
Mukunda Chethan Kumar
Department of Biochemistry, JSS College of Arts, Commerce and Science (Autonomous), Ooty Road, Mysore-570025, Karnataka, India
Kondagula Shivappa Swetha
Department of Biochemistry, JSS College of Arts, Commerce and Science (Autonomous), Ooty Road, Mysore-570025, Karnataka, India
Asian Journal of Biochemistry
Year: 2015 | Volume: 10 | Issue: 2 | Page No.: 78-85







ABSTRACT
This study was conducted to elucidate the effect of bioconjugation of oligodeoxynucleotides with Lutein on antifibrotic activity using cultured adult human retinal pigment epithelial (ARPE-19) cells as model system. Fibrosis was induced in ARPE-19 cells by N-Nitrosodimethylamine (DMN) at a dose of 10 μL (diluted 1:100 with 0.15 M sterile NaCl). Oligodeoxynucleotides specific for Hepatocyte Growth Factor (HGF) and p38 Mitogen Activated Protein Kinase (MAPK) genes modified with a sulfhydryl group at the 3' end (500 nmol) was conjugated with 10 mg of Lutein. Expression of HGF, p38 MAPK and production of extracellular matrix proteins (ECM) such as collagen type I and fibronectin were assayed. The intracellular expression of Hepatocyte Growth Factor (HGF) and its receptor gene, p38 MAPK showed suppression in ARPE-19 cells treated with DMN+Lutein 15 μg/20 μL conjugated with oligodeoxynucleotides for HGF and p38 MAPK, respectively. A 50% of ARPE-19 cells showed an increase in type I collagen (582±92 pg) in DMN treated cells versus 220±42, p<0.001 (untreated) and it was 389±61 pg in cells treated with DMN+Lutein 15 μg/20 μL oligodeoxynucleotides conjugated. The fibronectin level was 1,422±60 pg in DMN treated cells versus 360±24, p<0.001 (untreated) and it was 760±59 pg in cells treated with DMN+Lutein 15 μg/20 μL oligodeoxynucleotides conjugated. These results demonstrate that bioconjugation of ODNs with Lutein would significantly suppress HGF/cMet, p38MAPK expression, collagen type I, fibronectin and quench the associated reactive oxygen species contributing for the progression of fibrosis in ARPE-19 cells.
PDF Abstract XML References Citation
Received: June 08, 2014;
Accepted: March 06, 2015;
Published: June 04, 2015
How to cite this article
Mukunda Chethan Kumar and Kondagula Shivappa Swetha, 2015. Effect of Bioconjugation of Oligodeoxynucleotides with Lutein on N-nitrosodimethylamine Induced Fibrosis in Cultured Human Retinal Pigment Epithelial Cells. Asian Journal of Biochemistry, 10: 78-85.
DOI: 10.3923/ajb.2015.78.85
URL: https://scialert.net/abstract/?doi=ajb.2015.78.85
DOI: 10.3923/ajb.2015.78.85
URL: https://scialert.net/abstract/?doi=ajb.2015.78.85
INTRODUCTION
Fibrosis is defined as a wound-healing response that has gone out of control (Pellicoro et al., 2014; Cheng et al., 2009). In the eye, of which the retina is a part, such wound-healing responses and associated fibrosis are mediated by Retinal Pigment Epithelium (RPE) cells and retinal glial cells which perform functions similar to those performed by fibroblasts in the rest of the body. Under the retina, fibrosis can occur subsequent to subretinal hemorrhage associated with neovascular age-related macular degeneration (Danis et al., 2014; Qazi et al., 2009). Once new blood vessels begin to grow in the eye, they are prone to rupture, leading to further activation of wound-healing responses and ultimately the development of severe macular fibrosis.
