
J.J. Oloche
Department of Pharmacology, College of Health Sciences, Benue State University, Makurdi, Nigeria
G.O. Obochi
Department of Biochemistry, College of Health Sciences, Benue State University, Makurdi, Nigeria
Asian Journal of Biochemistry
Year: 2015 | Volume: 10 | Issue: 5 | Page No.: 180-189







ABSTRACT
The sodium pump or Na+/K+ ATPase is a membrane-bound enzyme that maintains the Na+ and K+ gradients across the plasma membrane of animal cells. Many basic and specialized cellular functions depend on the activity of this enzyme, hence, its adaptability to changing cellular and physiological stimuli. This review presents an overview of the many adaptations or mechanisms that regulate sodium pump activity in a tissue-specific manner. These mechanisms include regulations by substrates, membrane associated components such as cytoskeletal elements and the γ-subunits and circulating endogenous inhibitors as well as a variety of hormones, including corticosteroids, peptide-hormones and catecholamine. In addition, this review considers the effects of a range of specific intracellular signaling pathways involved in the regulation of sodium pump activity.
PDF Abstract XML References Citation
Received: July 23, 2015;
Accepted: August 28, 2015;
Published: September 21, 2015
How to cite this article
J.J. Oloche and G.O. Obochi, 2015. Sodium Pump Adaptability to Tissue-Specific Regulation: A Review. Asian Journal of Biochemistry, 10: 180-189.
DOI: 10.3923/ajb.2015.180.189
URL: https://scialert.net/abstract/?doi=ajb.2015.180.189
DOI: 10.3923/ajb.2015.180.189
URL: https://scialert.net/abstract/?doi=ajb.2015.180.189
INTRODUCTION
The Na+/K+ adenosine triphosphatase (ATPase) is an enzyme and a glycoprotein composed of 2 α and 2 β chains. Its activity depends on the presence of Na+ and K+ and requires adenosine triphosphate (ATP) and Mg2+ as cofactors. The enzyme hydrolyzes a high energy phosphate bond of ATP and uses the energy thus released to transport 3 Na+ outside and simultaneously 2 K+ inside across the cell membrane (Therien and Blostein, 2000).
Sodium pump is also called Na+/K+ ATPase, located at basal and lateral plasma membrane of intestinal and renal cells. It actively transports sodium ion (Na+) required for its activity. The Na+/K+ ATPase maintain both magnitude and direction of transmembrane concentration gradients of Na+ and K+ (Aperia et al., 1992; Anderson et al., 1998). The Na+/K+ ATPase is very active in those cells where activities depends largely on transmembrane Na+ fluxes such as in the nervous system, muscle fibres, renal tubule cells and intestinal mucosal cells (Clausen, 1996; Aperia et al., 1996). Na+/K+ ATPase exist in two isoforms; E1 and E2. The E1 presents its ion binding and phosphate binding sites on the cytoplasmic surface of the membrane to which three Na+ from the cytoplasm bind leading to phosphorylation of aspartate residue of E1 (Fig. 1). The process is ATP and Mg2+ dependent. This results in conformational change of E1→E2. The E2 exposes both ion binding and phosphate binding sites on the extracellular surface of the membrane, thus lowering the affinity of ATPase for Na+ and releases it into the extracellular fluid in exchange for K+ extracellular fluid (ECF) causing dephosphorylation. Dephosphorylation again changes the conformation of E2→E1 thereby lowering its affinity for K+ from the ATPase into the cell (Therien and Blostein, 2000).
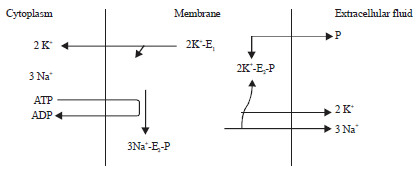 | |
| Fig. 1: | Mechanism of action of sodium pump (source: Anderson et al., 1998) |
Thus, the Na+ transported actively by sodium pump diffuses into the cell across its luminal or microvillus membrane from the lumen. Active absorption of Na+ is coupled with glucose absorption or amino acid absorption (Aperia et al., 1992).
Ouabain, a cardiotonic steroidal glycoside obtained from digitalis inhibits the Na+/K+ ATPase by blocking dephosphorylation of E2 conformer (Hamlyn et al., 1996), whereas, vanadate inhibits the pump in a similar manner when present inside the cell (Anderson et al., 1998).
