
Priyanka Siwach
Department of Biotechnology, Chaudhary Devi Lal University, Sirsa, Haryana, India
Kiran Grover
Department of Biotechnology, Chaudhary Devi Lal University, Sirsa, Haryana, India
Anita Rani Gill
Department of Biotechnology, Chaudhary Devi Lal University, Sirsa, Haryana, India
Asian Journal of Biotechnology
Year: 2011 | Volume: 3 | Issue: 3 | Page No.: 280-292







ABSTRACT
Several factors, affecting the induction and proliferation of callus cultures of Thevetia peruviana Schum.were investigated in this study, including explants nature, the phyto-hormones (nature, concentration and combination) and the sucrose concentration. Nodal segments responded with higher frequency of callus induction and proliferation, as compared to inter-nodal segments and shoot apices, on MS medium supplemented with various concentration of 2,4-D and kinetin, individually. Among twenty five different growth regulators formulations supplemented to Murashige and Skoog’s medium, nodal segments responded with a maximum of 100% frequency of callus induction on a combination of 9.05 μM 2,4-D with 0.93 μM kinetin, followed by frequency of 88.3% on 6.97 μM Kinetin, supplemented alone. A very low frequency of callus induction, for all the three explants, was observed on media supplemented with different concentrations of 2,4-D alone. Maximum callus proliferation, resulting in fresh weight of 2.556 g, after one month of culture, was observed for callus initiated from nodal segments, on MS medium supplemented with 9.05 μM 2,4-D and 0.93 μM kinetin (the same medium giving highest frequency of callus induction). Of the three levels, sucrose at 3% was found as the most favorable level for callus initiation and proliferation. Callus could be successfully maintained in healthy and proliferative form for consecutive nine months on the MS medium supplemented with 9.05 μM 2,4-D and 0.93 μM kinetin.
PDF Abstract XML References Citation
Received: November 06, 2010;
Accepted: January 12, 2011;
Published: February 07, 2011
How to cite this article
Priyanka Siwach, Kiran Grover and Anita Rani Gill, 2011. The Influence of Plant Growth Regulators, Explant Nature and Sucrose Concentration on in vitro Callus Growth of Thevetia peruviana Schum. Asian Journal of Biotechnology, 3: 280-292.
DOI: 10.3923/ajbkr.2011.280.292
URL: https://scialert.net/abstract/?doi=ajbkr.2011.280.292
DOI: 10.3923/ajbkr.2011.280.292
URL: https://scialert.net/abstract/?doi=ajbkr.2011.280.292
INTRODUCTION
Thevetia peruviana (Pers.) K. Schum. is a small ornamental shrub belonging to the family Apocyanaceae. It is commonly called as Be still tree, Digoxin, Lucky nut, Nerium Oleander, Yellow oleander and milk bush. It is an evergreen, dicotyledonous shrub, with origin from the forest of tropical America and now widely spread out in various tropical and subtropical regions of the world (Pearn, 1989). It grows to a height of about 10 to 15 feet and continues its blooming throughout the year. Flowers are bright yellow and funnel shaped with 5 petals spirally twisted. The fruits are green in color and changes from green to brownish black when it achieves maturity. Each fruit contains a nut which is longitudinally and transversely divided. Propagation is primarily by means of seeds, can also propagate from cuttings in spring early summer with hardwoods cuttings (Prajapati et al., 2003).
Besides being a very popular ornamental tree, the medical and toxicological properties of this plant have also been recognized. It has been used, for example, in the treatment of hemorrhoids, ulcers, leprosy, snake bites, in the induction of abortion and also reported to have anti-cancerous properties. The whole plant exudes milky juice which is very poisonous, quite unsafe for internal use. It has also been regarded as a potential source of biologically active compounds, namely insecticides, rodenticides, fungicides and bactericides (Patil et al., 2007). Its seeds, leaves, fruits and roots are being used in traditional medicine as an acrid purgative, as emetic and for intermittent fever treatment (Gata-Gonclaves et al., 2003). In recent years the plant has been reported to have strong anti-HIV activity also (Tewtrakul et al., 2002). Such diverse medicinal applications of the plant clearly indicate towards presence of diverse secondary metabolites in various parts. Many secondary metabolites like cardiac glycosides thevetin A and B, thevetoxin, peruvoside, iridoid glycosides, cerberin, perusitin, peruvoside, quercetin, flavonoids, triterpenes, monoterpenes, thevefolin, ruvoside, digitonin, peruvianoside kokiphin, theviridoside, theveside, nerifolin and oleandrin have been reported in different extracts of the plant (Abe et al., 1996). Oleandrin is reported to be a potent inhibitor of human tumor cell growth (Afaq et al., 2004). So there is a sharp rise in the commercial demand of Thevetia peruviana in the pharmaceutical industry, during recent years. Naturally occurring plants are unable to meet this demand because of many problems like non-availability of large scale true-to type planting material, natural propagation being vulnerable to environmental changes and the non applicability of genetic improvement techniques. These problems can be easily dealt with plant tissue culture techniques.
