
M. Dash
Department of Agricultural Biotechnology, Orissa University of Agriculture and Technology, Bhubaneswar, India
P.K. Mishra
Department of Agricultural Biotechnology, Orissa University of Agriculture and Technology, Bhubaneswar, India
D. Mohapatra
Department of Agricultural Biotechnology, Orissa University of Agriculture and Technology, Bhubaneswar, India
Asian Journal of Biotechnology
Year: 2011 | Volume: 3 | Issue: 4 | Page No.: 378-387







ABSTRACT
Mass propagation of sugarcane through meristem culture ensures quick availability of the genetically uniform virus free newly released varieties to the farmers but the occurrence of genetic variation however, limits its commercial application. Isoenzyme analysis may be used to screen somaclones to maintain varietal purity of a sugarcane hybrid like that of a breeder seed in conventionally propagated variety which will be of great benefit to the farmers. Organogenic plant regeneration directly as well as via callus induction from shoot tip of sugarcane indicated that the hormone treatment of KIN (2.0 mg L-1) and NAA (5.0 mg L-1) along with 3% table sugar was best for meristem tip enlargement and shoot initiation. For shoot multiplication a treatment combination of BAP and KIN (1.5 mg L-1 each) was found to be optimum. Highest number of shoots was obtained in treatment combinations of BAP and KIN (1.5 mg L-1 each) along with GA3 (3.0 mg L-1). NAA (5.0 mg L-1) was found optimum with high mean response for rhizogenesis. The detection of genetic variability by analysis of peroxidase isozyme of mericlones and calliclones indicated presence of polymorphism within the mericlones and calliclones. The mericlones exhibited genetic integrity for invertase isozyme while exhibiting the lowest polymorphism for total soluble proteins. The calliclones regenerated after third callus subculture exhibited lowest polymorphism over peroxidase, invertase and total soluble proteins. Thus, micropropagation by callus culture can be advanced up to third subculture to obtain large number of plantlets with minimum variability from that of the parental genotype.
PDF Abstract XML References Citation
Received: February 25, 2011;
Accepted: March 19, 2011;
Published: June 15, 2011
How to cite this article
M. Dash, P.K. Mishra and D. Mohapatra, 2011. Mass Propagation Via Shoot Tip Culture and Detection Of Genetic Variability of Saccharum officinarum Clones using Biochemical Markers. Asian Journal of Biotechnology, 3: 378-387.
DOI: 10.3923/ajbkr.2011.378.387
URL: https://scialert.net/abstract/?doi=ajbkr.2011.378.387
DOI: 10.3923/ajbkr.2011.378.387
URL: https://scialert.net/abstract/?doi=ajbkr.2011.378.387
INTRODUCTION
Sugarcane (Saccharum officinarum) is globally the main source of raw material for the production of sugar. India stands first in area and production among the sugarcane growing countries of the world. To overcome various constraints prevalent in different agro climatic zones of the country commercial use of location specific varieties in sugarcane is important. The newly released varieties take many years to reach the farmers field due to the low seed-multiplication rate ranging from 1:7 to 1:10. Thus, availability of quality seed of newly released varieties is a major constraint in their quick adoption for commercial use. Also there is a need for quality plant material with genetic uniformity of the multiplied plants. Micropropagation through meristem culture is the only methodology which can ensure a faster rate of multiplication to ensure quality seed availability of the new varieties. However, its application is limited due to the occurrence of somaclonal variation (Cavallini et al., 1996). Hence it becomes important to detect the presence or absence of variation in the micropropagated plants.
Biochemical markers like isoenzymes have been used for varietal fingerprinting in sugarcane. Several researchers have revealed variation among the relatives of sugarcane for peroxidases and other isozymes (Heinz and Mee, 1969; Manjunatha et al., 2003; Heinz and Mee, 1971; Hemaprabha et al., 2004). Micropropagation via callus culture is beneficial in terms of production of large number of plantlets of a variety in a short period of time but it is required to find out genetic variation among the regenerated plants. Hence, experiments were conducted to detect the occurance of variability between and within the calliclones. As described by Larkin and Scowcroft (1981), Evans et al. (1984), Karp (1995) and Zucchi et al. (2002) chromosomal variability increases with the increase in time of culture. Keeping this in view analysis using biochemical markers was carried for plants regenerated up to six subcultures. Isoenzyme markers are simple, reliable, faster and comparatively cheaper, so population samples can be analyzed at a reasonable cost and time. Also the micro propagated plants can be screened for their genetic stability at an early stage.
