
Rajeeva Gaur
Department of Microbiology (Centre of Excellence), Dr. Ram Manohar Lohia Avadh University, Faizabad, 224001, Uttar Pradesh, India
Soni Tiwari
Department of Microbiology (Centre of Excellence), Dr. Ram Manohar Lohia Avadh University, Faizabad, 224001, Uttar Pradesh, India
Sheffalika Singh
Department of Microbiology (Centre of Excellence), Dr. Ram Manohar Lohia Avadh University, Faizabad, 224001, Uttar Pradesh, India
Asian Journal of Biochemistry
Year: 2015 | Volume: 10 | Issue: 2 | Page No.: 52-66







ABSTRACT
Fifteen bacterial strains isolated from the dairy soil samples for acidic protease production. Among them, culture RGSS-09 was adjudged as the best protease producer and was identified as Pseudomonas aeruginosa RGSS-09 from MTCC Chandigarh. The isolate showed maximum protease production (1256 U mL-1) with 2.5 g L-1 biomass production in the presence of wheat bran and yeast extract at 37°C pH 6.5 within 48 h of incubation. Pseudomonas aeruginosa RGSS-09 also showed enhanced activity and stability in presence of Ca, Mg, Tween-20, Tween-40, triton-X-100 and SDS at 5 and 10 mM concentration. The enzyme was also stable in the presence of n-dodecane, isooctane, n-decane, xylene, toluene, n-haxane, n-butanol and cyclohexane, respectively. The presence of benzene, methanol and ethanol marginally reduced the protease stability, respectively. The enzyme was showed it 100% activity at 75°C and pH 7.5 with 110 and 100% stability at 70°C and pH 7.0, respectively. The purity of the enzyme was confirmed by the presence of a single band on SDS-PAGE and its molecular weight was approximately 42 kDa. Pseudomonas aeruginosa can be used profitably for the large scale production of protease to meet the present day demand of the industrial sector.
PDF Abstract XML References Citation
Received: March 15, 2015;
Accepted: May 23, 2015;
Published: June 04, 2015
How to cite this article
Rajeeva Gaur, Soni Tiwari and Sheffalika Singh, 2015. Production and Characterization of Thermotolerant-Organic Solvent Resistant Acidic Protease by Pseudomonas aeruginosa RGSS-09 Isolated from Dairy Sludge. Asian Journal of Biochemistry, 10: 52-66.
DOI: 10.3923/ajb.2015.52.66
URL: https://scialert.net/abstract/?doi=ajb.2015.52.66
DOI: 10.3923/ajb.2015.52.66
URL: https://scialert.net/abstract/?doi=ajb.2015.52.66
INTRODUCTION
Proteases are one of the industrially important enzymes, have been in use for many centuries, at first in the dairy industry as milk-clotting agents (rennet) for the manufacture of cheese. Proteases comprise a large and complex group of enzymes which vary in properties such as substrate specificity, active site and catalytic mechanism, pH and temperature optima and enzyme stability. The specificity of proteolytic enzymes is directed by the nature of the amino acid and other functional groups (aromatic or aliphatic or sulphur-containing) close to the bond being hydrolyzed. Proteases are present in all living things and play a significant role in regular and irregular physiological environment, catalyzing different metabolic reactions (Sandhya et al., 2004).
Proteases classified into two major groups based on their ability to cleave N-or C-terminal peptide bonds (exopeptidases) or internal peptide bonds (endopeptidases). Proteases are also distinguished by the presence or absence of charged groups in positions relative to the susceptible bond (Ward, 1985) and are classified on a number of bases: their pH optima (as acidic, neutral or alkaline); substrate specificity (collagenase, keratinase, elastase, etc.) or their homology to well-studied proteins such as trypsin, pepsin, etc. (trypsin-like, pepsin-like, etc.). Protease are employed in the dehairing and bating of hides, meat tenderizing, cheese flavour development, treatment of flour in the manufacture of baked goods, improvement of dough texture, flavor and color in cookies, etc., improving digestibility of animal feeds, reducing viscosity of fish press water, recovery of fish oils, silk degumming and proteinaceous stain removal in fabrics, etc. (Adinarayana and Ellaiah, 2002; Maase and van Tilburg, 1983; Anisworth, 1994; Outtrup et al., 1995; Wolff et al., 1996; Inhs et al., 1998).
Acidic proteases are the most interesting group of proteases with respect to apply in the food industry. They have maximum activity and stability at pH 2.0-6.5. They are resistant to SH-reagents, metal chelators, heavy metals and DFP, but are rapidly inactivated at higher pH values. The acid proteases exhibit limited esterase activity but split a wide range of peptide bonds (Ikram-ul-Haq and Mukhtar, 2007). The estimated value of the worldwide sales of industrial enzymes is $1 million a year, with proteases accounting for about 60% of it (Godfrey and West, 1996). Latest progresses in enzyme technology have seen the usage of acidic, neutral and alkaline proteases in their relevant industry as well as the application of their recombinant shape. Proteases are achieved from plants, animals and microorganisms, with the majority attained from microbial resources. Presently, a bulk quantity of commercially available proteases is derived from bacteria and fungi. The ideal sources of proteases are bacteria because of their rapid growth and the ease with which they can be genetically manipulated to manufacture new enzymes with altered properties and used at industry level (Rao et al., 1998). The aim of present study was to isolation and characterization of potential acidic protease producing microorganism from the natural ecosystem.