The basic function of Na+/K+ ATPase or sodium pump is to maintain Na+ and K+ gradient across the plasma membrane of animal cells. The sodium pump is important in regulating action potential, cell volume, cytoplasmic pH and Ca2+ levels through the Na+/H+ and Na+/Ca2+ exchangers, respectively and in driving a variety of secondary transport processes such as Na+-dependent glucose and amino acid transport.
In nerve transmission, transient propagated reversal of charge on the axonal membrane referred to as nerve action potential arriving pre-synaptic terminal cause the release of excitatory or inhibitory neurotransmitter (Rang et al., 2011; Stephanie, 2015). A combination of excitatory transmitter with pre-synaptic receptors produces a localized depolarization resulting in excitatory postsynaptic potential through an increase in permeability of Na+. On the other hand, inhibitory transmitter causes selective permeability to K+ or Cl‾ resulting in hyperpolarisation, hence, inhibitory postsynaptic potential (Rang et al., 2011; Harvey et al., 2012).
In skeletal muscles, released transmitter (Acetylcholine) at motor end-plate binds to nicotinic acetylcholine receptors causing increase in Na+ and K+ conductance in end-plate membrane with consequent generation of action potential in muscle fibres (Beebe, 1994; Rang et al., 2011; Stephanie, 2015). Action potential in muscle fibres lead to inward spread of depolarization along T-tubules, which in turn open L-type Ca2+ channels. Mechanical coupling of L-type Ca2+ channel with Ryanodine receptor activates the release of Ca2+ from sarcoplasmic reticulum and diffusion from thick and thin filaments. The Ca2+ activates troponin C resulting in the formation of cross-linkages between actin and myosin and sliding of thin and thick filaments produce contraction (Rang et al., 2011; Stephanie, 2015).
On the contrary, skeletal muscle relaxation occurs most importantly as a result of sequestration of Ca2+ in sarcoplasmic reticulum. ATP-dependent Ca2+ pump, pumps Ca2+ against concentration gradient into sarcoplasmic reticulum which then bind to calsequestrin and calreticulin (Rang et al., 2011). Another mechanism is by extrusion of Ca2+ across plasma membrane via Na+-Ca2+ exchanger and plasma membrane Ca2+ pump (Doucet, 1988; Harvey et al., 2012). These processes result in detachment of cross-bridges from the thin filament, restoration of troponin-tropomyosin regulated inhibition of actin and myosin interaction, followed by skeletal muscle relaxation.
In smooth muscles, contraction occurs when hormone, neurotransmitter or depolarization activates the release of Ca2+ from sarcoplasmic reticulum, ligand gated Ca2+ channels or voltaged gated Ca2+ channels. Released Ca2+ bind to calmodulin causing increase in cyclic adenosine triphosphate, which phosphorylates myosin light chain to form myosin ATPase a process catalysed by myosin light chain kinase. Phosphorylated myosin bind to actin forming cross-bridge producing contraction (Wang et al., 1995; Meister and Aperia, 1993; Rang et al., 2011).
The sodium pump is the target of multiple regulatory mechanisms activated in response to changing cellular requirements. The requirement for modulators of the Na+/K+ ATPase is likely to be greatest in tissues in which perturbations of the intracellular alkali cation content underlie their specialized functions, such as; nerve impulse propagation, exercise and changes in diet (Skou, 1957; Beebe, 1994).
One primary need for sodium pump adaptation comes from changes in dietary Na+ and K+. The mediators of natriuresis and diuresis; hormones that control the volume and ionic composition of blood and urine, often act directly on the sodium pump of the nephron and intestine. The function of the pump in absorption or reabsorption of Na+ and K+ and secondarily, other solutes requires tight regulation of the enzyme to maintain normal levels of Na+ and K+ during altered salt intake (Blanco and Mercer, 1998). The Na+ transport across epithelia is linked with water reabsorption. Thus, impairment of the sodium pump in kidney and small intestine is associated with the pathophysiology of hypertension and chronic diarrhea, respectively (Bonvalet, 1998; Blaustein, 1996).
In excitable tissues such as neurons, skeletal muscle cells and pace maker fibres of the heart, the sodium pump must reestablish the electrical potential across the plasma membrane following excitation-induced depolarization (Clausen, 1996; Hernandez, 1992). In skeletal muscle, continuous stimulation of muscle fibre during exercise leads to dissipation of the cation gradient necessary for muscle contraction. To offset excessive release of K+ from the muscle cells during exercise, rapid activation of Na+/K+ ATPase is essential to cause delay in the onset of muscular fatigue and reducing potentially toxic levels of plasma K+ (Anderson et al., 1998). The Na+/K+ ATPase regulation in cardiac muscle is particularly critical to the myocardium, where the enzyme indirectly regulate contraction (Clausen, 1996; Blaustein, 1996). Thus, the sodium pump controls the steady-state cytoplasmic Na+ concentration which then determines Ca2+ concentration via the Na+/Ca2+ exchange. The Ca2+, in turn, is pumped into the Sarcoplasmic Reticulum (SR) by the Sarco-Endoplasmic Reticulum Calcium (SERCA) pumps (Doucet, 1988; Clausen, 1996).
