
Cemaluk C. Egbuonu Anthony
Department of Biochemistry, Michael Okpara University of Agriculture, Umudike, Nigeria
Ambrose E. Ogbu
Department of Biochemistry, University of Nigeria Nsukka, Nigeria
Lawrence U.S. Ezeanyika
Department of Biochemistry, University of Nigeria Nsukka, Nigeria
Asian Journal of Biochemistry
Year: 2015 | Volume: 10 | Issue: 2 | Page No.: 67-77







ABSTRACT
The study assessed the effect on some serum functions and organ histology following esculetin exposure in rats. Five groups (n = 6) of Wistar rats respectively received (A) 6.0 mg kg-1 body weight (b.wt.), (B) 12.0 mg kg-1 b.wt. (C) 24.0 mg kg-1 b.wt. of esculetin, (D, vehicle control) 0.2 mL kg-1 b.wt. of 10% dimethylsulfoxide, DMSO and (E, normal control) 0.2 mL kg-1 b.wt. of Distilled Water (DW) per orally and daily for 28 days. On comparison with the controls, a dose dependent and significant (p<0.05) decrease in the concentration of serum sodium ion (141.00±5.02 mmol L-1) but an increase in that of potassium ion (9.39±3.24 mmol L-1) was observed in the group C rats. The decrease in the serum protein concentration (6.58±0.77 mmol L-1) in the group C rats, though dose dependent, was not significant (p>0.05). In the group C rats, the decrease (p<0.05) in the serum urea concentration (10.95±0.61 mmol L-1) was dose dependent but the increase (p<0.05) in the serum creatinine concentration (85.87±8.31 mmol L-1) was not dose dependent. Histological examination revealed a severely dilated hepatic vein (A) and proliferated cells around the bile canaliculi (B) in group C rats. Other organs examined (kidney, testis, brain and heart) did not show any lesion in the test groups. The results showed that the esculetin-induced improvement in the serum parameters might not alter the studied organ histology in the rats, especially at the lower tested concentrations.
PDF Abstract XML References Citation
Received: January 04, 2015;
Accepted: March 24, 2015;
Published: June 04, 2015
How to cite this article
Cemaluk C. Egbuonu Anthony, Ambrose E. Ogbu and Lawrence U.S. Ezeanyika, 2015. Sub-Chronic Oral Esculetin (6, 7-Dihydroxy-Coumarin) Exposure in Male Wistar Rats: Effect on Some Serum Functions and Organ Histology. Asian Journal of Biochemistry, 10: 67-77.
DOI: 10.3923/ajb.2015.67.77
URL: https://scialert.net/abstract/?doi=ajb.2015.67.77
DOI: 10.3923/ajb.2015.67.77
URL: https://scialert.net/abstract/?doi=ajb.2015.67.77
INTRODUCTION
Coumarins, a group of non-nutrient phenolic phytochemicals (Leung et al., 2005; Kostova, 2005), including umbelliferone (7-hydroxy coumarin), scopoletin (7-hydroxyl-6-methoxy coumarin) and esculetin (6,7- dihydroxycoumarin), are present in plants and in foodstuffs (Masamoto et al., 2003; Tien et al., 2011). They are widely used as flavour-enhancing agents (Leung et al., 2005) and in the synthesis of, especially anti-cancer, drugs (Kostova, 2005).
Coumarins possess diverse biological and pharmaceutical properties, including anti-edema, anti-inflammatory and anti-tumour activities (Hoult and Paya, 1996). In particular, protective effect of esculetin against oxidative stress (Kaneko et al., 2003; Tahara et al., 2005; Kim et al., 2008), cancer/proliferative activity (Lee et al., 2013) and inflammation (Kim et al., 2014) have been reported. These reports may in particular increase the use of esculetin in diets and drugs even above the approximate content of mixed coumarins in western diet reported by Lacy and O’Kennedy (2004).