MATERIALS AND METHODS
Isolation, screening and identification of protease producing bacteria: The soil samples were collected aseptically from different site of dairy which are potentially good sources of protease producing micro-organism. One gram soil was suspended in 9.0 mL sterile distilled water, agitated for a min and 0.1 mL suspension was spread over skimmed milk agar plates (pH 7.0) containing 10.0% skimmed milk, 0.1% potassium chloride, 0.2% MgSO4 and 2.0% agar at 37°C. Bacterial colonies showing clear zones were selected, streaked twice on skimmed milk agar plates for purification and maintained as pure culture over nutrient agar slants (pH 7.0, 4°C). Bacterial isolates having maximum protease activity, as measured by caseinolytic zone diameter (mm) (Ellaiah et al., 2002). The isolate having maximum clearance zone was selected for further studies. The selected bacterial isolate was identified by morphological and biochemical characterization as per the Bergey’s Manual of Systematic Bacteriology (Krieg and Holt, 1984).
Enzyme assay: The production of acidic protease was carried out in the broth production medium. One milliliter of the overnight grown bacterial strain was inoculated in 100 mL of the production medium containing: Glucose 0.5%, peptone 0.5%, yeast extract 0.1%, K2HPO4 0.1%, KH2PO4 0.1% and MgSO47H2O 0.05% at pH 6.5. After incubation, the medium was centrifuged at 10,000 rpm for 10 min. Supernatant (culture filtrate) was used as the source of crude enzyme. Culture filtrate (1 mL) was added with 1ml of 1% (w/v) casein solution in phosphate buffer of pH 7.0 and incubated for 30 min at 37°C. The reaction was stopped by addition of 3 mL of 10% trichloroacetic acid (TCA solution). The reaction mixture was filtered with whatman filter paper and collect supernatant. The amount of tyrosine released was determined UV-Spectrophotometrically at 280 nm against the enzyme blank. One unit of protease activity was equivalent to amount of enzyme required to release 1 g of tyrosine mL-1 min-1 under standard assay conditions.
Preparation of inoculum: A volume of 25 mL of protease broth taken in a 100 mL Erlenmeyer flask was inoculated with a loop full of cells from a 24 h old slant and kept at 37°C. After overnight of incubation, 1 mL of this culture was used as the inoculum.
Optimization of physico-chemical and nutritional parameters for acidic protease production: The various process parameters influencing acidic protease production were optimized individually and independently of the others. Therefore, the optimized conditions were subsequently used in all the experiments in sequential order. For optimization, the basal medium (glucose 2.0%, peptone 0.3%, yeast extract 0.2%, MgSO4 0.5% and pH 7.0) was inoculated and incubated at different temperature viz., 25, 35, 37, 40, 45, 50 and 55°C under the standard assay conditions. The samples were withdrawn at every 12 h interval up to 96 h to study the effect of incubation period. The influence of pH on the enzyme production was determined by measuring the enzyme activity at varying pH values ranging from 4.0-9.0 at 37°C using different suitable buffers at concentration of 100 mM acetate buffer (pH 4.0-6.0), phosphate buffer (pH 6.5-8.0) and Tris-HCl buffer (pH 8.0-9.0) under standard assay conditions. The growth medium was supplemented with different carbon sources viz., fructose, sucrose, maltose, starch, glucose, lactose, wheat bran, rice bran, rice husk and maize bran (at the level of 2%, w/v). Different organic nitrogen sources (beef extract, skimmed milk, casein, malt extract, peptone and yeast extract, 0.5% w/v) and inorganic nitrogen sources (ammonium nitrate, ammonium chloride, potassium nitrate and ammonium sulphate 0.5%, w/v) were also used for enzyme production.
Characterization of crude enzyme
Effect of temperature on protease activity and stability: The influence of temperature on activity of protease was studied by incubating the reaction mixture at different temperatures (25-95°C). The enzyme was incubated at different temperatures 25-100°C for 1 h to study the stability of the enzyme. The residual protease activity was measured by conducting the reaction at temperature 37°C and pH 6.0. The activity of the enzyme was considered as 100% under standard assay conditions.
Effect of pH on protease activity and stability: The effect of pH on protease activity was measured in the pH range of 4.0-11.0, using the appropriate buffers at concentration of 100 mM (4.0-6.0, sodium acetate; 6.0-8.0, sodium phosphate; 8.0-10.0, Tris-HCl and 9.0-11.0, glycine-NaOH) under standard assay conditions. To study stability as a function of pH, 100 μL of the crude enzyme was mixed with 100 μL of the buffer solutions and incubated at 37°C for 1 h then aliquots of the mixture were taken to measure the residual protease activity (%) under standard assay conditions.
