
Timir Baran Jha
Laboratory of Plant Biotechnology, Department of Botany, Presidency College, Kolkata-700073, India
LiveDNA: 91.19935
Arpita Dafadar
Laboratory of Plant Biotechnology, Department of Botany, Presidency College, Kolkata-700073, India
Rituparna Kundu Chaudhuri
Bethune College, Kolkata-700006, India
Asian Journal of Biotechnology
Year: 2011 | Volume: 3 | Issue: 2 | Page No.: 186-193







ABSTRACT
The primary objective of this present investigation was to induce embryogenesis from leaf explants of in vitro grown shoots of Swertia chirata, a multipotent indigenous medicinal herb. Somatic embryogenesis is an important system of micropropagation has not been reported earlier in any species of Swertia. Aseptically grown leaf segments of nearly 1 cm in length were placed in B5 medium with different concentration and combination of growth regulators. Eighty percent leaf explants within 6 weeks of culture produced nodular, creemish yellow calli on B5 medium supplemented with 4.5 μM 2, 4-D (2,4-dichlorophenoxy acetic acid) and 2.3 μM Kn (Kinetin). The highest frequency of embryogenesis (40%) with maximum number of globular somatic embryos (35.6±4.3) was recorded in the combination of 0.4 μM 2,4-D and 4.6 μM Kn per g of embryogenic calli. Addition of adenine sulfate (100 μM) helped in maturation of somatic embryos. Complete somatic embryo plants were obtained on transferring the cultures in basal B5 medium. Sixty percent plants survived in the field condition. The complete procedure required 20-22 weeks. Chromosomal stability was noted in somatic embryo plants.
PDF Abstract XML References Citation
How to cite this article
Timir Baran Jha, Arpita Dafadar and Rituparna Kundu Chaudhuri, 2011. Somatic Embryogenesis in Swertia chirata Buch. Ham. ex Wall. - A Multipotent Medicinal Herb. Asian Journal of Biotechnology, 3: 186-193.
DOI: 10.3923/ajbkr.2011.186.193
URL: https://scialert.net/abstract/?doi=ajbkr.2011.186.193
DOI: 10.3923/ajbkr.2011.186.193
URL: https://scialert.net/abstract/?doi=ajbkr.2011.186.193
INTRODUCTION
Medicinal plants are the traditional source of drugs. Swertia chirata, is an indigenous multipotent medicinal herb. The medicinally important bioactive compounds identified so far from this species, are ophelic acid, iridoid glycosides and xanthones. Its demand in the domestic and international market is increasing. On the other hand in nature, its seed viability is low and germination rate is very poor. Indiscriminate overharvesting from the wild is the present option of meeting the market demand. Indian National Medicinal Plant Board has listed this herb in their priority list and the present status of the plant is endangered. Keeping in mind its potentialities and problems we have systematically attempted to formulate alternative strategies of plant production and conservation for its sustainable use. We have reported mass propagation of genetically uniform plants through node culture and direct organogenesis from in vitro leaf explants (Chaudhuri et al., 2007, 2008). Indirect propagation using immature seeds has also been reported for the first time in chirata (Chaudhuri et al., 2009). Some other published reports on micropropagation of Swertia chirata are also available. Wawrosch et al. (1999), reported micropropagation of S. chirata using root segments of in vitro raised seedlings. A protocol for rapid shoot multiplication from nodal explants of S. chirata has been developed by Ahuja et al. (2003), which is under IPR protection. Joshi and Dhawan (2007) as well as and Balaraju et al. (2009) reported efficient shoot regeneration from axillary meristems of S. chirata. Miura et al. (1978) reported micropropagation in two other important species of Swertia namely, S. pseudo chinensis and S. japonica. Embryogenesis has been reported from the family Gentianaceae including Gentiana cruciata (L.), Gentiana pannonica (Scop.), Gentiana tibetica (King.) (Mikula and Rybczynski, 2001), Gentiana pneumonanthe (Bach and Pawloska, 2003), G. punctata L. (Mikula et al., 2004) and Gentiana kurroo Royle (Agnieszka and Rybczynski, 2008) using leaf, root, hypocotyl and cotyledon explants. On the other hand only one published report by Wang et al. (2009) claimed adventitious shoot regeneration from in vivo leaf explants of S. chirata through the formation of somatic embryos. It appears from the above noted reports that well documented published report of somatic embryogenesis in Swertia is lacking. Somatic embryos are considered important propagules of micropropgation and transgenic researches. It is considered to be an effective technique for plant regeneration because of its high-regeneration potential and low frequency of mutations. In this present communication we have described an efficient and well documented process of somatic embryogenesis from in vitro grown leaf tissues and their conversion to complete plants.