Regulation of the sodium pump in these tissues is paramount for determination of the "set point" for cardiac muscle contraction and the steady-state contraction of vascular smooth muscle (Nielsen and Clausen, 1996). Physiological regulation that act in a manner analogous to that ascribed to cardiac glycoside inhibition of the Na+/K+ ATPase are likely to be critical for normal heart contraction. Thus, the force of contraction is increased via increasing cell Na+ and may be considered to be the basis of digitalis therapy for cardiac insufficiency (Hernandez, 1992; Gloor, 1997).
This study therefore, focuses on adaptations to tissue-specific regulation of the sodium pump with emphasis on two mechanisms involving pathways that result in modulation in sodium pump activity and the interaction of the pump complex with other membrane components, causing further modulation via other signaling cascades.
Role of substrate concentration in sodium pump activity: The simplest and most straight forward determinants of sodium pump activity are the concentrations of substrates. The sodium pump is activated by Na+ at extracellular sites. Thus, small changes in the cytoplasmic Na+ concentration secondary to activation of either various Na+ dependent transporters or Na+ channels can have dramatic effect on sodium pump activity (Blaustein, 1996; Skou, 1957).
The high affinity of the enzyme for K+ at activating sites generally precludes the effects of variations in extracellular K+ concentrations which have been shown to act as a competitive inhibitor of Na+ binding at cytoplasmic sites (Garay and Garrahan, 1973). The variation in cytoplasmic K+ concentration, or, more likely, in the affinity of the enzyme for K+ as an antagonist at cytoplasmic Na+ activating binding sites, is a plausible mechanism for determining the set point for the physiological concentration at half maximal activation (k 0.05) for cytoplasmic Na+ activation. Hence, the use of K+ channels activators as antiarrhythmic agents. Half-maximal activation of the enzyme by intracellular Na+ occurs at concentrations of -10 to 40 mm, which, depending on the tissue, are often at or above the steady state Na+ concentration (Blaustein, 1996). Because the physiological concentration at half maximal activation of the Na+/K+ ATPase for ATP is between 300 and 800 mm (Blaustein, 1996), the ATP concentration in most cells is saturating for the enzyme. However, in some tissues and under certain conditions, ATP levels may fall to sub-saturating levels. For example, cells of the kidney medulla are known to function under near anoxic conditions (Brezis and Rosen, 1995) and such conditions could lead to dramatic drop in ATP levels (Soltoff and Mandel, 1984). Thus, variations in ATP concentration or in the affinity of the sodium pump for ATP may be a physiologically relevant mechanism of pump regulation in this tissue.
Role of membrane-associated components in sodium pump activity: The Na+/K+ ATPase is a membrane-embedded protein, hence, the nature of the membrane components might be important determinants of enzyme function. Little is known on the nature and mechanistic basis of sodium pump modulation by specific membrane components (the role of membrane lipids). The main effects of lipids on the sodium pump are related to membrane fluidity and thickness (Paller, 1994). Membrane lipids promote bilayer formation of physiological thickness and increased fluidity which tend to promote optimal Na+/K+ ATPase activity as do negatively charged lipids such as phosphatidylserine and phosphatidylglycerol (Paller, 1994; Kimelberg and Papahadjopoulos, 1972). The effects of cholesterol on enzyme activity are often related to membrane fluidity free fatty acids present in the membrane or as the products of phospholipase A2 (PL A2) dependent regulatory pathway tend to inhibit the Na+/K+ ATPase (Oguchi et al., 1993).
Interactions of sodium pump with components of the cytoskeleton of cells: Components of the cytoskeleton are proteins include spectrin, actin, adducin, pasin and ankyrin (Cantiello, 1995, 1997). The main consequence of interactions between the Na+/K+ ATPase and the cytoskeleton is proper processing and targeting of sodium pump to the appropriate membrane-compartment. Example, disruptions in the cellular distribution of Na+/K+ ATPase induced either by ATP depletion or hypoxia, are linked to alterations in cytoskeleton proteins and a spectrin-ankyrin complex is required for transport of pumps from the endoplasmic reticulum to the Golgi apparatus (Paller, 1994). Recently, actin has been shown to activate the sodium pump by a mechanism mediated by cAMP-dependent protein kinase (Skou, 1957).