Plant cell cultures were introduced as an important tool for studying and producing plant secondary metabolites in the mid 1960s. Improved cell and tissue culture technologies would help in producing the active compounds in vitro with better productivities without cutting down the natural resources. Some highly effective secondary metabolites that are used in pharmaceuticals, food industry and cosmetics have been produced through plant cell cultures, callus cultures, shoot cultures and root cultures (Ramachadra-Rao and Ravishankar, 2002; Zhang et al., 2002; Mathur et al., 2007; Taha et al., 2008). The direct manipulation of plant cell and tissue culture systems has many advantages over the conventional isolation of secondary metabolites (Kim et al., 1995). Such manipulations have been reported to enhance the secondary metabolites production in many species (Zhang et al., 2002; Mathur et al., 2007).
Till now, there is no report regarding the use of callus cultures of Thevetia peruviana for secondary metabolites isolation. To the best of our knowledge, there are only a few reports about the tissue culture work on Thevetia peruviana. The available ones report the callus induction and plant regeneration using leaf discs as the explant (Kumar, 1992; Zibbu and Batra, 2010). No studies are available regarding the use of stem segments for callus or shoot regeneration. In the present study we report a highly efficient protocol for callus induction from stem segments of Thevetia peruviana and its subsequent maintenance for a period of consecutive nine months for further use in regeneration studies and biochemical analysis.
MATERIALS AND METHODS
Plant material and explants preparation: The plant material used in the present investigation was collected from 6-7 year-old shrub of Thevetia peruviana, growing near the science block of the Chaudhary Devi Lal University, Sirsa, Haryana, India, during the months of January to July, 2009. Three explants viz., nodal segments, inter-nodal segments and shoot apices (each of 3-4 cm in size) were excised from healthy and juvenile shoots and were washed under running tap water for 10 min to remove the traces of dust and dirt. These were then swabbed with an alcohol (50% v/v) soaked muslin cloth followed by washing in tween-20 (2% v/v) for 10 min. After it, these were subjected to surface sterilization by treating with 0.1% (w/v) mercuric chloride solution for 8 min. Subsequently, explants were thoroughly rinsed 5 times with sterilized distilled water. The explants were trimmed to a final size of 0.5-2 cm and were placed aseptically in 150 mL Erlenmeyer flasks/Magenta boxes containing 35-40 mL medium.
Callus induction
Effect of various concentrations of 2,4-D and kinetin, individually: All the three types of explants were cultured vertically on MS (Murashige and Skoog, 1962) medium supplemented with five different levels of 2,4-D (2.26, 4.52, 6.79, 9.05, 11.31 and 13.57 μM) and kinetin (2.32,4.65, 6.97, 9.29 and 11.61 μM), individually, to select out the best explant and most favoring growth regulator regime with regard to callus induction and proliferation.
The medium was fortified with 3% (w/v) sucrose and pH was adjusted to 5.8 before being solidified with 0.8% (w/v) agar. The culture vessels containing the media were autoclaved at 15 lb and 121°C for 20 min. Cultures were maintained at 25±2°C, at a photoperiod of 16 h (80 μmol m-2 sec-1) for 30 days. On 30th day, observations were made for % callus induction, fresh weight of callus and callus morphology. Though dry weight is more reliable criteria to assess the proliferation, we needed to grow more amount of callus tissue for subsequent biochemical investigation (unpublished data). So fresh weight recordings were done by placing the weighing balance in Laminar air flow chamber, under strict aseptic conditions and the cultures were then used for further sub-culturing experiments. Observations were made daily for noting the number of days required for callus initiation.