The technique described here entails standardization of shoot tip culture and detection of isozyme variation between mericlones and calliclones. Also it addresses the presence and rate of polymorphism present within the calliclones over a period of time. This will enable to determine the number of subcultures to be done to get the maximum clones with a minimum of genetic variation. Also it will help in screening of genetic variability at the grass root level in the laboratory.
MATERIALS AND METHODS
The tissue culture experiments and the isoenzyme studies were carried out in the Department of Agricultural Biotechnology of College of Agriculture, Orissa University of Agriculture and Technology, India from Sept 2001-2004.
Plant material: The field-collected tops of sugarcane variety Co.87044 (Uttara) (derived from a cross of Co 62198 and CoC 671, having 19.0% sucrose) was washed thoroughly under running tap water and pretreated with carbendazim (0.5%) for 10 min followed by hot water pretreatment for 20 min along with surface sterilization using 70% alcohol for 5 min and HgCl2 (0.1%) for 8 min (Dash et al., 2007).
In vitro culture via direct regeneration and callusing: In order to study the morphogenetic response of sugarcane shoot tip, the most widely accepted MS media was supplemented with 6-benzylaminopurine (BAP), kinetin, naphthalene acetic acid (NAA), 2,4-D and gibberellic acid (GA3) at various concentrations. For direct regeneration the sterilized shoot tips were first inoculated in cell proliferation media and incubated in a light regime of 16 h light and 8 h dark. After shoot initiation they were sub cultured into shoot multiplication medium at due interval by subdividing the growing shoots under aseptic conditions over a period of 6 weeks for multiplication. Subculturing was done for 7-8 cycles of multiplication followed by transferring to rooting media. For callus induction three level of 2, 4-D [dichlorophenoxyacetic acid] were tested at 3% sugar level under light and dark conditions of photoperiod. The tubes were dark incubated for seven days followed by light incubation for shoot regeneration. Full grown callus mass were carefully separated and transferred to the redifferentiation media under normal light conditions. The callus was sub cultured up to 6 generations (R0, R1, R2, R3, R4 and R5). Healthy elongated shoots were separated from the clumps and transferred to rooting media.
Crude enzyme and total soluble protein extraction: The leaf samples (approx.500 mg) were grounded to fine powder with liquid nitrogen and transferred to 1.5 mL eppendroff tubes. Crude enzyme was extracted at 4°C using 0.2 M cold acetete buffer (pH-5.0). Total soluble protein was extracted using 10% TCA (w/v) and 1% 2-ME in cold acetone extraction buffer in 4:1 ratio respectively and incubated for 2 h at –20°C followed by centrifugation at 14,000 g for 15 min with refrigerated high-speed centrifuge (Sigma-3K 15) at 4°C. The pellet was washed in chilled 100% acetone till colourless. The washed and colorless pellet was dried to drive off acetone under vacuum for 5 to 10 min and suspended in 50 mM NaH2PO4 (pH-7.0) buffer. The extracted enzyme and protein was quantified following the procedure of Bradford (1976) and 10 μg of each sample was used for PAGE. Leaves of normal sett grown plants were taken as control. All the experimental materials were taken at 5-6-leaf stage of the plants.
Electrophoresis: The discontinuous Laemmli system (Laemmli, 1970) was used for the present study.
Staining gel: The gels were rinsed with distilled water. This was followed by staining immediately for peroxidase and invertase isoenzymes. The peroxidase staining (50 mL of 1M Sodium acetate, pH-4.6; 100% Methanol, 50 mL; Benzidine, 50 mg; 30% H2O2, 2 mL) was carried out in dark for nearly 30 min till brown color bands were observed. Observations were recorded without delay as the minor active bands diffuse after some time. For invertase staining the gel was incubated for 20 min first in a solution containing 0.1 M sucrose in 0.1 M acetate buffer (pH-4.5) followed by incubating in a second solution containing 0.3 g 2,3,5- TTZ in 200 mL of 1.0 M NaOH in boiling water bath till red bands appear. The red bands disappeared within 3-5 min. The SDS-PAGE gel was stained with 1% Coomassie Blue-R-250 overnight and destained appropriately.