Thus, the role of cytoskeleton proteins in regulating sodium pump is suggestive of a mechanism, which appears to involve acceleration of the rate of the conformational change E2 (K+)→E1 (Na+) or E2 (K+) ATP→E1 (Na+) ATP.
Role of gamma (γ)-subunit in sodium pump activity: The γ-subunit is a small transmembrane protein that specifically associate with the Na+/K+ ATPase in a tissue-specific manner and thus mediate increase in the affinity of the enzyme for ATP (Arystarkhova et al., 1999), which may however, lead to a secondary decrease in its apparent affinity for K+ (Ewart and Klip, 1995). The γ-subunit is expressed in the presence of Na+/K+ ATPase (Arystarkhova et al., 1999; Ewart and Klip, 1995). The likely change in ATP affinity for K+ that is perhaps of physiological importance. In most cells, ATP levels are sufficient to saturate the Na+/K+ ATPase and therefore, a modest shift in ATP affinity should not have dramatic effects (Ewart and Klip, 1995).
Role of circulating endogenous inhibitors in sodium pump activity: Circulating endogenous cardiac glycosides such as ouabain inhibit Na+/K+ ATPase (Hamlyn et al., 1996) and is synthesized in the adrenal gland. Structurally, endogenous cardiac glycosides are homologous to ouabain, consisting of a cholesterol core conjugated to either a lactone or pyrone ring and containing various combinations of hydroxylsulfate or carbohydrate groups (Blaustein, 1977; Rossi et al., 1995). The main physiological role of endogenous cardiac glycosides appears to be in regulating blood pressure. Thus, hypertension has been linked to increased levels of plasma endogenous cardiac glycosides and can result from long term treatment with cardiac glycosides (Rossi et al., 1995). The mechanism by which endogenous cardiac glycosides mediate increased blood pressure is linked to the transmembrane equilibrium between Na+ and Ca2+ via the Na+/Ca2+ exchanger (Blaustein, 1977). Thus, inhibition of the sodium pump in vascular smooth muscle cells and myocytes by endogenous cardiac glycosides leads to an increase in the cytoplasmic Na+ concentration, causing Ca2+ to enter the cell and be sequestered in the sarcoplasmic reticulum. Increased Ca2+ in the sarcoplasmic reticulum causes greater and sustained contraction of the vascular and heart muscle fibers, directly increasing blood pressure. Such a mechanism is believed to be the basis for the partial reversal of cardiac insufficiency following treatment with cardiac glycosides (Thomas et al., 1990). However, renal dysfunction appears to be more implicated in hypertension rather than changes in peripheral tension caused by endogenous cardiac glycosides.
Role of peptide hormone and catecholamine on activity sodium pump: The Na+/K+ ATPase is subjected to both short and long term regulation by a variety of hormones. Some hormones appear to alter sodium pump activity by changing its apparent affinity for Na+ (Wang et al., 1995). Sodium ion has been shown to induce other mechanisms of up-regulation of the sodium pump (Whorwood and Stewart, 1995). The Na+ influx is thought to be the first signal leading to an increase in membrane Na+ in one kind of aldosterone-mediated short-term regulation (Geering et al., 1982).
Short term regulation involves either direct effect on the kinetic behavior of the enzyme or translocation of sodium pump between the plasma membrane and intracellular stores. On the other hand, long term regulatory mechanisms generally affect de novo Na+/K+ ATPase synthesis or degradation.
Catecholamine such as epinephrine, norepinephrine and dopamine affect Na+/K+ ATPase activity in the kidney and as such regulate salt and fluid reabsorption (Meister and Aperia, 1993). Dopamine is a natriuretic factor synthesized in the kidney proximal tubule. Dopamine inhibits Na+/K+ ATPase in the kidney and this represents a physiologically important mechanism for regulating salt reabsorption during high salt intake (Nathanson et al., 1995). Dopamine-dependent inhibition of Na+/K+ ATPase appears to be both age related and cell specific (Bertorello and Aperia, 1990).