Effect of organic solvents on protease stability: Cell free supernatant having maximum protease activity was filtered with nitrocellulose membrane (pore size 0.22 μm) and incubated with 25% (v/v) of different organic solvents viz., n-dodecane, n-decane, isooctane, n-octane, xylene, n-hexane, n-butanol, cyclohexane, n-heptane, benzene, toluene, ethanol, methanol and propanol for 7 days in screw crapped tubes at 37°C and 120 rpm. The residual protease activity was estimated against the control, in which solvent was not present.
Effect of metal ions on protease activity and stability: The effect of various metal ions (5 and 10 mM) on enzyme activity was investigated using NaCl, CaCl2, MgCl2, HgCl2, FeSO4, NiCl2, CuSO4, KCl, MnCl2 and ZnSO4. The enzyme was incubated with different metals at 37°C for 1 h to study metal ion stability and assayed under standard assay conditions.
Effect of surfactant on enzyme stability: The effect of various surfactants (1.0%) on enzyme activity was investigated using Triton-X-100, Tween-40, Tween-60, Tween-80 and SDS. The protease sample was incubated with different chemicals for 1 h at 37°C and then the residual activity (%) was tested under standard assay conditions.
SDS-PAGE analysis: The enzyme supernatant, with maximum enzyme activity along with marker was electrophoresed by Sodium Dodecyl Sulphate-Poly Acrylamide Gel Electrophoresis (SDS-PAGE) in a 12.5% polyacrylamide gel according to the method of Laemmli (1970). Approximate molecular weight of the protease was estimated by SDS-PAGE against the molecular mass markers i.e., lysozyme (14.3 kDa), β-lactoglobulin (20 kDa), carbonic anhydrase (29 kDa), ovalbumin (43 kDa), bovine serum albumin (66 kDa) and phosphorylase B (97.4 kDa) (Sigma-Aldrich Pvt Ltd., USA) run with the samples.
Statistical analysis: All experiments were carried out in triplicates and the results are presented as the mean of three independent observations. Standard deviation for each experimental result was calculated using Microsoft Excel.
RESULTS AND DISCUSSION
Isolation, screening and identification of acidic protease producing bacterial cultures: Fifteen bacterial isolates producing variable proteolytic zones on milk agar plates were isolated from the dairy soil samples by serial dilution methods. The zones of clearance by isolates reflect their extent of proteolytic activity. Those having clearance zone greater than >0.9 cm were considered as significant isolate. Among 15 bacterial isolates, one exhibited good proteolytic activity which was reassessed by loading their culture broth in the wells on milk agar plate (pH 7.0). The culture broth of good protease producers cleared more than >0.9 cm zone within 3-4 h of incubation at 37±1°C, thereby indicating an extra-cellular nature of the proteolytic (Fig. 1). The isolate, showing maximum clearance zone diameter was selected for further studies.
The efficient strain RGSS-09 was rod-shaped, gram-negative, aerobe and facultative, motile, with positive urease, catalase and oxidase test. It grew over a wide range of pH (2.0-9.0), temperature (25-65°C), NaCl concentration (0.0-8%) and was able to hydrolyze Gelatin, Tween-20 and 40 and produce acid from glucose. The strain was halotolerant as it grew in the presence of 0.0-8% NaCl. On account of morphological and biochemical characteristics, it was identified as Pseudomonas aeruginosa (Table 1).
Effect of incubation time on protease production: The incubation period for getting the maximum enzyme production is regulated by the characteristics of the isolates (Bhattacharya et al., 2011). Figure 2 shows that maximum enzyme production (721.5 U mL-1) was achieved within 48 h of incubation with 1.98 g L-1 biomass production. Similarly, Samanta et al. (2012) also reported that Pseudomonas aeruginosa showed maximum protease activity within 48 h of incubation time. In another study, Bacillus sp. showed maximum acidic protease production with in 48 h of incubation (Shivakumar, 2012).
 | |
| Fig. 1: | Photographic representation of proteolytic clear zone on skimmed milk |
| Table 1: | Morphological, physiological and biochemical characteristics of the selected isolate |
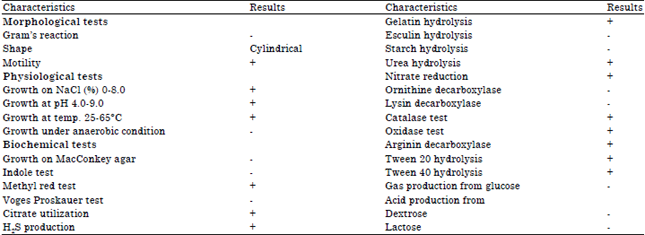 | |
| +: Positive, -Negative | |
Incubation periods play a vital role at which an organism exhibits highest enzyme production while organisms show substantial deviation at different incubation periods (Kumar et al., 2012). In contrast to our result, acidic protease production from Pseudomonas fluorescens was found to be higher at 24 h further increase the incubation periods, enzyme activity gradually decreased (Kalaiarasi and Sunitha, 2009).