Data analysis: The hormones were tested in three sets with 10 meristems per replication per treatment and analyzed in completely randomized design to study their effect on days to cell proliferation and shoot initiation, average number of explants showing multiple shoots and multiple shoots per explant. Significance of treatment effects was tested using analysis of variance using 2 way test (ANOVA, p≤0.05). The experiment for callus induction was conducted in four replicates with 10 explants per replication. The analysis was done as follows:
Average days to callus induction = ΣNi/4 where, [ΣNi = Total number of days to callus initiation per treatment, i = 4] and Mean response to callus induction as Number of explants inducing calli per treatment/4.
The dataset for the isoenzymes and total soluble protein profiles constituted of nine randomly selected mericlones and calliclones per culture over six subcultures (R0-R5). The experiment was conducted to detect presence or absence of isoenzyme polymorphism among the mericlones and calliclones. All the polymorphic bands were scored for their presence (+) or absence (-) in all the samples from the isozyme profile as discrete character states. The total number of electrophoretic bands (T) was based on all the bands observed over samples and control for each clonal generation. The percent polymorphism was calculated as:
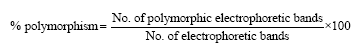 |
RESULTS
Induction of organogenesis and shoot multiplication: The minimum days to tip enlargement was 2.59 and 2.67 days in a combination treatment of 2.0 mg L-1 KIN along with 5.0 mg L-1 NAA without GA3 (Fig. 1). The treatment containing 2.0 mg L-1 KIN, 5.0 mg L-1 NAA and 3.0 mg L-1 GA3 recorded the lowest duration (5.4 days) to shoot initiation. The MS media containing 3% sugar supplemented with BAP and KIN (1.5 mg L-1 each) and GA3 (3.0 mg L-1) resulted in 100% explants exhibiting shooting (Fig. 2). Presence or absence of gibberellic acid in the media had not influenced the regeneration frequency of the explants. The media supplemented with equal dose of BAP and KIN (1.5 mg L-1) in absence of NAA and GA3 produced high number (15.50) of multiple shoots. The average number of effective shoots increased to 15.64 with addition of GA3 in the same media (Fig. 2).
Regeneration via callusing: High dose of 2, 4-D (3.0 mg L-1) induced callus formation within 10 days and showed highest Callus Induction Frequency (CIF) of 95% (Fig. 3) under dark conditions. Callus tissues when transferred to MS media without any hormone supplement turned green faster (within 10 days) and formed green microshoots. The microshoots when transferred to MS media supplemented with growth hormones formed multiple shoots.
Isoenzyme analysis: Polymorphism was observed for peroxidase isozymes among the mericlones and calliclones obtained from six subcultures. Most of the mericlones showed similar peroxidase isozyme pattern as that of the parent. Calliclones regenerated after first subculture (R0) showed a high degree of electrophoretic polymorphism (87.5%) with respect to peroxidase (Table 1). Maximum polymorphism (100%) was observed among the regenerants from fifth (R4) and sixth (R5) callus subcultures. Electrophoretic analysis of mericlones for invertases indicated absence of polymorphism among them but calliclones exhibited polymorphism within the same culture and also between different subcultures.