Epinephrine is involved in stimulating K+ uptake by skeletal muscle after exercise induced hyperkalemia, whereas norepinephrine, acting as dopamine antagonist, appears to have a role in Na+ reabsorption in the nephron (Meister and Aperia, 1993). Several catecholamine, including norepinephrine, act as neurotransmitters in the central nervous system. Their likely importance in the activation of Na+/K+ ATPase in neural tissue is to reestablish the electrochemical cation gradient across the cell membrane following transmission of electrical impulses (Hernandez, 1992).Peptide hormones such as insulin regulate Na+/K+ ATPase activity. Insulin, a metabolic hormone that regulates glycolytic storage plays an important role in K+ homeostasis. A well known effect of insulin is the stimulation of the Na+/K+ ATPase which result in increased uptake of K+ by various tissues (Vaandrager and de Jonge, 1996). Insulin therefore, mediates cell function by binding to the insulin receptor, which leads to activation of a variety of intracellular signaling processes. In some diabetic conditions, insulin decreases Na+/K+ ATPase activity (Vaandrager and de Jonge, 1996).
Effects of protein phosphorylation cascade in long term regulation of Na+/K+
ATPase by corticosteroids: Steroid hormones have specific long and short term regulatory effects on the Na+/K+ ATPase (Bertorello and Aperia, 1989). Long-term effects are generally, mediated by changes in mRNA/protein synthesis induced by direct interaction of receptor/corticosteroid complexes with nuclear DNA example, mineralocorticoid [aldosterone] and the glucocorticoid [dexamethasone] (Verrey et al., 1996).
Endogenous corticosteroid such as aldosterone is synthesized in and released by the adrenal cortex. Aldosterone plays an important role in Na+ and K+ transport in epithelial tissues of the kidney. Its physiological role involves adaptation to decrease in Na+ or increase in K+ intake (O'Neil, 1990), which is a target site for aldosterone antagonist in the management of hypertension. It has been shown that the main effect of aldosterone and dexamethasone on the Na+/K+ ATPase is to sustain a long term increase in expression of sodium pump observed directly or as an increase in ouabain binding in kidney and kidney derived cell lines, colon, heart, inner ear and liver cells, vascular smooth muscle cells and cardiocytes (Whorwood and Stewart, 1995). While, aldosterone increase sodium pump mRNA expression of the α-and β-subunit genes via mineralocorticoid (type 1) receptors, dexamethasone presumably bind to glucocorticoid (type II) receptors (Wang et al., 1995).
Corticosteroid/receptor complexes mediate mRNA synthesis by interacting with regulatory elements of both the α1 and β1-subunits genes (Oguchi et al., 1993). Corticosteroid-mediated increase in protein synthesis of sodium pump may be dependent on changes in cytoplasmic Na+ concentration by abrogation of the effects in the presence of blockers of Na+ transport. In mammals the effects of corticosteroid may be facilitated by the thyroid hormone triiodothyronine (Geering et al., 1982). Interestingly, long-term stimulation of the sodium pump by aldosterone is abrogated by inhibitors of the protein phosphatase calcineurin (Aperia et al., 1996). There is evidence that cAMP-inducible factors have a role in mediating aldosterone-dependent increase in both α and β subunit mRNA (Whorwood et al., 1994). These findings may suggest the involvement of a protein phosphorylation cascade in long term regulation by corticosteroids.
Effect of cyclic AMP activated protein kinases on the Na+/K+ pump: Cyclic AMP (cAMP) activated protein kinase A is activated by the intracellular accumulation of cAMP. The cAMP levels in the cell is regulated by adenylate cyclase which synthesize it and cAMP phosphodiesterase, which degrades it. Therefore, signals that activate or inhibit these two enzymes affect cAMP levels and thus protein kinase A activation, which phosphorylates the sodium pump (Beebe, 1994).
The cascade of events that result in activation of protein kinase C is usually initiated by activation of the membrane bound phospholipase C, which cleaves phospholipids into two components; phosphatidylinositol triphosphate (which, in turn, increases cytosolic Ca2+) and diacylglycerol [DAG] (Lea et al., 1994). The DAG allows the inactive, cytoplasmic form of protein kinase C to bind to the membrane and increases its affinity for Ca2+ and phospholipids, its final activators. Activated protein kinase C is a potent regulator of many enzymes, including the Na+/K+ ATPase (Lea et al., 1994), which involves phosphorylation of the Na+/K+ ATPase activity. The phosphorylation of the sodium pump is facilitated by agents that stabilize the E2 conformation of the enzyme and that the Na+/K+ ATPase itself can stimulate autophosphorylation (Borin, 1997). Cyclic guanylate monophosphate (GMP)-dependent protein kinase G is a kinase with specific effect on the Na+/K+ ATPase. Protein kinase G is activated by the cytoplasmic concentration of cGMP which is regulated by guanylate cyclase and degraded by cGMP phosphodiesterase (Vaandrager and de Jonge, 1996). Increase in cGMP has been shown to inhibit the Na+/K+ ATPase in colon, skeletal muscle, brain and alveolar cells (Clausen, 1996; Nielsen and Clausen, 1996). The mechanism of protein kinase G activation appears to involve activation of guanylate cyclase by Nitric Oxide (NO). The NO stimulates cGMP synthesis, which is mediated by the neurotransmitter acetylcholine and glutamate (McKee et al., 1994; Nathanson et al., 1995). The cGMP appears to stimulate the phosphorylation of the sodium pump indirectly by increasing Na+ influx via the Na+/K+ ATPase co-transporter (O’Neil, 1990). Thus nitrates are used therapeutically to relieve angina pain as they release NO, which is a potent vasodilator and enhance myocardial blood flow.