Pathogenic fibrosis typically results from chronic inflammatory reactions which stimulate the deposition of connective tissue elements that progressively remodel and destroy normal tissue architecture (Katta et al., 2009; Wang et al., 2005). TGF-β (Transforming growth factor-β) signaling favors the transition of RPE cells and retinal glial cells to fibroblast-like cells, stimulates the synthesis of ECM proteins and inhibits its degradation (Ehrlich et al., 2008). Transition of RPE cells into fibrotic scar is also modulated by soluble growth factors and cytokines such as tumor necrosis factor-α (TNF-α), interleukin-1 (IL-1) and Platelet-Derived Growth Factor (PDGF) secreted by inflammatory cells which are drawn to the lesion site (Hirasawa et al., 2011). The basic pathomolecular mechanism by which the sedentary RPE cells become activated is still poorly understood. Among the involved factors, Hepatocyte Growth Factor (HGF) and its receptor c-Met is associated with increasing the mobility of RPE cells. Oxidative stress is shown to be yet another risk factor in macular fibrosis and is involved in inducing Vascular Endothelial Growth Factor (VEGF) and Connective Tissue Growth Factor (CTGF) expression in RPE cells and retinal glial cells (Jun et al., 2007).
Strategies aimed at disrupting signaling pathways involving HGF/c-Met expression and downstream mediators along with potent antioxidant and anti-inflammatory molecule is of vital importance in preventing macular fibrosis in Age-related Macular Degeneration (ARMD). Lutein is a carotenoid which, along with its stereo isomer Zeaxanthin is concentrated in the macula lutea. Current approaches aimed at treating macular fibrosis in ARMD and other diseases of the eye are primarily directed at inhibiting promoters of angiogenesis, such as VEGF, cytokines, specific Matrix Metalloproteinases (MMPs) and adhesion molecules (integrins) (Jonas et al., 2004; Saika et al., 2005). Unfortunately, pharmacological interventions are typically initiated at an advanced stage of disease and do not target the underlying pathophysiology, therefore, they often fail to substantially impact the progression of disease. A more successful therapeutic approach should involve restoring homeostasis to the areas that underlie the fibrotic response. In addition to the aberrant expression of soluble growth factors and cytokines, Reactive Oxygen Species (ROS) generation is shown to induce cytokine production, inflammatory cell recruitment and tissue injury in fibrotic diseases. Emergence of various gene silencing technologies has boosted the possible use of oligodeoxynucleotides (ODNs) to treat fibrovascular diseases (Xu et al., 2009). The potent gene silencing ability of ODNs and bioconjugating with Lutein and directing towards achieving a site-specific delivery is required to avoid toxic side effects in the body and maximize ODNs efficiency in vivo. This kind of novel approach will prevent macular injury, reduce inflammation, inhibit fibroblast-like cells activation, degrade scar matrix and inhibit aberrant collagen synthesis. Therefore, the present study is focused on the use of bioconjugated ODNs with Lutein for gene silencing and further quenching of ROS in modulating N-Nitrosodimethylamine (DMN) induced fibrosis using cultured adult human retinal pigment epithelial (ARPE-19) cells, thus restoring the normal cellular structure and function.
MATERIALS AND METHODS
Lutein and other chemicals: Lutein was obtained from Sigma-Aldrich (St. Louis, MO). All the other chemicals and solvents used were of analytical grade from Merck Biosciences India Pvt. Ltd. The Kits for gene expression were procured from Invitrogen Biosciences Pvt. Ltd. Bangalore, Karnataka, India. ARPE-19 cells were obtained from the American Type Culture Collection (ATCC) (Manassas, VA). Dulbecco’s Modified Eagle’s Medium (DMEM): Nutrient Mixture F12, fetal bovine serum (FBS), bovine serum albumin (BSA), trypsin-EDTA, 4-(2-hydroxyethyl)-1- piperazineethanesulfonic acid (HEPES), sodium bicarbonate, gentamycin, Phosphate-Buffered Saline (PBS), Triton X-100, penicillin and streptomycin was procured from Sigma-Aldrich (St. Louis, MO).