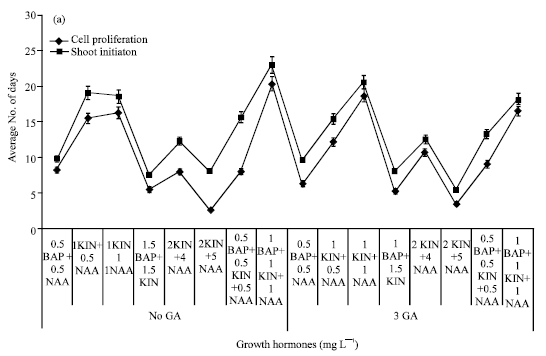 | |
| Fig. 1: | Effect of hormones on sugarcane shoot tips initiation and growth |
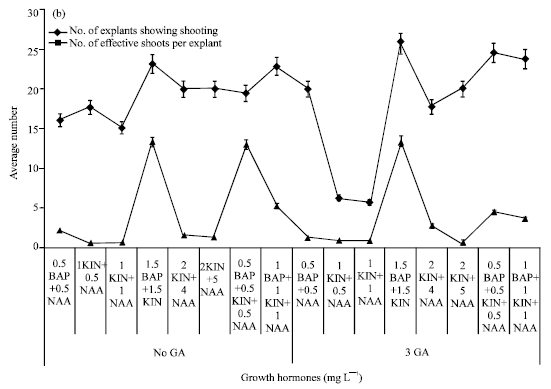 | |
| Fig. 2: | Effect of hormones on multiple shooting of sugarcane |
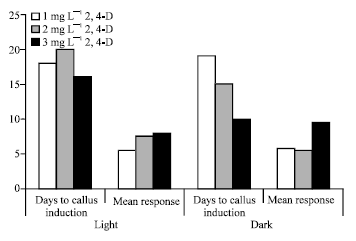 | |
| Fig. 3: | Callus induction in sugarcane shoot tips |
| Table 1: | Isoenzyme and TSP polymorphism of sugarcane mericlones and calliclones obtained at different stages of subculture |
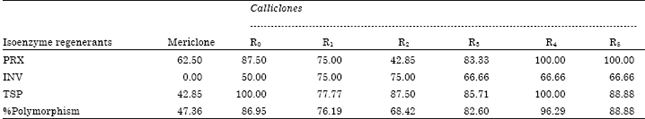 | |
| R0-R5: Callus subcultures, PRX: Peroxidase isoenzyme, INV: Invertase isoenzyme, TSP: Total soluble proteins | |
Lowest polymorphism (50%) was observed in regenerants from first subculture (R0) while maximum polymorphism of 75% was observed in regenerants from second subculture (R1) and third subculture (R2). From total soluble protein analysis by SDS-PAGE it was observed that most of the mericlones were similar with that of the parent exhibiting only 42.85% polymorphism for total soluble proteins. The regenerants from second callus subculture (R1) exhibited the lowest polymorphism (77.77%) followed by the R3 regenerants (85.71%).
DISCUSSION
Organogenesis of sugarcane consists of two critical stages. The first stage is shoot tip enlargement due to cell proliferation which is specifically controlled by endogenous levels of auxins (Murphy and Briske, 1992) and is followed by shoot initiation. The next important stage is shoot multiplication which is again influenced by a number of factors (media composition, temperature, photoperiod etc.). Effect of auxin on vascular tissue differentiation seems to be closely dependent on the presence of sugars (Bhojwani and Razdan, 2004). It is very interesting to note that addition of high concentration of NAA increased the rate of cell proliferation and tip enlargement. The result obtained is in general agreement with those of Hendre et al. (1983), Alam et al. (1995) and Saini et al. (2004). Alam et al. (1995) had obtained quick cell proliferation in presence of 4 mg L-1 NAA along with 0.5-1.0 mg L-1BA while, Hendre et al. (1983) observed shoot elongation in MS media fortified with 0.01 mg L-1 IBA and 0.1 mg L-1 GA3.