Effect of temperature on protease production: The growth and enzyme activity of microorganisms is greatly influenced by different incubation temperatures. The influence of temperature on protease production was investigated from 25-55°C, our isolate P. aeruginosa RGSS-09 showed maximum protease production (779.0 U mL-1) at 37°C with 2.1 g L-1 biomass production. Further, decrease and increases the temperature, decrease the protease production (Fig. 3). Several workers also reported that some bacterial isolates like Pseudomonas aeruginosa MTCC 7926 and Serratia liquefaciens showed maximum protease production at 37°C (Balaji et al., 2012; Smita et al., 2012).
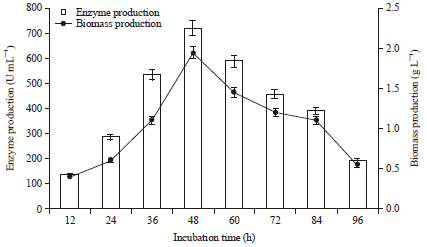 | |
| Fig. 2: | Effect of Incubation periods on protease and biomass production. The flasks were inoculated with culture were incubated at different incubation periods (12-96 h) at initial pH 7.0, 37°C. For enzyme activity the reaction was assayed at respective incubation periods at 37°C for 15 min. Error bars presented mean values of ±Standard deviation of triplicates of three independent experiments |
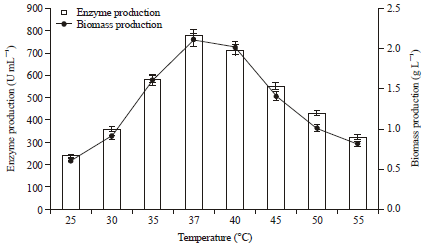 | |
| Fig. 3: | Effect of temperature on protease and biomass production. The flasks were inoculated with culture in the medium were incubated at different temperature (25-55°C) for 48 h at pH 7.0. For enzyme activity reaction mixture was incubated at 37°C for 15 min and reaction was conducted as standard assay method. Error bars presented mean values of ±Standard deviation of triplicates of three independent experiments |
Temperature significantly regulates the synthesis and secretion of bacterial extra-cellular protease by changing the physical properties of the cell membrane (Balaji et al., 2012). Therefore, temperature is a critical parameter that should be controlled in order to obtain an optimum protease production.
Effect of pH on protease production: Among the physical parameters, pH of the growth medium plays a vital role by inducing physiological changes in microbes and their enzyme secretion.
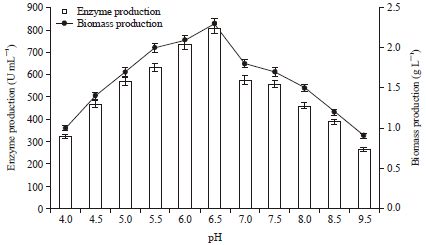 | |
| Fig. 4: | Effect of pH on protease and biomass production. The flasks were inoculated with culture were incubated at different pH (4-9.5) for 48 h at 37°C. For enzyme activity the reaction was assayed at respective pH with buffers (100 mM) at 37°C for 15 min. Error bars presented mean values of ±Standard deviation of triplicates of three independent experiments |
In this experiment, Fig. 4 depicted that protease production was observed over a broad pH range from 4.0-8.0 but maximum protease production (809.0 U mL-1) was reported at pH 6.5 by P. aeruginosa RGSS-09 with 2.3 g L-1 biomass production (Fig. 3). Similarly, Radha et al. (2011) also reported that higher acidic protease production was observed at acidic pH. A gradual increase in protease at pH from 4.0-6.5, whereas, it declined at neutral and alkaline pH. Microbial cells are significantly affected by the pH of their immediate environment because they apparently have no mechanism for adjusting their internal pH. Different organisms have different pH optima and decrease or increase in pH on either side of the optimum value results in poor microbial growth (Bhattacharya et al., 2011). A notable decline in the enzyme productivity occurred at both higher and lower pH values (Qadar et al., 2009). Certain Bacillus species produced protease over the entire range of pH investigated (pH 5.0-10.0) (Josephine et al., 2012).
Effect of carbon sources on protease production: The influence of carbon sources such as wheat bran, soluble starch, fructose, glucose, maltose, lactose and sucrose (2.0%) on protease production from P. aeruginosa RGSS-09 was also studied. Among the carbon sources tested, wheat bran showed maximum (1089.5 U mL-1) protease production with 2.3 g L-1 biomass production by P. aeruginosa RGSS-09 (Fig. 5). Similarly, Mukherjee et al. (2008) reported that maximum protease production was observed in the presence of wheat bran.