MATERIALS AND METHODS
This research project was conducted from January 2004 to Dec. 2008.
Plant materials and culture conditions: Plant materials were collected from West Sikkim (10,500 ft) in the month of December and the study was carried out in the Plant Biotechnology Lab., Department of Botany, Presidency College, Kolkata, India. Aseptic cultures with nodal explants were developed following our earlier protocol (Chaudhuri et al., 2007, 2008). Each leaf explants of nearly 1 cm length were dissected out from the leaves approximately 2.5-4 cm in length and taken from near to the apical meristem of in vitro grown shoots of Swertia chirata. A set of 10 explants per treatment was cultured and each experiment was repeated thrice. Data was recorded after 6 weeks of culture.
Somatic embryogenesis, maturation and their conversion to plantlets: B5 medium (Gamborg et al., 1968) was used to induce embryogenic calli with various concentrations and combinations of 2, 4-D (4.5-9 μM), NAA (Napthalene acetic acid) (4.5-9 μM) and Kn (2.32-4.65 μM). The pH of the culture media was adjusted to 5.5 before autoclaving. All the aseptic cultures were incubated at 22±2°C under a photoperiod of 16 h light/8 h dark cycles. Subcultures were performed at an interval of 3 to 4 weeks.
The 6-8 week-old embryogenic calli were subcultured on B5 medium with same additives mentioned above for another 4-6 weeks. The embryogenic calli induced globular somatic embryos which were transferred to the maturation medium i.e., B5 medium with reduced amount of 0.4 μM 2, 4-D, 4.6 μM Kn and 100 μM adenine sulphate. After 6 weeks of culture, the mature somatic embryos were germinated in B5 basal medium. The embryo-derived plantlets were hardened following earlier protocol (Chaudhuri et al., 2007, 2008) and transferred to the polyhouse of our experimental garden at the Presidency College Kolkata.
Chromosomal analysis: Healthy root-tips from somatic embryo plants were pre-treated with 0.002 M Hydroxyquinoline for 3 h at room temperature followed by overnight fixation in Carnoy’s fluid. Materials were stained with Orcein, HCl mixture (9:1) for 4 h and squashed in 45% acetic acid. Chromosome numbers per cell were manually counted and photographs were taken under Zeiss photomicroscope.
Statistical analysis: The experiments were set up in a randomized design. Data were analyzed by analysis of variance (ANOVA) to detect significances between means. Means differing significantly were compared using Duncan’s Multiple Range Test (DMRT) at a 5% probability level. Variability of data has also been expressed as the Mean±Standard Error (SE).
RESULTS
Induction of embryogenic calli and development of somatic embryos: Leaf explants started swelling within 3 weeks. Initiation of callus was noted on cut end and midrib region of lower surfaces of the leaf segments after 4 weeks of culture. Out of many hormonal combinations, 80% leaf explants within 6 weeks of culture produced calli on B5 medium supplemented with 4.5 μM 2, 4-D, 2.3 μM Kn (Table 1). The calli was nodular, creamish yellow in nature (Fig. 1a). Embryogenic calli were differentiated on the edges and highly organized, round, clusters of globular somatic embryos were visible on lowering the concentrations of 2,4-D after 4-6 weeks of culture (Table 2). It was also found to be embedded in the embryogenic callus tissue, while other areas of the calli remained white and translucent. The highest frequency of embryogenesis (40%) was recorded in the combination of 0.4 μM 2,4-D and 4.6 μM Kn (Table 2). In this medium maximum number (35.6±4.3) of globular and heart shaped somatic embryos were also developed (Fig. 1b). Globular somatic embryos on subculture to the same embryo tissue proliferation medium were found to proliferate gradually with distinct bipolarity (Fig. 1c). The process of embryogenesis in Swertia chirata is asynchronus (Fig. 1d) and we obtained various stages of somatic embryos such as oblong, heart and cotyledonary stage embryos with distinct bipolarity in one culture.