Cell culture: ARPE-19 cells were cultured in 95% air and 5% CO2 at 37°C in DMEM/F12 medium containing 10% FBS, 50 units mL-1 penicillin and 50 μg mL-1 streptomycin. The cells were seeded (5000 cells/cm2) on 60 mm multidishes. The culture medium was changed on the second day and confluent cultures were obtained on the third day. Cells were then immediately processed for the analyses (Maminishkis et al., 2006).
Bioconjugation of oligodeoxynucleotides (ODNs): ODNs modified with a sulfhydryl group at the 3’ end (500 nmol) was treated with 0.2 M dithiothreitol in 1.5 mL of 0.1 M, pH 9.0, glycine buffer containing 0.1 M NaCl for 3 h at room temperature to generate a 3’-thiol functional group with modifications to the earlier reported method (Gamper et al., 1993). ODNs were precipitated by adding 3 volumes of ethanol after the addition of sodium acetate to 0.3 M. The mixture was kept at -30°C overnight and centrifuged at 12,000 g for 30 min at 4°C. A 5 mg of ODNs with 3’-thiol functional group was dissolved in 600 μL of boric acid/borax buffer (pH 8.0, 60 mM). A 10 mg of Lutein was added, followed by 1.5 mL of dimethylformamide. The mixture was incubated with stirring under nitrogen protection at 40°C for 24 h.
HGF/c-Met expression assay: Fibrosis was induced in ARPE-19 cells by dimethylnitrosamine (DMN) at a dose of 10 μL (diluted 1:100 with 0.15 M sterile NaCl). The total RNA was isolated from the ARPE-19 cells treated with or without DMN and test samples (Lutein 15 μg/20 μL conjugated with ODNs for HGF) and was used for the first strand of cDNA synthesis. The resulting RT products were amplified with the following sets of primers: HGF forward primer 5’-ATC-CTC ATG GAC CCT GGT G-3’ and HGF reverse primer 5’-GGC CTG GCA AGC TTC ATT A-3’, c-Met/HGF-R forward primer 5’-AGA TCA TCC ATT GCA TTC GA-3’ and c-Met/HGF-R reverse primer 5’-TGA CGA TCT TGT TGA AGA AG-3’. Southern blot analysis of the PCR products was conducted and detection of the probes was accomplished using antidigoxigenin-alkaline phosphatase Fab fragments and the chemiluminescence alkaline phosphatase substrate (Jun et al., 2007).
p38 MAP kinase assay: p38 MAPK activity in the ARPE-19 cells was quantified by using ELISA (Saika et al., 2005). Cells were seeded in 96-well culture plates. While still subconfluent, the cells were treated with or without DMN and test samples (Lutein 15 μg/20 μL conjugated with ODNs for p38 MAPK) for specific time intervals. Specimens were then processed for detection of p38 MAPK activity using antibodies. After reaction with the primary and then secondary antibodies the color reaction was performed. Absorbance at 450 nm was determined by a UV-Vis microplate spectrometer reader.
Collagen type I and fibronectin assay: Productions of collagen type I and fibronectin, the major ECM components were assayed (Dooley et al., 2010). ARPE-19 cells grown to confluency in a six-well culture plate in DMEM/F-12 supplemented with 15% fetal calf serum was further incubated with or without DMN and test samples (Lutein 15 μg/20 μL conjugated with ODNs for collagen type I and fibronectin. At the end of the culture interval, the medium was harvested and cells were sonicated in 500 mL of PBS. Culture medium and cell lysate were processed for collagen type I C-terminal peptide and fibronectin analysis using ELISA. Color reaction was measured at 450 nm.
Data analysis: Analysis of Variance (ANOVA) test, followed by individual comparison by Student’s‘t’-test for the determination of level of significance among the Mean±SEM in various groups were performed.