Effect of nitrogen source supplementation on protease production: It is well known that nitrogen is metabolized to produce primarily amino acid, nucleic acid, protein and cell wall components and have regulatory effect on the enzyme synthesis. Protease production is highly dependent on both carbon and nitrogen sources existing in the medium (Shanthakumari et al., 2010). The effect of various nitrogen sources on acidic protease production was tested and found that yeast extract reported the highest enzyme production (1256 U mL-1) with 2.5 g L-1 biomass production followed by beef extract from P. aeruginosa RGSS-09 (Fig. 6). Similarly, Raj et al. (2012) also reported that Pseudomonas aeruginosa showed maximum protease production in the presence of yeast extract as a nitrogen source.
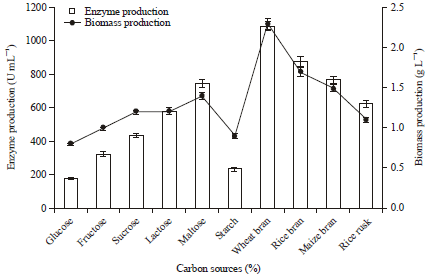 | |
| Fig. 5: | Effect of different carbon sources on protease and biomass production. Test flasks contained different carbon sources in the medium at a level of 2% (w/v). The flasks were inoculated with culture and incubated at 37°C for 48 h at pH 6.5. Error bars presented are mean values of ±Standard deviation of triplicates of three independent experiments |
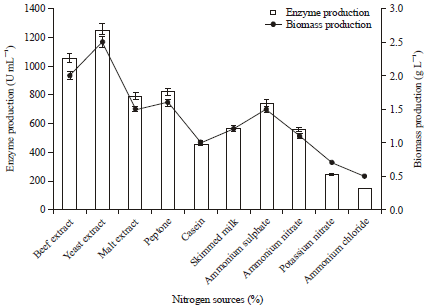 | |
| Fig. 6: | Effect of different nitrogen sources on protease and biomass production. Test flasks contained different nitrogen sources in the medium at a level of 0.5% (w/v). The flasks were inoculated with culture and incubated at 37°C for 48 h at pH 6.5. Error bars presented are mean values of ±Standard deviation of triplicates of three independent experiments |
In contrast to our result, Mukherjee et al. (2008) reported that beef extract followed by yeast extract served as the best co-nitrogen sources for protease production by Bacillus subtilis. Protease production is believed to be affected by the presence of complex nitrogen sources in the growth medium and each organism vary from the other in terms of its requirement for specific nitrogen source.
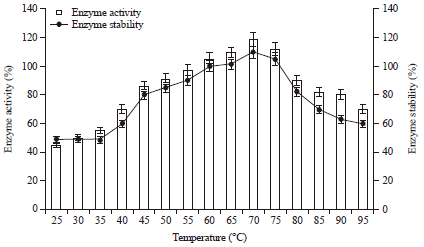 | |
| Fig. 7: | Effect of temperature on enzyme activity and stability. For enzyme activity reaction mixture was incubated at different temperatures (25-95°C) and for stability enzyme was pre-incubated at respective temperatures for 1 h and reaction was conducted as standard assay method |
Characterization of enzyme activity and stability
Effect of temperature on enzyme activity and stability: The protease of Pseudomonas aeruginosa RGSS-09 was completely stable in the broad temperature range of 45-75°C during 1 h incubation. However, with further increase in every 5°C temperature, there was a gradual decrease in enzyme stability ranging between 10-30% up to 95°C. The enzyme retained 90, 82, 80 and 70% activity even after treatment at 80, 85, 90 and 95°C, respectively (Fig. 7). Similarly 100% activity at 70°C for 1 h for protease of Pseudomonas aeruginosa strain K has been reported by Rahman et al. (2006). The protease of strain RGSS-09 is more thermostable than protease studied by several other researchers. These are the properties considered to be very important for industrial protease production. Hence, it is evident that the protease of Pseudomonas aeruginosa RGSS-09 is more thermostable and could be applied for several biotechnological and industrial purposes.
Effect of pH on enzyme activity and stability: The pH stability on the purified amylase was determined by measuring the enzyme activity at varying pH values ranging from 4.0-11.0 using different suitable buffers. It observed that maximum protease activity was established at pH 7.5, however, it was found to be most stable at pH 7.0 (Fig. 8). The relative activities at pH 4.0, 4.5, 5.0 5.5, 6.0, 6.5, 7.0, 7.5 and 8.0 were determined to be 45, 51, 63, 78, 90, 98, 105, 100 and 95%, respectively. At pH above 8.0, the protease activity decreased rapidly. The protease from Pseudomonas aeruginosa RGSS-09 was stable in a range of pH 6.0-10.0 and at pH 11.0 approximately 63% of its activity was retained (Fig. 8). Rahman et al. (2006) also reported that proteases of Pseudomonas aeruginosa strain K are generally stable over a wide range of pH from 6.0-11.0.