Effect of various combinations of 2,4-D and kinetin: The best responding explant, as observed in the above study, was subjected to fifteen different combinations of 2,4-D and kinetin (five levels of 2,4-D viz., 2.26, 4.52, 6.79, 9.05, 11.31 and 13.57 μM, with three levels of kinetin viz., 0.46, 0.93 and 1.39 μM, in 5x3 factorial combinations) and observations were made as reported eariler. Medium sterilization and culture conditions were as described earlier.
Effect of sucrose concentration: Retaining the best explant and two most favoring growth regulator regimes with regard to % callus induction and fresh weight of callus, two more levels of sucrose (2 and 2.5%) were tested and observations were made as reported earlier. Medium sterilization and culture conditions were as described earlier.
Callus maintenance: Calli, initiated from nodal segments were selected for long term growth because of maximum proliferation as observed after one month of culture. Out of twenty five media (having different concentration and combinations of growth regulators) used for callus induction study, two were selected for callus maintenance study because of high fresh weight of callus observed on them after one month of culture.
Calli were routinely sub cultured for consecutive eight months, each sub culture after a regular interval of 30 days, on the two selected media formulation. Calli pieces weighing 20-25 mg (fresh weight), obtained by division of larger callus, were inoculated on fresh medium and allowed to proliferate for 30 days. On 30th day, fresh weight increase (fwi) was calculated as the difference between the final and the initial fresh weight. Fresh weight recording was done under aseptic conditions.
Experimental design and statistical analysis: All the experiments were conducted using a Complete Random Design (CRD). There were 10 replicates per treatment and each treatment was repeated in three sets. Data were subjected to analysis of variance (ANOVA) using the SPSS version 12. The significant differences among the treatments were compared using Duncan’s Multiple Range Tests (DMRT). Treatment values were represented as the Mean±SE.
RESULTS
Callus induction
Effect of various concentrations of 2,4-D on the three explants: Inter-nodal segments and shoot apices did not respond on any of the five concentrations of 2,4-D. The two way ANOVA for the differences among various concentrations of 2,4-D and among experimentally replicated sets with regard to % callus induction, number of days for callus induction and fresh weight of callus indicated that all the three responses from nodal segments were significantly affected by concentration of 2,4-D (p<0.01).
The various concentrations of 2,4-D used in this study did not favor the callus induction to the optimum level. The nodal segments responded with the maximum frequency of only 20% on 6.79 μM 2,4-D, significantly not different to the frequency of 18.33% observed on 4.52 μM level (Table 1). Frequency of callus induction was quite low (10%) on 2.26, 9.05 and 11.21 μM concentration of 2,4-D. Callus initiation from the nodal segments was observed from the lower cut portion, after a minimum of 19.636 days and 20.591 days on 6.79 μM and 9.05 μM 2,4-D, respectively significantly earlier than that on other concentrations (Table 1). Maximum proliferation resulting in significantly higher fresh weight, of 0.4064 g, callus with greenish white color, was observed after thirty days of culture on 6.79 μM 2,4-D (Fig. 1a), while on other concentrations of 2,4-D significantly lower proliferation was observed, as interpreted by DMRT (Table 1).
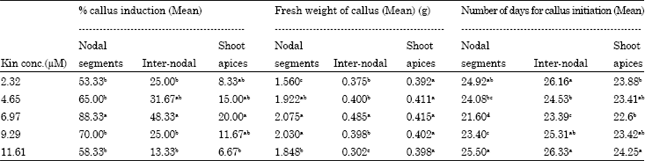
Effect of various concentrations of kinetin on the three explants: The three-way ANOVA indicated that the three responses were significantly affected by the concentration of kinetin (p<0.01) as well as the nature of explant (p<0.01). At each level of kinetin, nodal segments responded with high frequency of callus induction followed by inter-nodal segments and shoot apices (Table 2).
For nodal explants, maximum frequency of callus induction (88.33%) was observed on 6.97 μM kinetin, significantly higher from that on other concentrations, as analyzed by DMRT (Table 2). Kinetin, at 6.97 μM, was also observed as the most productive level for inter-nodal and shoots apices, in terms of maximum % callus induction (Table 2). The mean number of days required for callus initiation from nodal segments was significantly low (22.60 days) on 6.97μM kin, amongst all the five concentrations.