RESULTS
In the present study, ODNs conjugated with Lutein as mentioned above was found to be stable and possess signature wavelength at 471 nm which is specific for Lutein still after bioconjugation. Reverse transcriptase-PCR method was used to detect HGF and p38 MAPK mRNA expressions. In DMN treated cells HGF/cMet expression was significantly increased in comparison to their respective control (untreated) cells. DMN+Lutein (15 μg/20 μL) conjugated with ODNs for HGF showed significant suppression in HGF/cMet expression (Fig. 1).
The results show that p38 MAPK expression is significantly increased in DMN treated ARPE-19 cells in comparison to their respective control (untreated) cells. DMN+Lutein (15 μg/20 μL) conjugated with ODNs for p38 MAPK showed significant suppression (Fig. 2), suggesting that suppression of expression of p38 MAPK would inhibit the onset and progression of fibrosis induced by DMN in ARPE-19 cells.
To elucidate the effect of Lutein conjugated ODNs interactions with regard to ECM proteins production, ARPE-19 cells were cultured with or without DMN and test samples as detailed above. Type I collagen and fibronectin production were quantified by enzyme-linked immunosorbent assay.
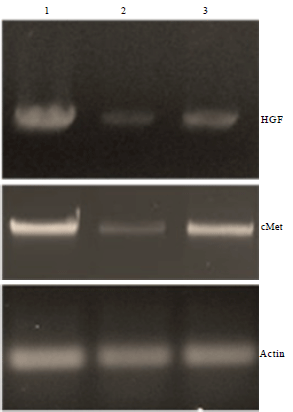 | |
| Fig. 1: | Analysis of HGF/cMet expression in ARPE-19 cells, Lane 1: Treated cells (with DMN), Lane 2: Treated Cells (with DMN + Lutein 15 μg/20 μL conjugated with ODNs for HGF/cMet) and Lane 3: Untreated cells |
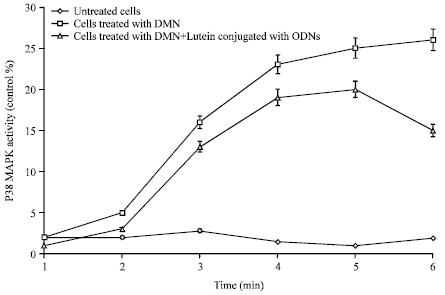 | |
| Fig. 2: | Quantification of activation of p38 MAP Kinase, p38 MAPK activity in the cells was quantified by using ELISA. Absorbance at 450 nm was determined by a UV-Vis microplate spectrometer reader |
Fifty percent of cells showed an increase in type I collagen (582±92 pg) in DMN treated cells versus 220±42 pg, p<0.001 (untreated) and it was 389±61 pg in DMN + Lutein 15 μg/20 μL conjugated with ODNs for type I collagen. The fibronectin level was 1,422±60 pg in DMN treated cells versus 360±24 pg, p<0.001 (untreated) and it was 760±59 pg in DMN + Lutein 15 μg/20 μL conjugated with ODNs for fibronectin.
DISCUSSION
Homeostasis of the eye, as in tissues elsewhere in the body, depends on the presence of normal vasculature, Extra Cellular Matrix (ECM) and various cell types (Mochizuki et al., 2013). If homeostasis is disturbed by infection, inflammation and or ROS, visual function becomes impaired. The end result of these conditions is often fibrosis. Fibrosis is a wound-healing response that has gone out of control resulting in substantial remodeling of the ECM. In the retina, such wound-healing responses and associated fibrosis are mediated by RPE cells and retinal glial cells. The changes that frequently occur in RPE cells in the vitreous cavity and subretinal space include proliferation and production of ECM components on the retina. TGF-β, a potent fibrotic factor is known to induce the transformation of RPE to fibroblast-like cells, production of ECM such as collagen type I and fibronectin and cell migration. An excessive accumulation of ECM components contribute to the formation of a permanent fibrotic scar (Wynn, 2011). Therefore a potent antifibrotic therapy is urgently needed to eliminate progression to fibrosis.