Effect of organic solvents on protease stability: In another approach, the effect of various organic solvents (25%, v/v) on protease stability was also investigated for 7 days and the results are depicted in Table 2. The protease of P. aeruginosa RGSS-09 is extraordinarily stable in the presence of all organic solvents under study. It was observed that except benzene, methanol, ethanol and presence of other solvents enhanced the amylase activity.
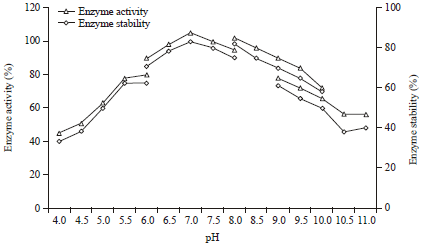 | |
| Fig. 8: | Effect of pH on enzyme activity and stability. For enzyme activity the reaction was assayed at respective pH and for stability enzyme was pre-incubated with buffers (100 mM, in ratio 1:1) of different pH (4.0-11.0) at 37 °C for 1 h and assayed by standard assay method |
| Table 2: | Stability of crude protease activity in presence of various organic solvents |
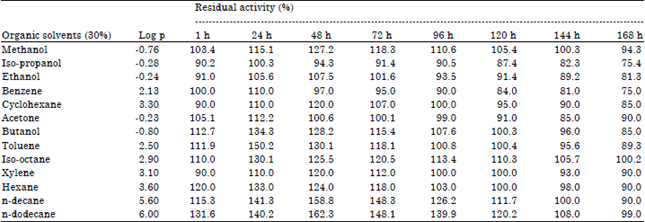 | |
Enzyme was pre-incubated with different organic solvents at a concentration of 25% (v/v) at 37°C for different time period and assayed as standard assay method. The enzyme activity without incubation with organic solvent was taken as 100%. Mean standard deviation for all the values is <±5.0% | |
After incubation with n-dodecane, isooctane, n-decane, xylene, toluene, n-haxane, n-butanol and cyclohexane, the protease activity increased to 162.3, 130.1, 158.8, 120, 150.2, 134.3, 144 and 120%, respectively. The presence of benzene, methanol and ethanol marginally reduced the protease with residual activities of 75, 94.3 and 81.3%, respectively (Table 2). An organic solvent stable alkaline protease has been reported from P. aeruginosa PseA by Gupta and Khare (2007). After 10 days of incubation with organic solvents (25%, v/v), the residual protease activities were 112, 75, 98, 92, 97, 94, 75, 90, 96, 102 and 104% in the presence of ethanol, 1-butanol, benzene, toluene, xylene, cyclohexane, hexane, heptane, isooctane, n-decane and n-dodecane, respectively. Abusham et al. (2009) also reported that protease of B. subtilis strain showed enhanced activity in the presence of organic solvents (25%, v/v). It is therefore; evident from our study that protease of P. aeruginosa RGSS-09 is remarkably stable in the presence of broad range hydrophilic as well as hydrophobic organic solvents employed in this study.
| Table 3: | Effect of different metal ions on protease activity and stability |
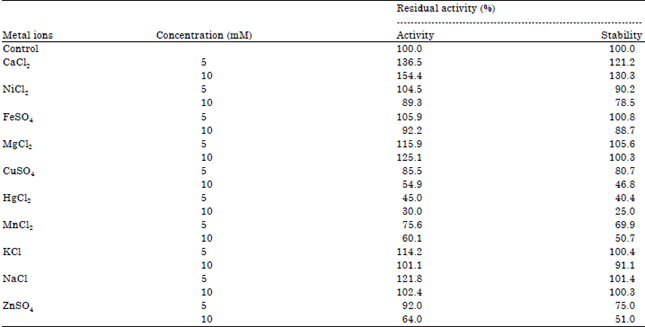 | |
Enzyme activity was determined at 37°C in the presence of metal ions in the reaction mixture directly and for stability enzyme was pre-incubated with different metal ions at 37°C for 1 h and assayed as standard assay method. The enzyme activity without incubation with metal ions was taken as 100%. Mean standard deviation for all the values is <±5.0% | |
Effect of different metal salts on protease activity and stability: For the study of the effect of metal ions viz. NaCl, CaCl2, MgCl2, HgCl2, FeSO4, NiCl2, CuSO4, KCl, MnCl2 and ZnSO4 at a concentration of 5 and 10 mM were individually tested in the basal medium at their optimal temperature, incubation period and pH to observe the effect on enzyme activity and stability by P. aeruginosa RGSS-09. Out of these metal ions, calcium and magnesium ions were showed maximum protease activity (154.4 and 125.1%) and stability (130.3 and 100.3%). While, protease activity and stability was slightly reduced in the presence of K+, Fe2+, Cu2+, Ni2+, Mn2+ and Zn2+. P. aeruginosa RGSS-09 protease activity was strongly inhibited by HgCl2 (Table 3). Nadeem et al. (2007) also reported that protease activity and stability was enhanced in the presence of calcium and magnesium. Similarly, Mn2+, Ca2+ and Mg2+ ions have been described to increase the relative protease activity produced by Bacillus megatarium isolated from thai fish sauce (Yossan et al., 2006). It has been suggested that concerned metal ions apparently protect the enzyme against thermal denaturation and play a vital role in maintaining the active conformation of the enzymes at higher temperatures (Donaghy and Mckay, 1993). Identification of proper ions and their suitable concentrations for rendering thermostability to the enzymes are very important for their applications at commercial levels. The inhibition of protease activity by Hg2+ ions could be due to competition between the exogenous cations and the protein associated cations, resulting in decreased metalloenzymes activity (Sevinc and Demirkan, 2011). Bhatiya and Jadeja (2010) reported that protease activity was strongly inhibited inhibition by Hg2+, Cu2+, Fe2+, Mg2+ and Zn2+.