The growing apical portion produces endogenous auxins (IAA) which stimulate cell enlargement and development of the growing point. Growth promotive concentrations of auxin have been found to stimulate the synthesis of both DNA and RNA within the nuclei. This in turn controls various metabolic pathways. The sugars are responsible for greater osmotic potential and increased respiration leading to more synthesis of cell wall material i.e., cellulose and lipid. These events lead to cell elongation. Auxin on the other hand causes a preferential movement of nutrients, cytokinins and other growth promoters towards the shoot tip. This reduces their availability to the lateral buds inhibiting their further growth. The lateral buds are more sensitive to higher concentrations of auxins than the apical buds. This inhibition of lateral buds to grow by the presence of apical meristem is known as apical dominance. Presence of apical dominance is an inherent characteristic of all crops and is predominant in sugarcane. When the growing tip is excised for use as explant the cells get disturbed and reduces auxin production. So external application of auxins (NAA) stimulates cell enlargement and helps in cell proliferation and tip enlargement (Morris et al., 2001; Booker et al., 2003). Quick shoot initiation in presence of GA3, NAA and KIN may be primarily due to the mode of action of the hormones. The effect of gibberellic acid depends on availability of auxin and conversely auxin induced growth depends on gibberellin (Ralph, 1970). Gibberellin induced growth is mediated by changes in the microtubules and increase in template activity of DNA, followed by RNA polymerase activity and synthesis of RNA and proteins. Thus gibberellic acid acts as gene derepressors and activates some segments of DNA (Bhatia and Parasher, 2003). Cytokinins are an important regulatory factor of plant meristem activity and morphogenesis, with opposing roles in shoots and roots. Growing on cytokinin rich media can reduce apical dominance leading to more numbers of shoots (Kozlowski, 1964). High cytokinin concentration is required for meristem culture t o induce shoot multiplication. The above results are at par with Lal (1992, 1993), Shukla et al. (1994) and Chattha et al. (2001) for sugarcane. Dhumale et al. (1994) observed that percentage induction of multiple shoot and mean multiple shoot number were highest (90% and 16.5, respectively) with 3.0 mg L-1 BAP+1.0 mg L-1 IAA. Effect of growth regulators on Shoot-Tip culture has been demonstrated in Papaya (Kabir et al., 2007).
Micropropagation via callus culture is known to produce large number of shoots as compared to direct regeneration in the same time period. Auxins applied extrinsically seem to be capable of erasing the genetically programmed physiology of plant tissues, thereby reverting them to a dedifferentiated state. It has been suggested that auxins cause changes in DNA methylation pattern, leading to reprogramming of differentiated cells. Absence of light creates a stress environment and helps in dedifferentiation of the organized tissues to form unorganized callus tissues. Auxin destruction occurs by an oxidase system called IAA-oxidase in absence of light. Oxidation of IAA consumes oxygen and liberates carbondioxide. This in turns leads to release of monophenols which promote IAA-oxidase activity resulting in very low endogenous auxins and no organ formation (Bhatia and Parasher, 2003). High amount of auxin (3.0 mg L-1 2, 4-D) results in somatic embryogenesis (Gamborg and Phillips, 1995) but for further growth high auxin content is inhibitory and a hormone free medium induces development of somatic embryos which turned into plantlets. Growth in absence of hormones can be attributed to the growth promoting effect of hydrolyzed agar (Englivd, 1964). The results are in agreement with those of Mohatkar et al. (1993), Nand and Singh (1994) while, Kale et al. (2004) obtained quick callus initiation in 1.0 mg L-1of 2,4-D under dark conditions. Use of 3.0 mg L-1 2, 4-D for highest percentage of callus induction was observed by Karim et al. (2002). Niaz and Azra (2002) reported that Medium containing BA @1.0 mg L-1 performed better than Kinetin with the highest percentage of shoot induction. Similar results for shoot tip culture was also reported by Roy and Kabir (2007).
Several researchers have revealed variation among the relatives of sugarcane for peroxidases and other isozymes (Heinz and Mee, 1969; Manjunatha et al., 2003). Liu and Chen (1978) for esterases and Nagai et al. (1991) have conducted isoenzyme analysis to detect and confirm genetic variation among sugarcane somaclones developed via callus cultures. Hemaprabha et al. (2004) used peroxidase, esterase and phosphorylase isoenzyme analysis to detect the relationship between parental clones and progeny performance in sugarcane. Studies to detect genetic variation or stability using isoenzymes has also been reported in date coconut (Geethalakshmi et al., 2005) and palm (Zivdar et al., 2008), respectively.
The electrophoretic gel analysis for peroxidase indicated increase in the number of polymorphic bands over different subcultures and also within subcultures (Table1). This indicated that the meristematic tissues were subjected to high stress environment during in vitro culture. The low degree of polymorphism among the clones obtained from third callus subculture (R2) indicate that most of the genetic instability was recovered during redifferentiation of the callus tissues after two subcultures. This may be due to chromosomal rearrangement and convergence of the variant genes which had emerged during two subcultures (Brewer, 1970).