ODNs are short, single-stranded nucleic acids which disrupt expression of target protein by binding to complementary mRNA or forming triplex with genomic DNA. The pharmacological activity and pharmacokinetics of ODNs can be significantly improved by conjugation with Lutein, the dietary xanthophyll carotenoid. Lutein is a potent antioxidant and anti-inflammatory molecule (Ciccone et al., 2013).
HGF, a multifunctional protein act as a potent mitogen has now emerged as a potent antifibrotic cytokine that prevents tissue fibrosis. It is also thought to be a growth factor for mature hepatocytes. Although, HGF is present at low concentrations in the serum of apparently normal individuals, serum concentrations of HGF rise in response to many pathological consequences such as liver injury. In contrast, exogenous administration of HGF in animal models prevented onset of hepatic fibrosis/cirrhosis and progression of hepatic disease (Liu, 2004; Matsumoto and Nakamura, 2001; Matsuda et al., 1997; Ueki et al., 1999).
The importance of pathological changes leading to progressive fibrosis in the macular region is not well recognized. With increasing awareness of the importance of the pathological changes, interest has focused on the role of transformation of RPE cells to fibroblasts like cells in the initiation and progression of a fibrotic response. In macular fibrosis, the phenotypic transformation RPE cells from the quiescent to the activated state are accompanied by increased expression of HGF/cMet, p38MAPK and accumulation of extracellular matrix proteins, such as collagen type-I and fibronectin (Naylor et al., 2013).
In this study, it is demonstrated that the expression of HGF/cMet was spontaneously induced in cultured ARPE-19 cells. The overexpression of cMet may be associated with the increased expression of matrix metalloproteases which are mediated by HGF expression. HGF, although has an antifibrotic effect, HGF levels may be insufficient to prevent over production of matrix proteins possibly due to ROS generation (Ghatak et al., 2014).
The RPE cells function in an environment that is rich in endogenous Reactive Oxygen Species (ROS). The activity of RPE cells, the high local oxygen concentration and the exposure to light contribute to the production of ROS (Xie et al., 2011). Although multiple physiologic mechanisms protect the RPE from the toxic effects of light and oxidative damage, mounting evidence suggests that chronic exposure to oxidative stress over the long term may damage the RPE and predispose it to the development of macular fibrosis (Kim et al., 2010). Previous reports on the effect of Lutein in preventing oxidative DNA damage in relation to Age-Related Macular Degeneration (ARMD) is shown to play a significant role in modulating pathogenesis of ARMD.
Conjugation of Lutein with oligodeoxynucleotides is basically a carotenoid compound with strong antioxidative activity. The estimated intracellular expression of Hepatocyte Growth Factor (HGF), a known cell motility factor, in ARPE-19 cells showed the suppression of HGF and its receptor gene expression. The results suggest an involvement of the antioxidative property of Lutein and specific inhibition of HGF gene expression by oligodeoxynucleotides.
Protein kinases regulate several important functions within cells including metabolism, cell cycle progression, angiogenesis, cell adhesion, etc. Specifically, Mitogen-Activated Protein Kinases (MAPK) play a central role in the cellular response to environmental stress, growth factors and cytokines. The serine/threonine kinase, p38 is a member of the MAPK family and has been shown to be a critical enzyme in cell proliferation and the secretion of cytokines. Intense efforts are underway to find inhibitors of this enzyme for the treatment of many diseases including fibrosis. p38 MAPK, one of the mitogen activated protein kinase is responsive to oxidative stress (Gaitanaki et al., 2003). DMN treatment can induce ROS generation via oxidative stress reactions in ARPE-19 cells which in turn, activate the p38 MAPK signal pathway and both ROS and p38 MAPK molecules are possibly involved in fibrosis onset.