| Table 1: | Percent callus induction, fresh weight of callus, after thirty days of culture and number of days for callus initiation recorded from nodal segments on various concentrations of 2,4-D |
 | |
| Means within columns followed by the same letters are not significantly (p<0.05) different | |
 | |
| Fig. 1: | Callus, initiated from nodal segments, after 30 days of culture on MS medium supplemented with (a) 6.79 μM 2,4-D, (b) 6.79 μM kin and (c) 9.05 μM 2,4-D + 0.93 μM kin |
| Table 2: | Effect of various concentrations of kinetin, on the three explants, with regard to frequency of callus induction, fresh weight of callus, after 30 days of culture and number of days for callus initiation |
 | |
| Means within columns followed by the same letters are not significantly (p<0.05) different | |
On higher and lower concentration than 6.97 μM kin, the number of days for callus initiation was significantly higher than that observed on it. Same situation was observed for inter-nodal segments and shoot apices. Callus was initiated from the lower cut portion, touching the medium, of each of the three explants and was light greenish in color. Callus, initiated from nodal segments, proliferated more as compared to the callus from inter-nodal and shoot apices at each level of kinetin, as indicated by fresh weight recordings (Table 2). For nodal segments, maximum proliferation resulting in 2.075 g of callus was observed on 6.97 μM kinetin (Fig. 1b) and this was significantly not different to fresh weight of 2.03 g observed on 9.29 μM kinetin (Table 2). Inter-nodal and shoot apices initiated calli proliferated to a maximum of only 0.485 and 0.414 g, respectively both on 6.97 μM kinetin, after one month of culture.
Effect of different combinations of 2,4-D and kinetin: Since in the above two studies, nodal segments responded far better than inter-nodal and shoot apices (both in terms of frequency of callus induction and proliferation), only nodal segments were subjected to different combinations of 2,4-D and Kinetin. Two-way ANOVA indicated that the response of nodal segments was significantly affected by the combination type (p<0.01).Among fifteen combinations, maximum of 100% callus induction was observed on medium supplemented with a combination of 9.05 μM 2,4-D and 0.93 μM kinetin (Fig. 2). For each of the three levels of kinetin, frequency of callus induction was observed maximum with 9.05 μM 2,4-D (Fig. 2). Significantly lower frequency was observed on combinations having 2.26 μM 2,4-D (lowest concentration used) with each of the three concentration of kinetin. Quite similar frequencies of callus induction were observed on media having quite dissimilar ratio of 2,4-D to kinetin (Fig. 2).
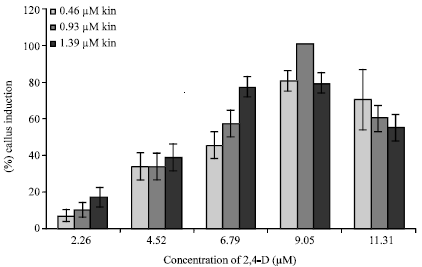 | |
| Fig. 2: | Frequency of callus induction from nodal segments on various combinations of 2,4-D and kinetin, as observed after 30 days of culture (Bars on the columns represent the standard error) |
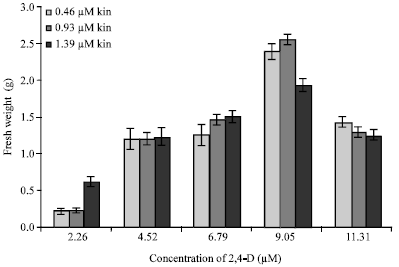 | |
| Fig. 3: | Fresh weight of callus (g), initiated from nodal segments, on various combinations of 2,4-D and kinetin, as observed after 30 days of culture (Bars on the columns represent the standard error) |
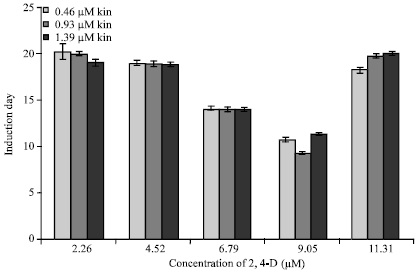 | |
| Fig. 4: | Number of days for callus initiation from nodal segments on different combinations of 2,4-D and kinetin (Bars on the columns represent the standard error) |
A similar trend was also observed for callus fresh weight increase, observed after one month of culture (Fig. 3). Significantly higher fresh weight callus mass of 2.556 g, of light green color, was observed after one month of culture on combination of 9.05 μM 2,4-D with 0.93 μM kinetin (Fig. 1c). Callus was observed to be initiated earliest (9.283 days) on combination of 9.05 μM 2,4-D with 0.93 μM kinetin, while in general the number of days required for callus initiation was significantly lower on 9.05 μM 2,4-D, with each of the three levels of kinetin, compared to other combinations (Fig. 4).