The invertase group of sugar catabolising enzymes that control ripening in sugarcane; high level of acid invertases (in vacuole) and low level of neutral invertase (in cytoplasm) are associated with rapid vegetative growth whereas the reverse pattern is associated with ripening (Mamat and Galwey, 1999). The hydrolysis of sugars by this enzyme is the principal mechanism by which hexose sugars are released for the metabolism in tissues undergoing rapid cell expansion (Alaoui-Sosse et al., 1996). The activity of invertases is high in early stages of development and decreases with maturation. Thus study of invertase isozymes can be useful to detect variation in the micropropagated sugarcane plantlets at an early stage of development. Hence, selection of clones for invertase stability at the lab level will probably lead to less variation for sucrose content at the field level. Most of the regenerants from third subculture (R2) were 100% similar and the two variants were more than 85% similar with the parent.
| Table 2: | A scheme for easy and quick detection of in vitro generated genetic variation |
 | |
The above findings point to the fact that there may be variation in final sucrose concentration of the mature stems obtained from calliclones while the mericlones and the R2 regenerants may not show any variation in final sucrose concentration of the mature stems.
High polymorphism among the calliclones and mericlones for total soluble protein may be due to changes in sequential expression of various genes controlling cell cycle and production of special type of proteins (Sethi and Mukherjee, 1990). High amount of growth regulators especially auxin used in the culture media induced stress leading to under or over expression of the genes. This must have resulted in production of new proteins and suppressed production of other proteins (Kalra and Bhatia, 1996). Mericlones exhibited polymorphism with respect to peroxidase (66%) and total soluble proteins (42.85%) electrophoretic patterns but did not show any polymorphism for invertases. On comparing the calliclones with the mericlones, it was observed that the trend of increase in genetic instability over the three characters was as follows: Mericlones<R2<R1<R3<R0<R5<R4. Over the subcultures the trend observed was INV<PRX<TSP. It was observed that R2 clones exhibited low degree of polymorphism as compared to other calliclones. Decrease in percent polymorphism in R2 may be due to convergence of the variant genes which had emerged during two subcultures. The high percent polymorphism for total soluble proteins (86.11%) among the clones and within the clones indicates that lot of change is occurring in most or all of the genes coding for different leaf proteins during callus phase. The changes may be in form of DNA methylation, addition or deletion of base pairs during callus tissue redifferentiation which result in a transcription for a different polypeptide.
CONCLUSION
Micropropagation by callus culture can be advanced up to third subculture to obtain large number of plantlets with minimum variability from that of the parental genotype. The intrinsic variability present in the sugarcane clones and the in vitro generated variability can be easily detected by use of isozyme markers and total protein markers (Table 2). This analysis can be done at the regeneration stage of mass propagation at the grass root level to rough out the off types before planting the clones in the field. Thus the present technique of shoot tip culture may be used to maintain varietal purity of a sugarcane hybrid like that of a breeder seed in conventionally propagated variety which will be of great benefit to the farmers.
REFERENCES
- Alaoui-Sosse, B., S. Ricaud, P. Barnola and P. Dizengremel, 1996. Rhythmic growth and carbon allocation in Quercus robur. Sucrose metabolizing enzymes in leaves. Physiol. Plant., 96: 667-673.
CrossRef - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Cavallini, A., L. Natali, E. Polizzl and T. Giordani, 1996. Variation of repetitive DNA sequences in progenies of regenerated plants of Pisum sativum. J. Heredity., 87: 233-237.
Direct Link - Englivd, K.C., 1964. Growth and chlorophyll formation of dark-grown pine embryos on different media. Physiol. Plant., 17: 866-874.
CrossRef - Evans, D.A., W.R. Sharp and H.P. Medina-Filho, 1984. Somaclonal and gametoclonal variation. Am. J. Bot., 71: 759-774.
Direct Link - Heinz, D.J. and G.W.P. Mee, 1971. Morphologic, cytogenetic and enzymatic variation in saccharum species hybrid clones derived from callus tissue. Am. J. Bot., 58: 257-262.
Direct Link - Murphy, J.S. and D.D. Briske, 1992. Regulation of tillering by apical dominance: Chronology, interpretive value and current perspectives. J. Range Manage., 45: 419-429.