In fibroproliferative disorders, a tissue injury is associated with activation of collagen producing cells, followed by the excessive synthesis and deposition of type I collagen (Fritz et al., 2011). In activated cells, the increased expression of collagen is the result of the increased rate of transcription of collagen I and fibronectin genes and the increased half-life of collagen I and fibronectin mRNAs. Targeting molecules involved in transcriptional and post transcriptional regulation of expression of type I collagen could lead to potential antifibrotic therapies. From the study it was found that Lutein conjugated with ODNs specific for type I collagen and fibronectin reduced the signaling of these molecules in a time and concentration-dependent manner. Carotenoids such as Lutein is shown to reduce platelet derived growth factor and hydrogen peroxide signaling and compromises migration in cultured vascular smooth muscle cells. Lutein conjugated with ODNs for type I collagen and fibronectin specifically modulate the over production of these ECM proteins. Thus bioconjugation of ODNs with Lutein would significantly suppress HGF/cMet, p38 MAPK expression, collagen type I, fibronectin and quench the associated reactive oxygen species contributing for the progression of fibrosis.
ACKNOWLEDGMENT
MCK and SKS gratefully acknowledge the financial support from Indian Council of Medical Research (ICMR), No. 52/2/2011-BMS, Government of India (GoI), India for carrying out this study.
REFERENCES
- Pellicoro, A., P. Ramachandran, J.P. Iredale and J.A. Fallowfield, 2014. Liver fibrosis and repair: Immune regulation of wound healing in a solid organ. Nat. Rev. Immunol., 14: 181-194.
Direct Link - Maminishkis, A., S. Chen, S. Jalickee, T. Banzon and G. Shi et al., 2006. Confluent monolayers of cultured human fetal retinal pigment epithelium exhibit morphology and physiology of native tissue. Invest. Ophthalmol. Vis. Sci., 47: 3612-3624.
PubMed - Dooley, A., X. Shi-Wen, N. Aden, T. Tranah and N. Desai et al., 2010. Modulation of collagen type I, fibronectin and dermal fibroblast function and activity, in systemic sclerosis by the antioxidant epigallocatechin-3-gallate. Rheumatology, 49: 2024-2036.
PubMed - Gaitanaki, C., S. Konstantina, S. Chrysa and I. Beis, 2003. Oxidative stress stimulates multiple MAPK signalling pathways and phosphorylation of the small HSP27 in the perfused amphibian heart. J. Exp. Biol., 206: 2759-2769.
PubMed - Jun, E.J., H.S. Kim and Y.H. Kim, 2007. Role of HGF/c-Met in Serum-Starved ARPE-19 cells. Korean J. Ophthalmol., 21: 244-250.
CrossRefPubMedDirect Link - Fritz, D., L. Cai, L. Stefanovic and B. Stefanovic, 2011. Progress towards discovery of antifibrotic drugs targeting synthesis of type I collagen. Curr. Med. Chem., 18: 3410-3416.
Direct Link - Gamper, H.B., M.W. Reed, T. Cox, J.S. Virosco and A.D. Adams et al., 1993. Facile preparation of nuclease resistant 3' modified oligodeoxynucleotides. Nucleic Acids Res., 21: 145-150.
PubMed - Jonas, J.B., I. Akkoyun, W.M. Budde, I. Kreissig and R.F. Degenring, 2004. Intravitreal reinjection of triamcinolone for exudative age-related macular degeneration. Arch. Ophthalmol., 122: 218-222.
PubMed - Kim, J.H., H.O. Jun, Y.S. Yu, B.H. Min, K.H. Park and K.W. Kim, 2010. Protective effect of clusterin from oxidative stress-induced apoptosis in human retinal pigment epithelial cells. Invest. Ophthalmol. Vis. Sci., 51: 561-566.
PubMed - Cheng, K., N.N. Yang and R.I. Mahato, 2009. TGF-β 1 gene silencing for treating liver fibrosis. Mol. Pharm., 6: 772-779.