Effect of protein phosphatases on Na+/K+ pump: Many of the effects of protein kinase on the Na+/K+ ATPase can be reversed by protein phosphatases. Majorly, protein phosphatase inhibitors and protein phsphatase-2B (PP-2B) are involved in protein phosphatase dependent modulation of the Na+/K+ ATPase. Protein phosphatase inhibitors counteract the effects of protein kinases by pump inhibition through dopamine bound dopamine-receptor and isoproterenol via the β-adrenergic receptor. Such inhibition is mediated in part by the activation of the protein phosphatase inhibitors dopamine receptor protein phosphatase-32 (DAR PP-32) and inhibitor-I (Aperia et al., 1992). Thus, it has been shown that increase in cAMP levels mediated by dopamine or isoproterenol in kidney and brain leads to phosphorylation of DAR PP-32, which, in turn, cause inhibition of protein phosphatase inhibitors (Cantiello, 1997). The physiological role of PP-2B, which is a Ca2+ and calmodulin-dependent enzyme upon activation by epinephrine and α-adrenergic receptor agonists, activates the Na+/K+ ATPase of most segments of the nephron (Lea et al., 1994), although, its main effects are on the enzyme of proximal convoluted tubules (Aperia et al., 1996).
The role of calcineurin in the kidney is to counter dopamine-induce inhibition of Na+/K+ ATPase and occurs by dephosphorylating targets of dopamine-stimulated protein kinases (Aperia et al., 1992; Lea et al., 1994). Thus, it may suggest that calcineurin mediates its stimulatory effects at least in part by increasing the apparent affinity of the sodium pump for Na+ (Lea et al., 1994). In addition to its role in the kidney, calcineurin mediates ouabain-pumps and has a role, in sodium pump activation during glutamate toxicity in neurons (Aperia et al., 1996).
Protein phosphatase 2A has been shown to modulate Na+/K+ ATPase activity by increasing sodium pump plasma membrane expression in cortical collecting duct (Blot-Chabaud et al., 1996) and antagonise protein kinase C-mediated inhibition of the Na+/K+ ATPase (Blanco and Mercer, 1998). Paradoxically, inhibitors of PP 2A stimulate the pump in hepatocytes (Meister and Aperia, 1993).
CONCLUSION
The need for the ubiquitous Na+/K+ ATPase to adapt to the diverse needs of different tissues underscores the importance of mechanisms for regulating its activity. The signaling cascades involved in hormonal regulation, in particular, are varied and complex. Alterations in activity may be the result of post translational modification, such as, phosphorylation. In certain instances, alterations in Na+/K+ ATPase activity and kinetic behavior result from specific interaction with other membrane components. These include proteins intrinsic to the plasma membrane as well as those of the cytoskeleton.
REFERENCES
- Therien, A.G. and R. Blostein, 2000. Mechanisms of sodium pump regulation. Am. J. Physiol.-Cell Physiol., 279: C541-C566.
PubMedDirect Link - Anderson, R.M., S.X.J. Cheng and A. Aperia, 1998. Forskolin-induced down-regulation of Na+,K+-ATPase activity is not associated with internalization of the enzyme. Acta Physiologica Scandinavica, 164: 39-46.
CrossRefDirect Link - Aperia, A., J. Fryckstedt, U. Holtback, R. Belusa and X.J. Cheng et al., 1996. Cellular mechanisms for bi-directional regulation of tubular sodium reabsorption. Kidney Int., 49: 1743-1747.
CrossRefDirect Link - Aperia, A., F. Ibarra, L.B. Svensson, C. Klee and P. Greengard, 1992. Calcineurin mediates α-adrenergic stimulation of Na+,K+-ATPase activity in renal tubule cells. Proc. Natl. Acad. Sci. USA., 89: 7394-7397.