Effect of sucrose concentration: Retaining the best explant (nodal segments) and two media formulation (one with 6.97 μM kinetin alone and other with 9.05 μM 2,4-D in combination with 0.93 μM kinetin), with regard to maximum frequency of callus initiation and subsequent proliferation as reported in the previous sections, two lower levels of sucrose (2.5 and 2%) were used for studying the effect on the above said responses (Table 3). The aim was to know whether at lower level we could achieve considerably similar/higher %callus induction with same or reasonably higher proliferation.
When the responses on 2.5 and 2% sucrose were compared to that on control (3%) for both the medium formulations, the %callus induction did not differ significantly on 2.5% sucrose while on 2% level, it was significantly lower. On the other hand, fresh weight of callus (after four weeks of culture) was found to be significantly different on all the three levels of sucrose, highest being for 3%. On 2% sucrose, callus proliferation was lowest and the callus obtained was white in color with friable appearance. Day for callus initiation was not affected significantly by the change in sucrose concentration (Table 3).
Callus maintenance: Calli, obtained from nodal segments were maintained for consecutively nine months, on the two medium formulations (one with 6.97 μM kinetin alone and other with 9.05 μM 2,4-D in combination with 0.93 μM kinetin), each with 3% sucrose (as it was observed to be most optimal concentration for callus proliferation), to define the best conditions for long term maintenance and proliferation of callus cultures of T. peruviana (Table 4).
| Table 3: | Percent callus induction, fresh weight of callus, after thirty days of culture and number of days for callus initiation on different levels of sucrose |
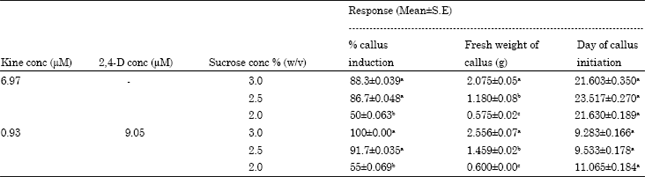 | |
| Means within columns followed by the same letters are not significantly (p<0.05) different | |
| Table 4: | Fresh weight (g) of callus as observed on 30th day of each sub-culturing, on different media formulations |
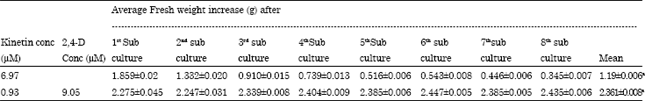 | |
| Means within columns followed by the same letters are not significantly (p<0.05) different | |
 | |
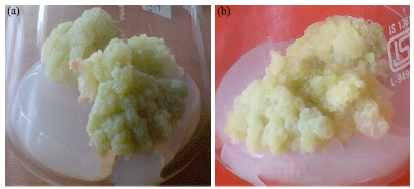
| Fig. 5: | Callus proliferation on MS medium supplemented with 6.97 μM kinetin after (a) Fifth sub-culture and (b) Eight sub-culture |
 | |
| Fig. 6: | Callus proliferation on MS medium supplemented with 9.05 μM 2,4-D+0.93 μM kinetin after (a) Fifth sub-culture and (b) Eight sub-culture |
It was observed that calli, maintained on MS medium supplemented with 0.93 μM kinetin along with 9.05 μM 2,4-D, proliferated with an average monthly fresh weight of 2.361 g, while on other medium (MS supplemented with 6.97 μM kinetin alone) the average monthly fresh weight of callus was observed to be significantly low, being 1.19 g (Table 4). Also, on 6.97 μM kinetin, the fresh weight of callus decreased gradually after each sub-culture, while on other medium (0.93 μM kin+9.05 μM 2,4-D) a nearly consistent value of fresh weight of callus was observed after each subculture (Table 4). Callus morphology was also found to be influenced by nature of growth regulators. Calli, on kinetin supplemented media, became whiter in color with each sub-culture (Fig. 5a, b). On the other hand, calli, sub-cultured on medium having both 2,4-D and kinetin were light green in color till the last subculture (Fig. 6a, b).