Maturation of somatic embryos: Present study revealed that a minimum time of 4-6 weeks were required for conversion of globular somatic embryos to mature embryos. Here addition of 100 μM adenine sulphate along with 0.4 μM 2,4-D and 4.6 μM Kn has led to significantly highest number (25.6±4.2) of mature somatic embryos (Table 2).
| Table 1: | The influences of PGRs on embryogenic callus induction frequency from leaf explants of Swertia chirata on B5 medium |
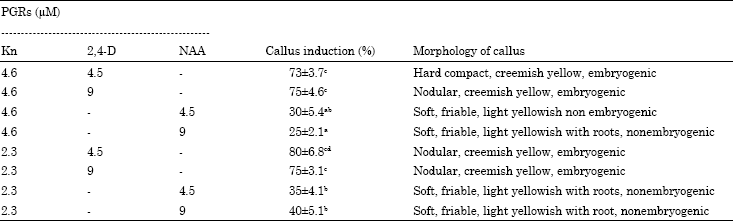 | |
| Values represent the Mean±SE explants. Means having different letters in superscript are significantly different from each other (p<0.05) according to Duncan’s Multiple Range Test | |
 | |
| Fig. 1: | (a) Yellow coloured, nodular, embryogenic calli developed after 6 weeks of culture, (b) Calli showing globular and heart shaped embryos after another 4-6 weeks, (c) Globular shaped embryos showing distinct bipolarity and development of shoot primordial, (d) Various stages of somatic embryo development showing globular, heart and mature cotyledonary stages, (e-f) Germination and proliferation of embryos after 14-16 weeks of culture, (g) Complete rooted plant derived from somatic embryos, (h-i) Acclimatized and hardened S. chirata plants growing under field condition and (j) Chromosomal analysis from root tip of a regenerant showing 2n=26 number of stable chromosomes in metaphase |
| Table 2: | Somatic embryogenesis from embryogenic calli, germination and plantlet conversion in Swertia chirata on B5 medium |
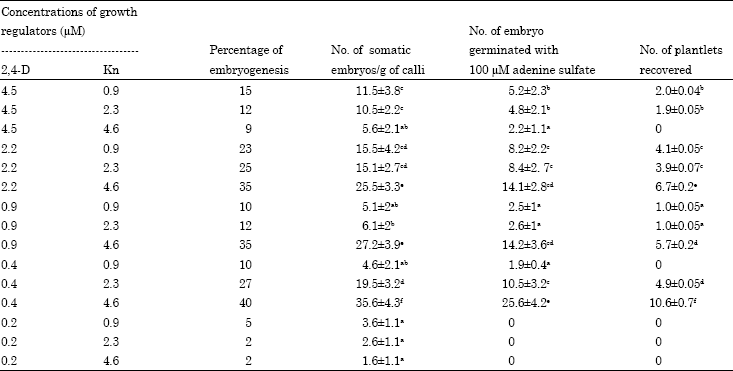 | |
| Values represent the Mean±SE. Means having different letters in superscript are significantly different from each other (p<0.05) according to Duncan’s Multiple Range Test Wang et al. (2009) | |
Germination and hardening of complete plantlets: Germination of somatic embryos was obtained in maturation medium earlier mentioned within the next 3-4 weeks time (Table 2). Further proliferation of shoots and roots were obtained on transferring the cultures in basal B5 medium. Figure 1e shows germination and Fig. 1f shows proliferation of the embryos. Nearly 10.6±0.7 number of complete plantlets could be recovered per culture. The somatic embryo plants with well-developed shoot and root system attaining an average length of 4.2 cm (Fig. 1g) were transferred in pots. Sixty percent plants survived in the field condition (Fig. 1h, i) at Presidency College campus, Kolkata, India. The complete procedure required 20-22 weeks.
Chromosomal analysis: Chromosomal analysis revealed presence of uniform diploid chromosome number of 2n=26 small chromosomes (Fig. 1j). No cytological anomaly was observed in our studies, indicating chromosomal stability in the somatic embryo plants.