CrossRefPubMedDirect Link - Arystarkhova, E., R.K. Wetzel, N.K. Asinovski and K.J. Sweadner, 1999. The γ subunit modulates Na+ and K+ affinity of the renal Na,K-ATPase. J. Biol. Chem., 274: 33183-33185.
CrossRefDirect Link - Beebe, S.J., 1994. The cAMP-dependent protein kinases and cAMP signal transduction. Seminars Cancer Biol., 5: 285-294.
PubMedDirect Link - Blanco, G. and R.W. Mercer, 1998. Isozymes of the Na-K-ATPase: Heterogeneity in structure, diversity in function. Am. J. Physiol.-Renal Physiol., 275: F633-F650.
PubMedDirect Link - Blaustein, M.P., 1996. Endogenous ouabain: Role in the pathogenesis of hypertension. Kidney Int., 49: 1748-1753.
CrossRefDirect Link - Blaustein, M.P., 1977. Sodium ions, calcium ions, blood pressure regulation and hypertension: A reassessment and a hypothesis. Am. J. Physiol.-Cell Physiol., 232: C165-C173.
PubMedDirect Link - Blot-Chabaud, M., N. Coutry, M. Laplace, J.P. Bonvalet and N. Farman, 1996. Role of protein phosphatase in the regulation of Na+-K+-ATPase by vasopressin in the cortical collecting duct. J. Membr. Biol., 153: 233-239.
CrossRefDirect Link - Bonvalet, J.P., 1998. Regulation of sodium transport by steroid hormones. Kidney Int., 65: S49-S56.
PubMedDirect Link - Borin, M.L., 1997. Roles of PKA and PKC in regulation of Na+ pump activity in vascular smooth muscle cells. Ann. N. Y. Acad. Sci., 834: 576-578.
CrossRefPubMedDirect Link - Brezis, M. and S. Rosen, 1995. Hypoxia of the renal medulla-its implications for disease. New Engl. J. Med., 332: 647-655.
CrossRefDirect Link - Cantiello, H.F., 1995. Actin filaments stimulate the Na+-K+-ATPase. Am. J. Physiol.-Renal Physiol., 269: F637-F643.
PubMedDirect Link - Cantiello, H.F., 1997. Changes in actin filament organization regulate Na+, K+-ATPase activity: Role of actin phosphorylation. Ann. N. Y. Acad. Sci., 834: 559-561.
CrossRefPubMedDirect Link - Clausen, T., 1996. The Na+, K+ pump in skeletal muscle: Quantification, regulation and functional significance. Acta Physiologica Scandinavica, 156: 227-235.
CrossRefDirect Link - Nielsen, O.B. and T. Clausen, 1996. The significance of active Na+,K+ transport in the maintenance of contractility in rat skeletal muscle. Acta Physiologica Scandinavica, 157: 199-209.
CrossRefPubMedDirect Link - Doucet, A., 1988. Function and control of Na-K-ATPase in single nephron segments of the mammalian kidney. Kidney Int., 34: 749-760.
CrossRefDirect Link - Ewart, H.S. and A. Klip, 1995. Hormonal regulation of the Na+-K+-ATPase: Mechanisms underlying rapid and sustained changes in pump activity Am. J. Physiol.-Cell Physiol., 269: C295-C311.
Direct Link - Garay, R.P. and P.J. Garrahan, 1973. The interaction of sodium and potassium with the sodium pump in red cells. J. Physiol., 231: 297-325.
CrossRefDirect Link - Geering, K., M. Girardet, C. Bron, J.P. Kraehenbuhl and B.C. Rossier, 1982. Hormonal regulation of (Na+,K+)-ATPase biosynthesis in the toad bladder. Effect of aldosterone and 3,5,3'-triiodo-L-thyronine. J. Biol. Chem., 257: 10338-10343.
PubMedDirect Link - Gloor, S.M., 1997. Relevance of Na,K-ATPase to local extracellular potassium homeostasis and modulation of synaptic transmission. FEBS Lett., 412: 1-4.
CrossRefDirect Link - Hamlyn, J.M., B.P. Hamilton and P. Manunta, 1996. Endogenous ouabain, sodium balance and blood pressure: A review and a hypothesis. J. Hypertens., 14: 151-167.
PubMed - Hernandez, R.J., 1992. Na+/K+-ATPase regulation by neurotransmitters. Neurochem. Int., 20: 1-10.
CrossRefDirect Link - Kimelberg, H.K. and D. Papahadjopoulos, 1972. Phospholipid requirements for (Na+ + K+)-ATPase activity: Head-group specificity and fatty acid fluidity. Biochimica Biophysica Acta (BBA)-Biomembr., 282: 277-292.