Direct Link - Booker, J., S. Chatfield and O. Leyser, 2003. Auxin acts in xylem-associated or medullary cells to mediate apical dominance. Plant Cell., 15: 495-507.
CrossRef - Kalra, G. and S.C. Bhatia, 1996. Auxin-Ethylene interaction in adventitious rooting and related changes in anodic peroxidase isozymes in sunflower hypocotyls. J. Plant Biochem. Biotech., 5: 37-42.
Direct Link - Karp, A., 1995. Somaclonal variation as a tool for crop improvement. Euphytica, 85: 295-302.
CrossRefDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Larkin, P.J. and W.R. Scowcroft, 1981. Somaclonal variation-a novel source of variability from cell cultures for plant improvement. Theor. Applied Genet., 60: 197-214.
CrossRefDirect Link - Mamat, L.D. and N.W. Galwey, 1999. A relationship between stalk elongation and earliness of ripening in sugarcane. Exp. Agric., 35: 283-291.
CrossRefDirect Link - Zucchi, M.I., H. Arizono, V.A. Morais, M.H.P. Fungaro and M.L.C. Vieira, 2002. Genetic instability of sugarcane plants derived from meristem cultures. Genet. Biol., 25: 91-96.
Direct Link - Morris, G.C., S.P. Chatfield and O. Leyser, 2001. NAA restores apical dominance in the axr3-1 mutant of Arabidopsis thaliana. Ann. Bot., 87: 61-65.
CrossRef - Ralph, O., 1970. The dependence of auxin-induced pea stem growth on gibberellin. Bot. Gazette, 131: 95-97.
Direct Link - Geethalakshmi, P., V.A. Parthasarathy and V. Niral, 2005. Genetic diversity among coconut (Cocos nucifera L.) genotypes using isozymes. Asian J. Plant Sci., 4: 678-683.
CrossRefDirect Link - Kabir, A.H., M.A. Bari, A.K.M.N. Huda, M.A. Rezvy and I. Mahfuz, 2007. Effect of growth regulators and carbon sources on axillary shoot proliferation from shoot-tip explant and successful transplantation of papaya (Carica papaya L.). Biotechnology, 6: 268-272.
CrossRefDirect Link - Karim, M.Z., M.N. Amin, M.A. Hossain, S. Islam, F. Hossin and R. Alam, 2002. Micropropagation of two sugarcane (Saccharum officinarum) varieties from callus culture. J. Biol. Sci., 2: 682-685.
CrossRefDirect Link - Lal, N., 1993. Proliferation and greening effects of sucrose on sugarcane shoot cultures in vitro. Sugarcane, 6: 13-15.
Direct Link - Liu, M.C. and W.H. Chen, 1978. Tissue and cell cultures as aids to sugarcane breeding II. Performance and yield potential of callus derived lines. Euphytica, 27: 273-282.
CrossRef - Manjunatha, B.R., S. Virupakshi and G.R. Naik, 2003. Peroxidase isozyme polymorphism in popular sugarcane cultivars. Curr. Sci., 85: 1347-1349.
Direct Link - Nagai, C., B.S. Ahloowalia and T.L. Tew, 1991. Somaclonal variants from an intergeneric hybrid Saccharum spp. hybrid X E. arundinaceum. Euphytica, 53: 193-199.
CrossRef - Niaz, F. and Q. Azra, 2002. Effect of growth regulators on the regeneration potential of two sugarcane cultivars SPF-213 and CPF-237. Pak. J. Biol. Sci., 5: 1081-1083.
CrossRefDirect Link - Roy, P.K. and M.H. Kabir, 2007. In vitro mass propagation of sugarcane (Saccharum officinarum L.) var. Isd 32 through shoot tips and folded leaves culture. Biotechnology, 6: 588-592.
CrossRefDirect Link - Saini, N., M.L. Saini and R.K. Jain, 2004. Large scale production, field performance and RAPD analysis of micropropagated sugarcane plants. Ind. J. Genet., 64: 102-107.
Direct Link - Zivdar, S., M. Mousawi and N. Alemzadeh Ansari, 2008. Genetic stability in date palm micropropagation. Asian J. Plant Sci., 7: 775-778.
CrossRefDirect Link