DISCUSSION
During the present investigation, nodal segments responded with higher frequency of callus induction and proliferation on different concentrations of 2,4-D and kinetin, used individually, compared to shoot apices and inter-nodal segments. Variation in the callus forming ability of different explants has been reported in many other plants also (Ishii et al., 1998; Zouine and Hadrami, 2004). Some have explained this callogenesis specificity of explants by their differential reactivity to media components (Zouzou et al., 1997; Ikram-ul-Haq, 2005).
The plant growth regulators play a key role in directing the organogenic response of any plant tissue/organ under in vitro conditions (Che et al., 2002; Sugiyama and Imamura, 2006). Auxins, mainly 2,4-D, have been reported to be as the most effective growth regulators for the induction and growth of callus (Sun et al., 2006; Burbulis et al., 2007 ). However, in the present study 2,4-D, at each of the five concentrations, when used alone, was found as quite ineffective growth regulator for callus induction in T. peruviana. Other studies documenting the less efficient role of 2,4-D for callogenesis in different plant species are very few. Callus formation is in fact the reversion of mature differentiated cell to dedifferentiating state followed by high rate of cell division. Molecular mechanism of this reprogramming is understood only to a very limited extent. Lo Schiavo et al. (1989) found that more than normal auxin level inside the cells cause the DNA to become more methylated than usual and suggested that this might be necessary for the re-programming of differentiated cells. 2,4-D, when added exogenously, is reported to be associated with a change (increase/decrease) in natural auxin biosynthesis inside the plant cell, by many workers (Elliott, 1970;Michalczuk et al., 1992). The exact molecular mechanism of this change is not clear but the increase or decrease in natural auxin inside the cell seems to be strongly guided by genotype. The exogenous supply of 2,4-D to the cells of Thevetia peruviana may be causing a change, in the overall auxin level, which might not be suitable for reprogramming of cells, though the exact explanations needs a molecular level investigation.
Kinetin, at all the five concentrations, when used alone, exhibited significantly high % callus induction. The lowest frequency of callus induction, from nodal segments, observed among various kinetin supplemented media was much higher than the highest frequency observed among various 2,4-D supplemented media This was in contrast to the general observation that cytokinins supplemented to synthetic medium support the shoot induction and proliferation from the explants. Some other studies reporting efficient callus induction, from different explants, on medium supplemented with cytokinins only are also available for some other plant species as well (Prasad et al., 2004; Osman et al., 2010; Aasim et al., 2008). In tissue cultures, cytokinins appear to be necessary for plant cell division, might be required to regulate the synthesis of proteins involved in the formation and function of mitotic spindle apparatus (Jouanneau, 1970, 1975). Callus tissue in which cell division proceeds without the addition of cytokinin to the culture medium are thought to produce their own natural cytokinins in required amount (Zazimalova et al., 1996). Also many dicotyledonous calluses can be cultured for long periods on high cytokinins, without any auxin supplementation to the synthetic mediumand here the cytokinin appears to increase the natural auxin content of the tissue (Syono and Furuya, 1972).
The available molecular level studies indicate that the exogenously supplied growth regulator, auxins and cytokinins, each have a direct or indirect role in the endogenous synthesis of both- auxins as well as cytokinins (Syono and Furuya, 1972; Zazimalova et al., 1996). In fact, the auxin to cytokinin ratio represents an important signal in the formation of cell phenotype and also in the onset and maintenance of the process of cell division (Stickens et al., 1996). In the present study, as compared to use of kinetin and 2,4-D alone, combination of 2,4-D and kinetin gave much better results in terms of callus induction and proliferation. Callus induction with a frequency of hundred % was achieved on one combination type (MS medium supplemented with 0.93 μM kin alongwith 9.05 μM 2,4-D). Combinations of auxins and cytokinin (with high auxin to cytokinin ratio) have been reported to be favouring the callus induction in some other plants also like Acacia manginum (Xie and Hong, 2001), Gossypium hirsutum L. (Abdellatef and Khalafallah, 2008) Kaempferia galanga (Rahman et al., 2004).
Callus initiation starts much earlier in the explants than that become visible on the surface. On observing with naked eyes, an early response was observed on combination of 2,4-D and kinetin, as compared to that on the individual level of each. The earliest callus initiation from nodal explants (after a mean of 9.238 days) was observed on the combination of 0.93 μM kin with 9.05 μM 2,4-D, proliferating to a callus mass of 2.556 g after thirty days of culture. A similar level of fresh weight (2.075 g) was observed on 6.97 μM kinetin after thirty days, though callus initiation was observed after a mean of 21.06 days. It means that though cell division in the explant started much earlier on combination of auxin and cytokinin as compared to kinetin alone, division rate was much fast in kinetin rich medium as compared to that on auxin-cytokinin combination. A very poor proliferation resulting in callus with very less fresh weight was observed on medium supplemented with 2,4-D alone.