Earlier, Ezeanyika and Egbuonu (2011) recognized the importance of dietary intervention in managing chronic diseases but effect, especially following repeated use and possible overuse need to be studied. This is because response from such agent in animals may vary (Cemaluk and Osakwe, 2011; Egbuonu et al., 2012; Egbuonu and Ezeanyika, 2013). The reported benefits of esculetin (6,7-dihydroxy-coumarin), including protection against oxidative stress (Kim et al., 2008) and cancer (Lee et al., 2013) may increase its use in diets and drugs with varied effects. These warranted the present study of the effect of esculetin on the concentration of serum sodium ion, potassium ion, urea and creatinine which are some bio-indicators of renal, cellular and (to some extent) cardiac functions in animals.
The maintenance of the physiological concentration of sodium and potassium ions by the Na+/K+ pump (Sweadner and Goldin, 1980; Cantley, 1981) is crucial for cells and organs viability. The serum concentration of Na+ and K+ aside indicating renal function may indicate neurological and cardiac functions (Bush, 1991) and presence or otherwise of high blood pressure (Jaitovich and Bertorello, 2010). Serum concentration of urea and creatinine indicated state of renal function (Bush, 1991) and oxidative stress status (D’Apolito et al., 2010). In particular, low serum concentration of creatinine strongly predicted type 2 diabetes mellitus in human (Harita et al., 2009; Hjelmesaeth et al., 2010). Furthermore, organ histology has been used to assess the effect of agents in animals (Egbuonu and Ezeanyika, 2013; Cemaluk et al., 2013). Thus, the study aimed to assess the effect on some serum functions and organ histology following esculetin exposure in rats. The objectives set to achieve the stated aim included the assessment of the effect of esculetin exposure on the concentration of some parameters (Na+, K+, urea, creatinine, protein) in the serum and the histology of some organs (liver, kidney, testis, brain and heart) in Wistar rats.
MATERIALS AND METHODS
Thirty male Wistar rats (age, ten-twelve weeks; average body weight, 114.8 g) were used in this study. The rats were purchased from the animal house of the Faculty of Biological Science, University of Nigeria, Nsukka, Nigeria.
Chemicals: Chemicals used in this research are products of May and Baker, England; Merck, Germany; BDH Chemicals, UK and Sigma Chemical Compound, USA. Reagents used were commercial kits and are products of Randox Laboratories Ltd., UK.
Experimental design: The rats were housed in separate cages, acclimatized for seven days and then randomly assigned to five groups (A, B, C, D and E) of six rats each. The rats in group A, B and C were given esculetin solution at a dose of 6.0, 12.0 and 24.0 mg kg-1 b.wt., respectively while those in group D received 0.2 mL of 10% dimethylsulfoxide (DMSO) as vehicle control whereas those in group E were given 0.2 mL of Distilled Water (DW) as normal control. The administration was per oral for twenty eight consecutive days. All the animals were ad libitum fed pelletised growers feed manufactured by Grand Cereals and Oil Mills limited, Jos, Nigeria.
Methods: The determination of the concentration of serum sodium ion and potassium ion was by flame photometry. Serum sample was nebulized (sprayed as a fine mist of droplets using compressed air) into a non-luminous gas flame to give light with a characteristic colour at a corresponding wavelength for sodium ion (589 nm) or potassium ion (767 nm). The characteristic light was filtered and the amount of light emitted which depends on the concentration of metallic ion present was measured on falling on a photosensitive detector system and compared with a standard.
Urea concentration in the serum was determined, using Randox enzyme kit based on the method of Weatherburn (1967) and on the principle that urea in the serum is hydrolyzed to ammonia in the presence of urease. The resultant ammonia reacts with hypochlorite and phenol (Berthelot’s reaction) to yield blue coloured indophenols that is measured by a spectrophotometer:
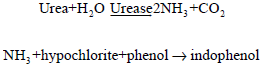 |
Creatinine concentration in the serum was determined using Randox enzyme kit by the method of Slot (1965). This method was based on the principle that creatinine reacts with picric acid in an alkaline medium to form a coloured complex which is measured with a spectrophotometer.
The serum total protein was determined by the method of Doumas (1975) based on the principle that cupric ions in alkaline medium interact with protein peptide bonds resulting in the formation of a coloured complex with intensity proportional to the amount of protein.
