
Rama Krishna Devaki
Department of Biochemistry, Kamineni Institute of Medical Sciences, Sreepuram, Narketpally, Nalgonda
M.N. Subhash
Department of Neurochemistry, National Institute of Mental Health and Neurosciences, Bangalore, India
Asian Journal of Biochemistry
Year: 2014 | Volume: 9 | Issue: 1 | Page No.: 49-56







ABSTRACT
Sumatriptan is a novel and highly effective drug against migraine and cluster headache attacks. This antimigraine drug is known to exert its effect through the modulation of serotonin (5-HT) mediated neurotransmission. 5-HT1B receptors in rats and 5-HT1D receptors in humans have been thought to be likely targets of sumatriptan. In the present study the effect of varying dose and duration of sumatriptan on the density of 5-HT1B receptors in the rat cerebellum were studied to understand its mechanism of action. Sprague Dawley rats were administered with different doses of sumatriptan (0.2-16 mg kg-1 b.wt. i.p.) for seven days and 2, 4 and 8 mg kg-1 of sumatriptan for 7, 14 and 21 days. The radioligand binding assays were performed in cerebellar membranes using [3H]5-HT. Treatment with different doses of sumatriptan for seven days showed a significant (p<0.0001) downregulation of 5-HT1B receptors in a dose dependent manner. A significant decrease in the density of 5-HT1B receptors was observed with 0.2 mg (45%), 0.5 mg (72%) and 1 mg (70%) of sumatriptan treatment, with a significant decrease (p<0.001) in Kd values. No further decrease in either the density or in the Kd values was observed with increasing doses of Sumatriptan from 2 to 16 mg kg-1 b.wt. The magnitude of decrease in the receptor density was more significant with a lower dose (2 mg) of sumatriptan for a prolonged period of exposure (21 days). However, such change was not observed with higher doses (4 and 8 mg kg-1). Furthermore, sumatriptan showed a higher affinity for 5-HT1B receptors with a Ki value of 9.4±0.9 nM, when compared to other agonists and antagonists. Taken together, these findings suggest that desensitization of 5-HT1B receptor is dose and time dependent, which may be an important factor underlying the mechanism of action of sumatriptan as an antimigraine drug.
PDF Abstract XML References Citation
Received: November 15, 2013;
Accepted: November 23, 2013;
Published: March 04, 2014
How to cite this article
Rama Krishna Devaki and M.N. Subhash, 2014. Differential Effect of Sumatriptan on Cerebellar 5-HT1B Receptors
in Rat Brain. Asian Journal of Biochemistry, 9: 49-56.
DOI: 10.3923/ajb.2014.49.56
URL: https://scialert.net/abstract/?doi=ajb.2014.49.56
DOI: 10.3923/ajb.2014.49.56
URL: https://scialert.net/abstract/?doi=ajb.2014.49.56
INTRODUCTION
Neurogenic inflammation within the meninges has been shown as an important event in the pathogenesis of migraine and headache (Millson, 2004) and evidence shows that 5-HT1B/1D ligands may be useful in the treatment of migraine (Peroutka, 1990; Burstein and Jakubowski, 2004). Sumatriptan is a novel and highly effective drug against migraine and cluster headache attacks. It shows a remarkable pharmacological profile in animals. 5-HT1D receptors, pharmacologically defined functional sites in human brain, exhibit high affinity for sumatriptan (Weinshank et al., 1992). So far 5-HT1B receptors in rats and 5-HT1D receptors in guinea pigs and humans have been thought to be the likely target of sumatriptan (Moskowitz, 1992). The amino acid sequence within the transmembrane domains of 5-HT1D receptors is found to be 96% identical with that of 5-HT1B receptors (Hoyer et al., 1994). 5-HT1F receptors in guinea pig brain have also been shown to have high affinity for sumatriptan (Waeber and Moskowitz, 1995).
Drugs acting via 5-HT1B/1D receptors are effectively used to treat migraine. However, the antinociceptive effects of many such agents have not been understood. Sumatriptan may act on the cerebral arteries by blocking neurogenic inflammation and nociceptive activity within trigemino-vascular afferents (Burstein and Jakubowski, 2004). This action has been argued to be due to the activation of 5-HT1B/1D receptors, which prevents protein extravasation induced by trigeminal ganglion (Reuter et al., 2004). It has been shown as early as in 1989 that sumatriptan has high affinity for 5-HT1D and 5-HT1B binding sites (Peroutka and McCarthy, 1989). It selectively interacts with 5-HT1B sites, which may be the basis for its apparent efficacy in the acute treatment of migraine (Peroutka and McCarthy, 1989). Sumatriptan has been shown to inhibit neurogenic inflammation dose dependently via 5-HT1D receptors in guinea pigs and via 5-HT1B receptors in rats (Xian-Jie et al., 1997). Blockade of dural extravasation is concomitant with an action at rat 5-HT1D receptors, while inhibition of dural vasodilation is concomitant with 5-HT1B receptors. Apart from mediating inhibition of K+ evoked release of 5-HT (Pineyro et al., 1995; Pineyro and Blier, 1996), sumatriptan is also known to increase growth hormone (GH) levels as well as its response to growth hormone releasing hormone (Mota et al., 1995) and activates pituitary-adrenal axis (Facchinetti et al., 1994). Study of the mechanism of action of sumatriptan via 5-HT1B/1D receptors would further our understanding of the pathophysiology of migraine and the putative involvement of 5-HT receptors.
