link - Haki, G.D. and S.K. Rakshit, 2003. Developments in industrially important thermostable enzymes: A review. Bioresour. Technol., 89: 17-34.
CrossRefPubMedDirect Link - Hamilton, L.M., C.T. Kelly and W.M. Fogarty, 1999. Purification and properties of the raw starch degrading α-amylase of Bacillus sp. IMD 434. Biotechol. Lett., 21: 111-115.
CrossRefDirect Link - Kar, S. and R.C. Ray, 2008. Statistical optimization of α-amylase production by Streptomyces erumpens MTCC7317 cells in calcium alginate beads using response surface methodology. Pol. J. Microbiol., 57: 49-57.
PubMed - Marchi, P., V. Longhi, S. Zangrossi, E. Gaetani, F. Briani and G. Deho, 2007. Autogenous regulation of Escherichia coli polynucleotide phosphorylase during cold acclimation by transcription termination and antitermination. Mol. Genet. Genomics, 78: 75-84.
CrossRefPubMedDirect Link - Mishra, S. and N. Behera, 2008. Amylase activity of a starch degrading bacteria isolated from soil receiving kitchen wastes. Afr. J. Biotechnol., 7: 3326-3331.
Direct Link - Narayana, K.J.P. and M. Vijayalakshmi, 2008. Production of extracellular α-amylase by Streptomyces albidoflavus. Asian J. Biochem., 3: 194-197.
CrossRefDirect Link - Pedersen, H. and J. Nielsen, 2000. The influence of nitrogen sources on alpha amylase productivity of Aspergillus oryzae in continuous culture. Applied Microbiol. Biotechnol., 53: 278-281.
CrossRefDirect Link - Poornima, R., M.K. Sahu, K. Sivakumar and V. Pushpavalli, 2008. Optimization of α-amylase production by actinomycete strain AE-19 isolated from shrimp pond. Trends Applied Sci. Res., 3: 45-52.
CrossRefDirect Link - Ruzzante, D.E., C.T. Taggart and D. Cook, 1996. Spatial and temporal variation in the genetic composition of a larval cod (Gadus morhua) aggregation: Cohort contribution and genetic stability. Can. J. Fish. Aquat. Sci., 53: 2695-2705.
Direct Link - Swain, M.R., S. Kar, G. Padmaja and C.R. Ray, 2006. Partial characterization and optimization of production of extracellular alpha-amylase from Bacillus subtilis isolated from culturable cow dung microflora. Pol. J. Microbiol., 55: 289-296.
PubMedDirect Link - Tamura, K., J. Dudley, M. Nei and S. Kumar, 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol., 24: 1596-1599.
CrossRefPubMedDirect Link - Trevors, J.T., 1986. Plasmid curing in bacteria. FEMS Microbiol. Lett., 32: 149-157.
CrossRefDirect Link - Vishwanathan, P. and N.R. Surlikar, 2001. Production of α-amylase with Aspergillus flavus on Amaranthus grains by solid-state fermentation. J. Basic Microbiol., 41: 57-64.
PubMed - Wijbenga, D.J., G. Beldman, A. Veen and D.L. Binnema, 1991. Production of native-starch-degrading enzymes by a Bacillus firmusflentus strain. Applied Microbiol. Biotechnol., 35: 180-184.
CrossRef - Gurudeeban, S., K. Satyavani and T. Ramanathan, 2011. Production of extra cellular α-amylase using Bacillus megaterium isolated from white mangrove (Avicennia marina). Asian J. Biotechnol., 3: 310-316.
CrossRefDirect Link - Regulapati, R., N.P. Malav and N.S. Gummadi, 2007. Production of Thermostable α-amylases by solid state fermentation: A review. Am. J. Food Technol., 2: 1-11.
CrossRefDirect Link