DISCUSSION
In our earlier studies with S. chirata we obtained direct organogenesis with NAA, BAP (Benzyl amino purine) and Kn from in vitro leaf explants and organogenic calli from immature seed culture in presence of 2,4-D (Chaudhuri et al., 2008, 2009). However the present report confirmed for the first time that the leaves of in vitro grown shoots could be ideal explants for induction of embryogenic calli. The role of auxins and cytokinins in the different stages of somatic embryogenesis is well established (Fujimara and Komamine, 1980; Jimenez, 2001). The combined effect of auxin and cytokinins observed in this present system is in accordance with culture response of somatic embryogenesis in hybrid tea roses (Noriega and Sondahl, 1991), in Cichorium intybus L. (Abdin and Ilah, 2007) and in Oryza nivara (Jaseela et al., 2009). In an earlier report, Wang et al. (2009), has highlighted the effects of phytohormones and media in regeneration of shoots from in vivo leaf explant of S. chirata. He obtained maximum 7 number of shoots in 190-2 media supplemented with 13.32 μM 6-benzylaminopurine and 0.54 μM α-naphthaleneacetic acid. Histological studies revealed that shoot regeneration took place through somatic embryogenesis. However, the present report for the first time has properly documented the stages of somatic embryogenesis with recovery of higher number (10.6) of plantlets from aseptic culture.
In earlier studies with different Gentiana species it was evident that presence of both 2,4-D and Kn was required for induction of embryogenic calli. Also reduction in 2,4-D concentration was necessary for conversion embryogenic calli to somatic embryo (Mikula and Rybczynski, 2001; Agnieszka and Rybczynski, 2008). Present study also showed that 2,4-D along with Kn played an important role in inducing embryogenic calli not only in 80% explants but 2,4-D in reduced concentration (0.4 μM) along with 4.6 μM Kn was required for further development of somatic embryos in maximum number (35.6±4.3). In this context it can be mentioned that such conversion of embryogenic calli to globular somatic embryo may be attributed to the continued presence of lowered 2,4-D, that promotes the completion of globular stage during embryogenesis (LoSchiavo et al., 1989).
Formation of distinct bipolar somatic embryos with root and shoot poles may be attributed to the accumulation of endogenous level of auxin (2,4-D) and cytokinin (Kn). Auxin contributes towards polar development of embryos and cytokinin stimulates shoot and tap root formation (Moncousin et al., 1989; Chang, 1991). However some low frequency oblong embryos that occurred as an intermediate stage between globular and heart shaped stage was earlier described by Schiavone and Cooke (1985).
Maturation of somatic embryo is an important step towards formation of a complete plant. Reduced amount of 2, 4-D and addition of adenine sulphate has helped to enhance the maturation process of somatic embryos in many species including Gentiana pneumonanthe, an allied family of Swertia (Das et al., 1993; Bach and Pawlowska, 2003; Martin, 2003; Jha et al., 2007; Agnieszka and Rybczynsky, 2008).
Chromosomal stability has also been reported from micropropagated plants, leaf regenerants and even short term callus regenerants in Swertia chirata (Chaudhuri et al., 2007, 2008, 2009). All our results confirm stability of Swertia genome at the chromosomal level.
Considering the multipotent value, endangered status and being a priority listed plant, somatic embryogenesis in S. chirata will help not only to formulate a new system of propagation and conservation but also as a potential alternative system in the field of medicinal plant biotechnology.
ACKNOWLEDGMENT
We acknowledge the financial support of the Council of Scientific and Industrial Research, New Delhi, India. (Sanction No. 38/(1068)/EMR-II).Authors contribution are equal.
REFERENCES
- Abdin, M.Z. and E. Ilah, 2007. Plant regeneration through somatic embryogenesis from stem and petiole explants of Indian chicory (Cichorium intybus L.). Indian J. Biotechnol., 6: 250-255.
Direct Link - Bach, A. and B. Pawlowska, 2003. Somatic embryogenesis in Gentiana pneumonanthe L. Acta Biologica Cracoviensia Ser. Botanica, 45.2: 79-86.
Direct Link - Balaraju, K., P. Agastian and S. Ignacimuthu, 2009. Micropropagation of Swertia chirata Buch.-Hams. ex Wall.: A critically endangered medicinal herb. Acta Physiol. Plant., 31: 487-494.