Collection of organs: Sample of the liver, kidney, heart, testis and brain were taken from the respective group on sacrifice and fixed in 10% formalin. The respective sample was washed with ethanol and xylene and embedded in paraffin wax. Sections (5 μm thick) were cut, stained in heamatoxylin and eosin and studied under the light microscope.
Statistical analysis: The data was analyzed by the Least Significant Difference (LSD) and significant difference in means accepted at p<0.05, using one way analysis of variance (ANOVA). The results were expressed as Mean value±Standard Deviation (SD) of the measured variables in each group.
RESULTS
Serum protein concentration: The result as presented in Fig. 1 showed that the serum protein concentration decreased in the test groups as compared with the normal control. The least serum protein concentration (6.58±0.77 mmol L-1) was recorded in the group of rats that received the highest concentration of esculetin (24.0 mg kg-1 b.wt.). The decrease, however was not significant (p>0.05).
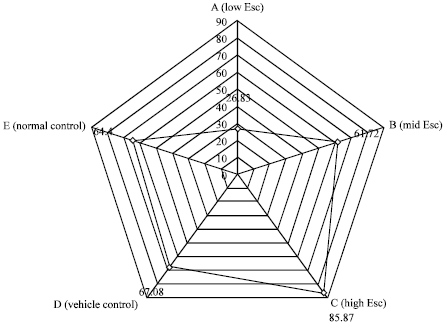
Serum sodium ion concentration: When compared with the controls, the serum sodium ion concentration decreased as the concentration of esculetin increased (Fig. 2). The decrease in serum sodium ion concentration observed in the rats given esculetin at 12.0 mg kg-1 b.wt. (154.00±18.07 mmol L-1) and 24.0 mg kg-1 b.wt. (141.00±5.02 mmol L-1) was significant (p<0.05) whereas that in the group of rats exposed to 6.0 mg kg-1 b.wt. of esculetin (158.00±15.95 mmol L-1) was not significant (p>0.05).
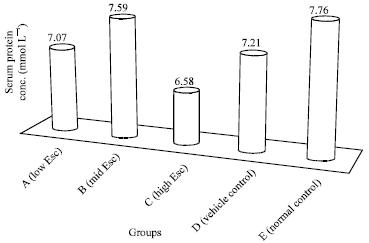 | |
| Fig. 1: | Effect of esculetin on the serum protein concentration of rats |
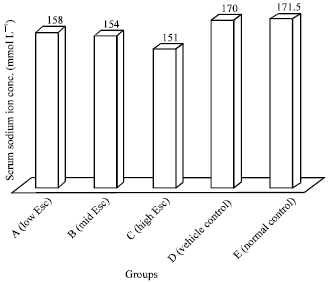 | |
| Fig. 2: | Effect of esculetin on the serum sodium ion concentration of rats |
Serum potassium ion concentration: Contrary to the controls, the serum potassium ion concentration in the test group of rats increased in a dose dependent manner (Fig. 3). However, the observed increase in the rats that received 12.0 mg kg-1 b.wt. (7.43±0.48 mmol L-1) and 24.0 mg kg-1 b.wt. (9.39±3.24 mmol L-1) but not 6.0 mg kg-1 b.wt. (6.85±1.85 mmo L-1), was statistically significant (p<0.05).
Serum urea concentration: The result in Fig. 4 showed a dose dependent and significant (p>0.05) decrease in the serum urea concentration in the test groups as against the controls. The least serum urea concentration (10.95±0.61 mmol L-1) was recorded in the group of rats that were given 24 mg kg-1 b.wt. of esculetin.
Serum creatinine concentration: The serum creatinine concentration decreased in the groups of rats that received 6.0 mg kg-1 b.wt. (26.83±8.31 μmol L-1) and 12.0 mg kg-1 b.wt. (61.72±6.57 μmol L-1) of esculetin but increased in the group of rats that received 24.0 mg kg-1 b.wt. (85.87±8.31 μmol L-1) of esculetin (Fig. 5).