PubMed - Liu, Y., 2004. Hepatocyte growth factor in kidney fibrosis: Therapeutic potential and mechanisms of action. Am. J. Physiol. Renal. Physiol., 287: F7-F16.
PubMedDirect Link - Hirasawa, M., K. Noda, S. Noda, M. Suzuki and Y. Ozawa et al., 2011. Transcriptional factors associated with epithelial-mesenchymal transition in choroidal neovascularization. Mol. Vis., 17: 1222-1230.
Direct Link - Ciccone, M.M., F. Cortese, M. Gesualdo, S. Carbonara and A. Zito et al., 2013. Dietary intake of carotenoids and their antioxidant and anti-inflammatory effects in cardiovascular care. Mediators Inflammation, Vol. 2013.
CrossRefDirect Link - Matsuda, Y., K. Matsumoto, A. Yamada, T. Ichida and H. Asakura et al., 1997. Preventive and therapeutic effects in rats of hepatocyte growth factor infusion on liver fibrosis/cirrhosis. Hepatology, 26: 81-89.
CrossRefDirect Link - Matsumoto, K. and T. Nakamura, 2001. Hepatocyte growth factor: Renotropic role and potential therapeutics for renal diseases. Kidney Int., 59: 2023-2038.
CrossRefDirect Link - Mochizuki, M., S. Sugita and K. Kamoi, 2013. Immunological homeostasis of the eye. Prog. Ret. Eye Res., 33: 10-27.
CrossRef - Ehrlich, R., A. Harris, N.S. Kheradiya, D.M. Winston, T.A. Ciulla and B. Wirostko, 2008. Age-related macular degeneration and the aging eye. Clin. Invest. Aging, 3: 473-482.
PubMed - Naylor, R.M., D.J. Baker and J.M. van Deursen, 2013. Senescent cells: A novel therapeutic target for aging and age-related diseases. Clin. Pharmacol. Ther., 93: 105-116.
CrossRef - Katta, S., I. Kaur and S. Chakrabarti, 2009. The molecular genetic basis of age-related macular degeneration: An overview. J. Genet., 88: 425-449.
PubMed - Saika, S., O. Yamanaka, K. Ikeda, S. Kim-Mitsuyama and K.C. Flanders et al., 2005. Inhibition of p38MAP kinase suppresses fibrotic reaction of retinal pigment epithelial cells. Lab. Invest., 85: 838-850.
CrossRef - Wynn, T.A., 2011. Integrating mechanisms of pulmonary fibrosis. J. Exp. Med., 208: 1339-1350.
CrossRefDirect Link - Ueki, T., Y. Kaneda, H. Tsutsui, K. Nakanishi and Y. Sawa et al., 1999. Hepatocyte growth factor gene therapy of liver cirrhosis in rats. Nat. Med., 5: 226-230.
CrossRefDirect Link - Wang, X.F., J. Cui, S.S. Prasad and J.A. Matsubara, 2005. Altered gene expression of angiogenic factors induced by calcium-mediated dissociation of retinal pigment epithelial cells. Invest. Ophthalmol. Vis. Sci., 46: 1508-1515.
PubMed - Xu, W., L.W. Wang, J.Z. Shi and Z.J. Gong, 2009. Effects of RNA interference targeting transforming growth factor-beta 1 on immune hepatic fibrosis induced by Concanavalin A in mice. Hepatobiliary Pancreat. Dis. Int., 8: 300-308.
Direct Link - Xie, W., W. Yu, M. Zhao, W. Zhou and H. Chen et al., 2011. Protective effect of paeoniflorin against oxidative stress in human retinal pigment epithelium in vitro. Mol. Vis., 17: 3512-3522.
PubMed - Qazi, Y., S. Maddula and B.K. Ambati, 2009. Mediators of ocular angiogenesis. J. Genet., 88: 495-515.
CrossRefDirect Link