CrossRefDirect Link - Chaudhuri, R.K., A. Pal and T.B. Jha, 2007. Production of genetically uniform plants from nodal explants of Swertia chirata Buch. Ham. ex Wall.-an endangered medicinal herb. In Vitro Cell. Dev. Biol. Plant, 43: 467-472.
CrossRefDirect Link - Chaudhuri, R.K., A. Pal and T.B. Jha, 2008. Conservation of Swertia chirata through direct shoot multiplication from leaf explants. Plant Biotechnol. Rep., 2: 213-218.
CrossRefDirect Link - Chaudhuri, R.K., A. Pal and T.B. Jha, 2009. Regeneration and characterization of Swertia chirata Buch.-Ham. ex Wall. plants from immature seed cultures. Sci. Hortic., 120: 107-114.
CrossRef - Das, P., G.R. Rout, and A.B. Das, 1993. Somatic embryogenesis in callus cultures of Mussaenda erythrophylla cvs. Queen sirikit and rosea. Plant Cell Tissue Organ Cult., 35: 199-201.
CrossRefDirect Link - Gamborg, O.L., R.A. Miller and K. Ojima, 1968. Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell Res., 50: 151-158.
CrossRefPubMedDirect Link - Jha, T.B., P. Mukherjee and M.M. Datta, 2007. Somatic embryogenesis in Jatropha curcas Linn., an important biofuel plant. Plant Biotechnol. Rep., 1: 135-140.
CrossRefDirect Link - Jimenez, V.M., 2001. Regulation of in vitro somatic embryogenesis with emphasis on the role of endogenous hormones. Revista Brasileira Fisiologia Vegetal, 13: 196-223.
CrossRef - Joshi, P. and V. Dhawan, 2007. Axillary multiplication of Swertia chirayita (Roxb. Ex Fleming) H. Karst., a critically endangered medicinal herb of temperate Himalayas. In Vitro Cell. Dev. Biol. Plant, 43: 631-638.
CrossRefDirect Link - LoSchiavo, F., L. Pitto, G. Giuliano, G. Torti and V. Nuti-Ronchi et al., 1989. DNA methylation of embryogenic carrot cell cultures and its variation as caused by mutation, differentiation, hormones and hypomethylating drugs. Theor. Applied Genet., 77: 325-331.
CrossRefDirect Link - Mikula, A. and J.J. Rybczynski, 2001. Somatic embryogenesis of Gentiana: Genus I. The effect of preculture treatment and primary explant origin on somatic embryogenesis of Gentiana cruciata (L.), Gentiana pannonica (Scop.), Gentiana tibetica (King). Acta Physiol. Plantarum, 23: 15-25.
Direct Link - Mikula, A., T. Tykarska, M. Zielin, M. Kuras and J.J. Rybczynski, 2004. Ultrastructural changes in zygotic embryos of Gentiana punctata L. during callus formation and somatic embryogenesis. Acta Biol. Cracoviensia Ser. Bot., 46: 109-120.
Direct Link - Martin, K.P., 2003. Plant regeneration through direct somatic embryogenesis on seed coat explants of cashew (Anacardium occidentale L.). Sci. Hortic., 98: 299-304.
CrossRefDirect Link - Schiavone, F.M. and T.J. Cooke, 1985. A geometric analysis of somatic embryo formation in carrot cell cultures. Can. J. Bot., 63: 1573-1578.
CrossRefDirect Link - Wang, L., L. An, Y. Hu, L. Wei and Y. Li, 2009. Influence of phytohormones and medium on the shoot regeneration from leaf of Swertia chirata Buch.-Ham. Ex wall. in vitro. Afr. J. Biotechnol., 8: 2513-2517.
Direct Link - Wawrosch, C., N. Maskay and B. Kopp, 1999. Micropropagation of the threatened Nepalese medicinal plant Swertia chirata Buch.-Ham. ex Wall. Plant Cell Rep., 18: 997-1001.
CrossRefDirect Link - Moncousin, C., J.M. Favre and T. Gaspar, 1989. Early changes in auxin and ethylene production in vine cuttings before adventitious rooting. Plant Cell Tissue Org. Cult., 19: 235-242.
CrossRefDirect Link