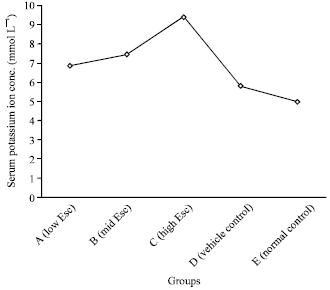 | |
| Fig. 3: | Effect of esculetin on the serum potassium ion concentration of rats |
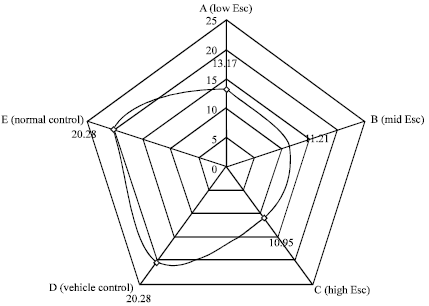 | |
| Fig. 4: | Effect of esculetin on the serum urea concentration of rats |
However, the observation in groups A (low Esc) and C (high Esc) but not B (mid Esc), was statistically significant (p<0.05) when compared with the controls. The result show an increasing trend in the serum creatinine concentration from the least (6.0 mg kg-1 b.wt.) to the highest (24.0 mg kg-1 b.wt.) dosed rats.
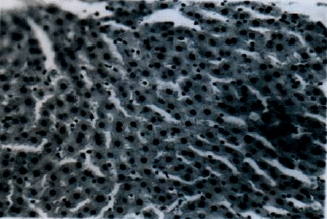
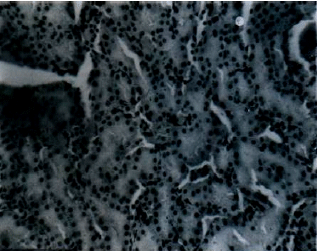
Histology of some organs: Liver section of rats given 24.0 mg kg-1 b.wt. of esculetin as presented in Fig. 6 shows severely dilated hepatic vein (V) and proliferation of cells around the bile canaliculi (B). A section of the liver from rats in Groups E (normal control), D (vehicle control), A and B showed no lesion (Fig. 7). The section of the kidney (Fig. 8), testis (Fig. 9), brain (Fig. 10) and heart (Fig. 11) did not show any lesion in all the groups (Group A, B, C, D and E).
 | |
| Fig. 5: | Effect of esculetin on the serum creatinine concentration of rats |
 | |
| Fig. 6: | Liver section of rat given 24.0 mg kg-1 b.wt. of esculetin showing dilated hepatic vein (A) and proliferation of cells around the bile canaliculi (B) (H and E stain x400) |
 | |
| Fig. 7: | Liver section of rats in the other groups showing no lesion (H and E stain x400) |
 | |
| Fig. 8: | Kidney section of rats showing no lesion (H and E stain x400) |
 | |
| Fig. 9: | Testis section of rats showing no lesion (H and E stain x400) |
 | |
| Fig. 10: | Brain section of rats showing no lesion (H and E stain x400) |
 | |
| Fig. 11: | Heart section of rats showing no lesion (H and E stain x400) |
DISCUSSION
Esculetin (6, 7-dihydroxycoumarin), coumarin derivative found in various natural plant products, may have beneficial biological activities. However, there is need to assess the effect of esculetin on organ functions following its repeated use and possible abuse. Thus, the present study investigated the effect of esculetin on some bio-indicators of organ functions in male rats.
The maintenance of the physiological concentration of sodium and potassium ions by the Na+/K+ pump (Sweadner and Goldin, 1980; Cantley, 1981) is crucial for cell viability. Sodium reduction is an important approach in the prevention and treatment of hypertension (Whelton et al., 1998; Sacks et al., 2001). Sodium ion accumulation caused high blood pressure in animals (Jaitovich and Bertorello, 2010) whereas decreased potassium ion (hypokalaemia) resulted to neurological and cardiac dysfunctions in animals (Bush, 1991). The dose dependent and significant (p<0.05) decrease in the concentration of serum sodium ion but an increase in that of potassium ion observed in this study suggests hyponatraemic and hyperkalaemic effects of esculein in the rats. This could be beneficial as the sodium ion concentration in the cytosol need to be lower than that of potassium. The observation could also imply improved renal function. Decreased sodium concentration stimulates osmoreceptors in the hypothalamus to release antidiuretic hormone, ADH (Fereidoun et al., 2007) which conserves water by increasing water re-absorption in the collecting ducts of the kidneys thereby reducing the sodium ion concentration in the blood. On the other hand, increased potassium ion concentration besides enhancing aldosterone secretion, hence renal clearance, could enhance many enzymatic reactions, the transmission of nerve impulse and heart functions (O’Toole, 1997). Thus, the result suggests possible potential of esculetin to improve the concentration of sodium and potassium ions and perhaps the related cellular and organ functions in the rats.