Sucrose is the main source of energy for in vitro plant tissue cultures as these have insufficient autotrophic ability. Sucrose not only acts as an external energy source but also contributes to the osmotic potential of the medium (Nowak et al., 2004). The osmotic potential would permit absorption of mineral nutrients, by the plant cells, present in medium essential to the cells growth and division. A significant effect of carbon source concentration, in culture media, on the osmotic potential and hence on the frequency of callus formation has been noticed in many plants like rice (Shahnewaz and Bari (2004) olive (Garcia et al., 2002) etc. In the present investigation, among the three levels of sucrose used, 3% concentration was found as the most favoring for callus initiation and proliferation. Mihaljevic et al. (2002) reported 20 % reduction in callus induction frequency from mature zygotic embryos of Taxus baccata L. Washingtonii when sucrose concentration was increased from 2 to 3%. On the other hand Shahnewaz and Bari (2004) observed high callus induction frequency in anther culture of rice on 4% sucrose, the frequency declining at low concentration of sucrose. The optimal sucrose concentration required for optimal callus induction and proliferation seems to be directed by explant nature as well as the genotype of the mother plant.
Repeated sub-culturing of callus is required for producing and maintaining it for a long time. During the present study, callus growth declined on media supplemented with kinetin alone. On the other hand auxin and cytokinin together (the combination of 0.93 μM kin with 9.05 μM 2,4-D) supported a healthy growth till the last (eighth) sub-culture. This requirement is probably governed by the genotype and the callus age as well.
CONCLUSION
We have developed a highly efficient protocol for sustainable callus production from stem segments of T. peruviana. The healthy and proliferative callus produced and maintained during the course of work can be subjected to many diverse applications like secondary metabolite isolation, large scale plant regeneration round the year, genetic transformation studies as well as the for studying the development pathways for cell differentiation of T. peruviana.
REFERENCES
- Aasim, M., K.M. Khawar and S. Ozcan, 2008. In vitro micropropagation from shoot meristems of turkish cowpea (Vigna unguiculata L.) cv. Akkiz. Bangladesh J. Bot., 37: 149-154.
CrossRefDirect Link - Abdellatef, E. and M.M. Khalafallah, 2008. Influence of growth regulators on callus induction from hypocotyls of medium staple cotton (Gossypium hirsutum L.). cultivar barac B-67. J. Soil Nat., 2: 17-22.
Direct Link - Abe, F., R.F. Chen and T. Yamauchi, 1996. Dinormonoterpenoids and their apiosylglucosides from Thevetia peruviana. Photochemistry, 43: 161-163.
PubMed - Afaq, F., S. Mohammad, H. Moammir and M. Hasan, 2004. Inhibition of 12-0-tetradecanoylphorbal-13-acetate induced tumor promotion marker in CD-1 mouse skin by Oleandrin. Toxicol. Appied Pharmacol., 195: 361-369.
CrossRef - Burbulis, N., A. Blinstrubiene, A. Sliesaravicius and R. Kupriene, 2007. Some factors affecting callus induction in ovary culture of flax (Linum usitatissimum L.). Biologia, 53: 21-23.
Direct Link - Che, P., D.J. Gingerich, S. Lall and S.H. Howell, 2002. Global and hormone induced gene expression changes during shoot development in Arabidopsis. Plant Cell, 14: 2771-2785.
Direct Link - Elliott, E.F., 1970. Axenic culture of meristem tips of Rosa multiflora. Planta, 95: 183-186.
CrossRef - Gata-Gonclaves, L., J.M.F. Nogueira, O. Matos and R.B. De-Sousa, 2003. Photoactive extracts from Thevetia peruviana with antifungal properties against Cladosporium cucumerinum. J. Photochem. Photobiol. B, 70: 51-54.
PubMed - Garcia, J.L., J. Troncoso, R. Sarmiento and A. Troncoso, 2002. Influence of carbon source and concentration on the in vitro development of olive zygotic embryos and explants raised from them. Plant Cell Tissue Organ Cult., 69: 95-100.