Effect of surfactants on protease activity and stability: None of the surfactants tested had a pronounced inhibitory effect on enzyme activities. Only Tween-60 showed a moderate inhibitory effect (13.5%) on protease activity of P. aeruginosa RGSS-09 (Table 4). On the other hand, the addition of SDS, Tween-20, Tween-40 and Triton-X-100 (1.0%) was found to stimulate activity by 10, 19.6, 5.7 and 12%, respectively.
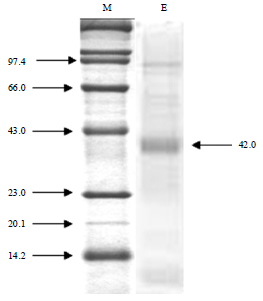 | |
| Fig. 9: | Photographic representation of the SDS-PAGE of protease of P. aeruginosa RGSS-09. Lane 1: Marker proteins, Lane 2: Crude enzyme, Molecular weights were presented in the form of kDa |
| Table 4: | Effect of different surfactants on protease activity |
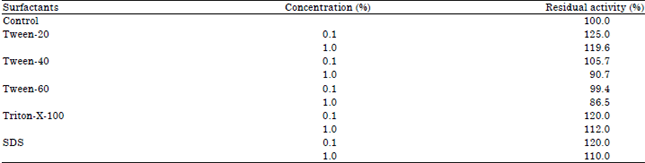 | |
Enzyme was pre-incubated with different surfactants at 37°C for 1 h and assayed as standard assay method. The enzyme activity without incubation with surfactants was taken as 100%. Mean standard deviation for all the values is <±5.0% | |
The stimulatory effect of surfactants on protease activity is controversial. Joo et al. (2004) also reported that the activity of protease of Bacillus sp. was stimulated in the presence of SDS, Tween-20, Tween-40 and Triton-X-100 (1%).
SDS-PAGE analysis of protease enzyme: The purity of the enzyme was confirmed by the presence of a single band on SDS-PAGE and its molecular weight was approximately 42 kDa (Fig. 9) which was similar to Bacillus sp. HUTBS71 (49 kDa) (Akel et al., 2009) but different from Bacillus sp. S17110 (75 kDa) (Jung et al., 2007).
CONCLUSION
A mesophilic strain of P. aeruginosa RGSS-09 was isolated for enhance production of acidic protease from soil sample of diary site. The isolate was able to produce higher protease in broad temperature and pH range. Maximum protease production (1256 U mL-1) was obtained within 48 h in the presence of wheat bran and yeast extract. Our strain was resistance against various metal ions, solvent and surfactant. The potential of this strain to produce higher protease production in the presence of cheap carbon source and short period promises its candidature for the industrial applications.
ACKNOWLEDGMENTS
Financial assistance by Council of science and technology, U.P., is greatly acknowledged by Rajeeva Gaur, Soni Tiwari and Sheffalika Singh.
REFERENCES
- Abusham, R.A., R.N. Rahman, A.B. Salleh and M. Basri, 2009. Optimization of physical factors affecting the production of thermo-stable organic Solvent-tolerant protease from a newly isolated halo tolerant Bacillus subtilis strain Rand. Microb. Cell Fact., Vol. 8.
CrossRef - Adinarayana, K. and P. Ellaiah, 2002. Response surface optimization of the critical medium components for the production of alkaline protease by a newly isolated Bacillus sp. J. Pharmacy Pharmaceut. Sci., 5: 272-278.
PubMedDirect Link - Akel, H., F. Al-Quadan and T.K. Yousef, 2009. Characterization of a purified thermostable protease from hyperthermophilic Bacillus strain HUTBS71. Eur. J. Sci. Res., 31: 280-288.
Direct Link - Balaji, N., K.M. Rajasekaran, N. Kanipandian, V. Vignesh and R. Thirumurugan, 2012. Isolation and screening of proteolytic bacteria from freshwater fish Cyprinus carpio. Int. Multidisciplin. Res. J., 2: 56-59.
Direct Link - Bhattacharya, S., S. Bhardwaj, A. Das and S. Anand, 2011. Utilization of sugarcane bagasse for solid-state fermentation and characterization of α-amylase from Aspergillus flavus isolated from Muthupettai mangrove, Tamil Nadu, India. Aust. J. Basic Applied Sci., 5: 1012-1022.