The observed decrease in the protein concentration of rats in this study, though dose dependent, was not significant (p>0.05). This is consistent with the findings of Elliott et al. (2001) and Watanabe et al. (1999), indicating that esculetin did neither inhibit protein synthesis nor enhanced tissue breakdown in the rats. This could ensure steady state in body weight of the rats which appears to support the non-significant change in the body weight of esculetin-exposed rats in a similar study (Ambrose et al., 2012). This may account for the decrease in the serum urea concentration of the rats in this study. Protein catabolism via its building block, amino acids results in the production of urea that is excreted along with urine.
The serum urea concentration decreased (p<0.05) dose dependently but the decrease (p<0.05) in the serum creatinine concentration was not dose dependent. Decreased concentration of creatinine and urea in serum indicated an improved functional capacity of the kidneys to filter fluids (Bagga et al., 2005). Further, the decreased urea concentration may suggest a reduced oxidative stress in the rats (D’Apolito et al., 2010). In apparent support of the result of this study, Ramesh et al. (2007) reported that umbelliferone (7-hydroxycoumarin), a coumarin derivative decreased urea and creatinine concentration in rats’ serum. The result shows an increasing trend in the serum creatinine concentration from the least (6.0 mg kg-1 b.wt.) to the highest (24.0 mg kg-1 b.wt.) dosed rats (Fig. 5). This indicates apparent esculetin-induced benefit on the renal function at a lower concentration. Creatinine is a nitrogenous waste product formed from the metabolism of creatine in the skeletal muscle (Murray et al., 2003). The significant increase in creatinine concentration but not in urea concentration, in rats’ serum that received 24.0 mg kg-1 b.wt. of esculetin may be indicating enhanced muscular dystrophy, perhaps following increased creatine metabolism in the skeletal muscle (Murray et al., 2003).
Histological examination revealed a severely dilated hepatic vein (A) and proliferated cells around the bile canaliculi (B) of liver sections in group C rats (Plate 1). These are common forms of liver reaction to toxic conditions but do not agree with the observation of Lin et al. (2000) that esculetin reduced the incidence of liver lesions induced by t-butyl hydroperoxide (t-BHP). However, the dose (0.5 and 5.0 mg kg-1), duration (5 days) and route of administration (intraperitoneal) used in Lin et al. (2000) differed from what was used in this study. All other organs examined (kidney, testis, brain and heart) did not show any lesion in all the groups treated with esculetin (Plate 2, 3, 4, 5 and 6). This agrees with the serum results in this study and in a similar study reported earlier (Egbuonu et al., 2012). The results of the histological examination of organs suggest that esculetin at higher dose may impair the liver histology whereas the administration of esculetin at a dose of 6.0 and 12.0 mg kg-1 may have no effect on the studied organ histology of rats.
CONCLUSION
The study showed that the esculetin-induced improvement in serum parameters might not alter the studied organ histology in the rats, especially at the lower tested concentrations. The implication of the result with muscular dystrophy underscores the need to study the effect of esculetin on the muscle of animals.
REFERENCES
- Bagga, A., A. Bajpai and S. Menon, 2005. Approach to renal tubular disorders. Indian J. Pediatr., 72: 771-776.
CrossRefDirect Link - Doumas, B.T., 1975. Standards for total serum protein assays-A collaborative study. Clin. Chem., 21: 1159-1166.