CrossRefPubMedDirect Link - Lea, J.P., J.M. Sands, S.J. McMahon and J.A. Tumlin, 1994. Evidence that the inhibition of Na+/K+-ATPase activity by FK506 involves calcineurin. Kidney Int., 46: 647-652.
CrossRefPubMedDirect Link - McKee, M., C. Scavone and J.A. Nathanson, 1994. Nitric oxide, cGMP and hormone regulation of active sodium transport. Proc. Natl. Acad. Sci. USA., 91: 12056-12060.
PubMedDirect Link - Meister, B. and A. Aperia, 1993. Molecular mechanisms involved in catecholamine regulation of sodium transport. Semin. Nephrol., 13: 41-49.
PubMedDirect Link - Nathanson, J.A., C. Scavone, C. Scanlon and M. McKee, 1995. The cellular Na+ pump as a site of action for carbon monoxide and glutamate: A mechanism for long-term modulation of cellular activity. Neuron, 14: 781-794.
CrossRefPubMedDirect Link - Oguchi, A., U. Ikeda, T. Kanbe, Y. Tsuruya and K. Yamamoto et al., 1993. Regulation of Na-K-ATPase gene expression by aldosterone in vascular smooth muscle cells. Am. J. Physiol.-Heart Circ. Physiol., 265: H1167-H1172.
PubMedDirect Link - O'Neil, R.G., 1990. Aldosterone regulation of sodium and potassium transport in the cortical collecting duct. Semin. Nephrol., 10: 365-374.
PubMed - Paller, M.S., 1994. Lateral mobility of Na,K-ATPase and membrane lipids in renal cells. Importance of cytoskeletal integrity. J. Membr. Biol., 142: 127-135.
CrossRefDirect Link - Rossi, G., P. Manunta, J.M. Hamlyn, E. Pavan, R. de Toni, A. Semplicini and A.C. Pessina, 1995. Immunoreactive endogenous ouabain in primary aldosteronism and essential hypertension: Relationship with plasma renin, aldosterone and blood pressure levels. J. Hypertens., 13: 1181-1191.
PubMed - Skou, J.C., 1957. The influence of some cations on an adenosine triphosphatase from peripheral nerves. Biochem. Biophys. Acta, 23: 394-401.
CrossRefPubMedDirect Link - Soltoff, S.P. and L.J. Mandel, 1984. Active ion transport in the renal proximal tubule. III. The ATP dependence of the Na pump. J. Gen. Physiol., 84: 643-662.
PubMedDirect Link - Vaandrager, A.B. and H.R. de Jonge, 1996. Signalling by cGMP-dependent protein kinases. Dev. Mol. Cell. Biochem., 17: 23-30.
CrossRefDirect Link - Verrey, F., J. Beron and B. Spindler, 1996. Corticosteroid regulation of renal Na,K-ATPase. Miner. Electrolyte Metab., 22: 279-292.
PubMedDirect Link - Wang, Z.M., M. Yasui and G. Celsi, 1995. Differential effects of glucocorticoids and mineralocorticoids on the mRNA expression of colon ion transporters in infant rats. Pediatr. Res., 38: 164-168.
CrossRefDirect Link - Whorwood, C.B. and P.M. Stewart, 1995. Transcriptional regulation of Na/K-ATPase by corticosteroids, glycyrrhetinic acid and second messenger pathways in rat kidney epithelial cells. J. Mol. Endocrinol., 15: 93-103.
CrossRefPubMedDirect Link - Whorwood, C.B., M.L. Ricketts and P.M. Stewart, 1994. Regulation of sodium-potassium adenosine triphosphate subunit gene expression by corticosteroids and 11 beta-hydroxysteroid dehydrogenase activity. Endocrinology, 135: 901-910.
CrossRefPubMedDirect Link - Thomas, R., P. Gray and J. Andrews, 1990. Digitalis: Its Mode of Action, Receptor and Structure-Activity Relationships. In: Advances in Drug Research, Volume 19, Testa, B. (Ed.). Academic Press, San Diego, CA., USA., ISBN: 978-0-12-013330-7, pp: 311-562.
Direct Link - Bertorello, A. and A. Aperia, 1990. Short-term regulation of Na+,K+-ATPase activity by dopamine. Am. J. Hypertens., 3: 51S-54S.
PubMedDirect Link - Bertorello, A. and A. Aperia, 1989. Regulation of Na+-K+-ATPase activity in kidney proximal tubules: Involvement of GTP binding proteins. Am. J. Physiol.-Renal Physiol., 256: F57-F62.
PubMedDirect Link