Direct Link - Donaghy, J.A. and A.M. McKay, 1993. Production and properties of an alkaline protease by Aureobasidium pullulans. J. Applied Bacteriol., 74: 662-666.
CrossRefDirect Link - Gupta, A. and S.K. Khare, 2007. Enhanced production and characterization of a solvent stable protease from solvent tolerant Pseudomonas aeruginosa PseA. Enzyme Microb. Technol., 42: 11-16.
CrossRefDirect Link - Ikram-ul-Haq and H. Mukhtar, 2007. Biosynthesis of acid protease by Penicillium griseoroseum IH-02 in solid-state fermentation. Pak. J. Bot., 39: 2717-2724.
Direct Link - Joo, H.S., C.G. Kumar, G.C. Park, S.R. Paik and C.S. Chang, 2004. Bleach-resistant alkaline protease produced by a Bacillus sp. isolated from the Korean polychaete, Periserrula leucophryna. Process Biochem., 39: 1441-1447.
CrossRef - Josephine, F.S., V.S. Ramya, N. Devi, S.B. Ganapa, K.G. Siddalingeshwara, N. Venugopal and T. Vishwanatha, 2012. Isolation, production and characterization of protease from Bacillus sp isolated from soil sample. J. Microbiol. Biotechnol. Res., 2: 163-168.
Direct Link - Jung, S.C., H.R. Paik, M.S. Kim, K.S. Baik, W.Y. Lee, C.N. Seong and S.K. Choi, 2007. InhA-like protease secreted by Bacillus sp. S17110 inhabited in Turban shell. J. Microbiol., 45: 402-408.
PubMed - Kalaiarasi, K. and P.U. Sunitha, 2009. Optimization of alkaline protease production from Pseudomonas fluorescens isolated from meat waste contaminated soil. Afr. J. Biotechnol., 8: 7035-7041.
Direct Link - Kumar, D.J.M., P. Venkatachalam, N. Govindarajan, M.D. Balakumaran and P.T. Kalaichelvan, 2012. Production and purification of alkaline protease from Bacillus sp. MPTK 712 isolated from dairy sludge. Global Veterinaria, 8: 433-439.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Maase, F.W.J.L. and R. van Tilburg, 1983. The benefit of detergent enzymes under changing washing conditions. J. Am. Oil Chem. Soc., 60: 1672-1675.
CrossRefDirect Link - Mukherjee, A.K., H. Adhikari and S.K. Rai, 2008. Production of alkaline protease by a thermophilic Bacillus subtilis under Solid-State Fermentation (SSF) condition using Imperata cylindrica grass and potato peel as low-cost medium: Characterization and application of enzyme in detergent formulation. Biochem. Eng. J., 39: 353-361.
CrossRefDirect Link - Qadar, S.A.U., E. Shireen, S. Iqbal and A. Anwar, 2009. Optimization of protease production from newly isolated strain of Bacillus sp. PCSIR EA-3. Indian J. Biotechnol., 8: 286-290.
Direct Link - Radha, S., V.J. Nithya, R.H. Babu, A. Sridevi, N.B.L. Prasad and G. Narasimha, 2011. Production and optimization of acid protease by Aspergillus spp under submerged fermentation. Arch. Applied Sci. Res., 3: 155-163.
Direct Link - Rahman, R.N.Z.R.A., L.P. Geok, M. Basri and A.B. Salleh, 2006. An organic solvent-stable alkaline protease from Pseudomonas aeruginosa strain K: Enzyme purification and characterization. Enzyme Biotechnol. Biomol. Sci., 39: 1484-1491.
CrossRef - Raj, A., N. Khess, N. Pujari, S. Bhattacharya, A. Das and S.S. Rajan, 2012. Enhancement of protease production by Pseudomonas aeruginosa isolated from dairy effluent sludge and determination of its fibrinolytic potential. Asian Pac. J. Trop. Biomed., 2: S1845-S1851.
CrossRefDirect Link - Rao, M.B., A.M. Tanksale, M.S. Ghatge and V.V. Deshpande, 1998. Molecular and biotechnological aspects of microbial proteases. Microbiol. Mol. Biol. Rev., 62: 597-635.
Direct Link - Sevinc, N. and E. Demirkan, 2011. Production of protease by Bacillus sp. N-40 isolated from soil and its enzymatic properties. J. Biol. Environ. Sci., 5: 95-103.
Direct Link - Yossan, S., A. Reungsang and M. Yasuda, 2006. Purification and characterization of alkaline protease from Bacillus megaterium isolated from Thai fish sauce fermentation process. Sci. Asia, 32: 377-383.
CrossRef - Smita, G.S., P. Ray and S. Mohapatra, 2012. Quantification and optimisation of bacterial isolates for production of alkaline protease. Asian J. Exp. Biol. Sci., 3: 180-186.
Direct Link