PubMedDirect Link - D'Apolito, X. Du, H. Zong, A. Catucci and L. Maiuri et al., 2010. Urea-induced ROS generation causes insulin resistance in mice with chronic renal failure. J. Clin. Invest., 120: 932-932.
CrossRefDirect Link - Egbuonu, A.C.C., A.E. Ogbu and L.U.S. Ezeanyika, 2012. Dose-related influence of esculetin (6,7-Dihydroxy-coumarin) on some liver and prostate function markers of male Wistar rats. J. Biol. Sci., 12: 253-257.
CrossRefDirect Link - Egbuonu, A.C.C. and L.U.S. Ezeanyika, 2013. L-arginine exposure improves renal function markers of metabolic syndrome in female rats. Am. J. Biochem. Mol. Biol., 3: 50-60.
CrossRefDirect Link - Egbuonu, A.C.C. and O.N. Osakwe, 2011. Effects of high monosodium glutamate on some serum markers of lipid status in male Wistar rats. J. Med. Med. Sci., 2: 653-656.
Direct Link - Cemaluk, E.A.C., E.P. Madus, L.U.S. Ezeanyika and O. Obidoa, 2013. Combined oral arginine and monosodium glutamate exposure induces adverse response on the prostate function and testis histology of rats. Br. J. Pharm. Res., 3: 247-258.
CrossRefDirect Link - Elliott, S., A.D. Rowan, S. Carrere, P. Koshy, J.B. Catterall and T.E. Cawston, 2001. Esculetin inhibits cartilage resorption induced by interleukin 1α in combination with oncostatin M. Ann. Rheum. Dis., 60: 158-165.
CrossRefDirect Link - Ezeanyika, L.U.S. and A.C.C. Egbuonu, 2011. Impact of nitric oxide and insulin resistance on the pathophysiology of the metabolic syndrome: Possible role of L-arginine and glutamate. J. Med. Med. Sci., 2: 657-662.
Direct Link - Fereidoun, H., A. Bahram, K.S. Sadraddin, A. Abbas and H. Pouria, 2007. Function of ADH and thirst mechanisms in the face of consuming salty water in male Wistar rat. Pak. J. Physiol., 3: 1-4.
Direct Link - Harita, N., T. Hayashi, K.K. Sato, Y. Nakamura, T. Yoneda, G. Endo and H. Kambe, 2009. Lower serum creatinine is a new risk factor of type 2 diabetes: The Kansai healthcare study. Diabetes Care, 32: 424-426.
CrossRefPubMedDirect Link - Hjelmesaeth, J., J. Roislien, N. Nordstrand, D. Hofso, H. Hager and A. Hartmann, 2010. Low serum creatinine is associated with type 2 diabetes in morbidly obese women and men: A cross-sectional study. BMC Endocrine Disorders, Vol. 10.
CrossRefDirect Link - Hoult, J.R.S. and M. Paya, 1996. Pharmacological and Biochemical actions of simple coumarins: Natural products with therapeutic potential. Gen. Pharmacol.: Vasc. Syst., 27: 713-722.
CrossRefDirect Link - Jaitovich, A. and A.M. Bertorello, 2010. Intracellular sodium sensing: SIK1 network, hormone action and high blood pressure. Biochimica Biophysica Acta, 1802: 1140-1149.
CrossRefPubMedDirect Link - Kaneko, T., N. Baba and M. Matsuo, 2003. Protection of coumarins against linoleic acid hydroperoxide-induced cytotoxicity. Chem. Biol. Interact., 142: 239-254.
CrossRefDirect Link - Kim, S.H., K.A. Kang, R. Zhang, M.J. Piao and D.O. Ko et al., 2008. Protective effect of esculetin against oxidative stress-induced cell damage via scavenging reactive oxygen species. Acta Pharmacologica Sinica, 29: 1319-1326.
CrossRefDirect Link - Kim, Y., Y. Park, S. Namkoong and J. Lee, 2014. Esculetin inhibits the inflammatory response by inducing heme oxygenase-1 in cocultured macrophages and adipocytes. Food Funct., 5: 2371-2377.