CrossRefDirect Link - Ikram-ul-Haq, 2005. Callus proliferation and somatic embryogenesis in cotton (Gossypium hirsutum L.). Afr. J. Biotechnol., 4: 206-209.
Direct Link - Ishii, Y., T. Jakamura, M. Goi and M. Tanaka, 1998. Callus induction and somatic embyogenesis of Phalaenopsis. Plant Cell Rep., 17: 446-450.
CrossRef - Jouanneau, J.P., 1975. Protein synthesis requirement for the cytokinin effect upon tobacco cell division. Exp. Cell Res., 91: 184-190.
CrossRef - Michalczuk, L., D.M. Ribnicky, T.J. Cooke and J.D. Cohen, 1992. Cohen regulation of indole-3-acetic acid biosynthetic pathways in carrot cell cultures. Plant Physiol., 100: 1346-1353.
PubMed - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Osman, M.G., E.A. Elhadi and M.M. Khalafalla, 2010. Callus formation and organogenesis of tomato (Lycopersicon esculentum Mill, C.V. Omdurman) induced by thidiazuron. Afr. J. Biotechnol., 9: 4407-4413.
Direct Link - Patil, H.S.R., H.K. Makari and H. Gurumurthy, 2007. In vitro antimicrobial activity of ethanol extract of Thevitia peruviana. Electr. J. Environ. Agric. Food Chem., 6: 2318-2322.
Direct Link - Rahman, M.M., M.N. Amin, T. Ahamed, M.R. Ali and A. Habib, 2004. Efficient plant regeneration through somatic embryogenesis from leaf base-derived callus of Kaempferia galanga L. Asian J. Plant Sci., 3: 675-678.
CrossRefDirect Link - Rao, S.R. and G.A. Ravishankar, 2002. Plant cell cultures: Chemical factories of secondary metabolites. Biotechnol. Adv., 20: 101-153.
CrossRefPubMedDirect Link - Shahnewaz, S. and M.A. Bari, 2004. Effect of concentration of sucrose on the frequency of callus induction and plant regeneration in anther culture of rice (Oryza sativa L.). Plant Tissue Cult., 14: 37-43.
Direct Link - Stickens, D., W. Tao and J.P. Verbelen, 1996. A single cell model system to study hormone signal transduction. Plant Growth Regul., 18: 149-154.
CrossRef - Sugiyama, M. and K. Imamura, 2006. Dose-, time- and tissue-dependent effects of 5-bromo-2`-deoxyuridine on the in vitro organogenesis of Arabidopsis thaliana. Plant Cell Tissue Organ Cult., 87: 17-25.
Direct Link - Sun, Y., X. Zhang, C. Huang, X. Guo and Y. Nie, 2006. Somatic embryogenesis and plant regeneration from different wild diploid cotton (Gossypium.) species. Plant Cell Rep., 25: 289-296.
CrossRef - Syono, K. and T. Furuya, 1972. Effects of cytokinins on the auxin content of tobacco calluses. Plant Cell Physiol., 13: 843-856.
Direct Link - Tewtrakul, S., N. Nakamura, M. Hattori, T. Fujiwara and T. Supavita, 2002. Flavanone and flavonol glycosides from the leaves of Thevetia peruviana and their HIV-1 reverse transcriptase and HIV-1 integrase inhibitory activities. Chem. Pharm. Bull., 50: 630-635.
PubMed - Xie, D. and Y. Hong, 2001. In vitro regeneration of Acacia mangium via organogenesis. Plant Cell Tissue Organ Cult., 66: 167-173.
CrossRef - Zazimalova, E., A. Brezinova, J. Holik and Z. Opantrny, 1996. Partial auxin deprivation affects endogenous cytokinins in an auxin dependent, cytokinins-independent tobacco cell strain. Plant Cell Rep., 16: 76-79.
CrossRef - Zibbu, G. and A. Batra, 2010. Effect of adeninesulphate on organogenesis via leaf culture in an ornamental plant: Thevetia peruviana (pers.) schum. Int. J. Pharma. Biosci., 1: 1-9.
Direct Link - Zouine, J. and I. El-Hadrami, 2004. Somatic embryogenesis in Phoenix dactylifera L: Effect of exogenous supply of sucrose on proteins, sugars, phenolics and peroxidases activities during the embryogenic cell suspension culture. Biotechnology, 3: 114-118.
CrossRefDirect Link