CrossRefDirect Link - Kostova, I., 2005. Synthetic and natural coumarins as cytotoxic agents. Curr. Med. Chem. Anti-Cancer Agents, 5: 29-46.
CrossRefDirect Link - Lacy, A. and R. O'Kennedy, 2004. Studies on coumarins and coumarin-related compounds to determine their therapeutic role in the treatment of cancer. Curr. Pharm. Des., 10: 3797-3811.
CrossRefPubMedDirect Link - Lee, S.Y., T.G. Lim, H. Chen, S.K. Jung and H.J. Lee et al., 2013. Esculetin suppresses proliferation of human colon cancer cells by directly targeting β-catenin. Cancer Prev. Res., 6: 1356-1364.
CrossRefPubMedDirect Link - Leung, K.N., P.Y. Leung, L.P. Kong and P.K. Leung, 2005. Immunomodulatory effects of esculetin (6,7-dihydroxycoumarin) on murine lymphocytes and peritoneal macrophages. Cell. Mol. Immunol., 2: 181-188.
PubMedDirect Link - Lin, W.L., C.J. Wang, Y.Y. Tsai, C.L. Liu, J.M. Hwang and T.H. Tseng, 2000. Inhibitory effect of esculetin on oxidative damage induced by t-butyl hydroperoxide in rat liver. Arch. Toxicol., 74: 467-472.
CrossRefPubMedDirect Link - Masamoto, Y., H. Ando, Y. Murata, Y. Shimoishi, M. Tada and K. Takahata, 2003. Mushroom tyrosinase inhibitory activity of esculetin isolated from seeds of Euphorbia lathyris L. Biosci. Biotechnol. Biochem., 67: 631-634.
CrossRefPubMedDirect Link - Ambrose, E.O., C.C.E. Anthony and U.S.E. Lawrence, 2012. Time and dose dependent effects of esculetin on some routine parameters of biochemical function in male Wistar rats. J. Biochem. Bioinform., 2: 105-108.
Direct Link - Ramesh, B., P. Viswanathan and K.V. Pugalendi, 2007. Protective effect of umbelliferone on membranous fatty acid composition in streptozotocin-induced diabetic rats. Eur. J. Pharmacol., 566: 231-239.
CrossRefDirect Link - Sacks, F.M., L.P. Svetkey, W.M. Vollmer, L.J. Appel and G.A. Bray et al., 2001. Effects on blood pressure of reduced dietary sodium and the Dietary Approaches to Stop Hypertension (DASH) diet. N. Engl. J. Med., 344: 3-10.
CrossRefPubMedDirect Link - Slot, C., 1965. Plasma creatinine determination: A new and specific Jaffe reaction method. Scand. J. Lab. Invest., 17: 381-387.
CrossRefPubMedDirect Link - Sweadner, K.J. and S.M. Goldin, 1980. Active transport of sodium and potassium ions: Mechanism, function and regulation. N. Engl. J. Med., 302: 777-783.
PubMed - Tien, Y.C., J.C. Liao, C.S. Chiu, T.H. Huang, C.Y. Huang, W.T. Chang and W.H. Peng, 2011. Esculetin ameliorates carbon tetrachloride-mediated hepatic apoptosis in rats. Int. J. Mol. Sci., 12: 4053-4067.
CrossRefDirect Link - Watanabe, K., A. Ito, T. Sato, T. Saito, H. Hayashi and Y. Niitani, 1999. Esculetin suppresses proteoglycan metabolism by inhibiting the production of matrix metalloproteinases in rabbit chondrocytes. Eur. J. Pharmacol., 370: 297-305.
CrossRefDirect Link - Weatherburn, M.W., 1967. Phenol-hypochlorite reaction for determination of ammonia. Anal. Chem., 39: 971-974.
CrossRefDirect Link - Whelton, P.K., L.J. Appel, M.A. Espeland, W.B. Applegate and W.H. Ettinger Jr. et al., 1998. Sodium reduction and weight loss in the treatment of hypertension in older persons: A randomized controlled trial of nonpharmacologic interventions in the elderly (TONE). J. Am. Med. Assoc., 279: 839-846.
CrossRefDirect Link