Although, migraine is inextricably bound with 5-HT receptors and its many subtypes, its precise role continues to elude the scientists and there is still no clear evidence suggesting either vascular or neurogenic hypothesis, unequivocally. Therefore, the present study was designed to investigate the dose and duration dependent effects of sumatriptan administration on the density of 5-HT1B receptors in rat cerebellum.
MATERIALS AND METHODS
Chemicals: [3H]5-HT (s.a. 8.2 Ci mmol-1) was obtained from M/s. Amersham Plc. UK. 8-hydroxy-2 (di-n-propylamino) tetralin (8-OH-DPAT), mianserin, 5-HT, N-3[(Tri-fluoromethyl) phenyl] piperazine (TFMPP) and pargyline were obtained from M/s. Sigma Chemicals, MO, USA. Sumatriptan was a generous gift from M/s. Torrent Pharmaceuticals, India. GF/B filters were obtained from M/s. Millipore, UK.
Animals and administration of drugs: Adult male Sprague Dawley rats (n=6 for each treatment group), weighing 200-240 g, were procured from Central Animal Research Facility (CARF) of NIMHANS. Rats were housed four per cage with food and water ad libitum in a well-ventilated room, with 12 h light and dark cycles. Rats were injected, once in the morning, with different doses of sumatriptan (0.2, 0.5, 1.0, 2.0, 4.0, 8.0, 12.0 and 16.0 mg kg-1 b.wt., i.p.) for seven days. In another experiment, rats were injected (i.p.) once in the morning, with different doses (2, 4 and 8 mg kg-1 b.wt.) of sumatriptan, each dose for 7, 14 and 21 days. Control rats were similarly treated with saline and housed under identical conditions. Animals were sacrificed 24 h after the last dose and brains removed. The cerebellum was dissected from each rat brain and immediately used for the assay. All animal experimental procedures were performed in accordance with local guidelines for the Care and Use of Laboratory Animals and approved by the institute’s Animal Ethics Committee.
Membrane preparation and receptor binding assay: Crude synaptic membranes were isolated from the cerebellar tissues as described earlier (Subhash et al., 1998) for the radioligand binding studies. The membrane pellet was suspended in 50 mM Tris-HCl buffer (pH 7.4) containing 10 μM pargyline, 4 mM CaCl2 and 0.1% ascorbic acid. The protein content of the membrane pellet was determined by Lowry’s method (Lowry et al., 1951). The saturation experiments for 5-HT1B receptors were done with [3H]5-HT (0.2-2.0 nM) in presence of 100 nM 8-OH-DPAT and 3000 nM mianserin, to block 5-HT1A and 5-HT2 receptors, respectively, essentially following the procedure described earlier (14). Non-specific binding of the radioligand was obtained using 100 nM TFMPP, a selective 5-HT1B receptor agonist. Drug displacement studies were done by incubating cerebellar membranes obtained from control rats with 2 nM of [3H] 5-HT, in presence of 100 nM 8-OH-DPAT and 3000 nM mianserin and with different concentrations of drugs (10-11-10-4 M). The binding data were analyzed using LIGAND program (McPherson, 1983) to obtain the equilibrium dissociation constant (Kd), the density of receptor (Bmax), the Hill co-efficient (n), inhibitor constant (Ki) and the concentration of the inhibitor at half of the maximal binding (IC50).
Statistical analysis: Statistical analysis was performed using Sigma Stat software (SPSS, Inc., Chicago, IL). The differences in binding parameters between and among the groups were analyzed using one-way ANOVA and p-values were considered significant at p<0.05.
RESULTS
Acute sumatriptan exposure: The density of cerebellar 5-HT1B receptors in saline treated rats was 60.3±3.8 fmol mg-1 protein with a Kd of 0.96±0.02 nM. Seven days of treatment with different doses of sumatriptan showed a significant (Fdf59 = 326.6, p<0.0001) decrease in the density of 5-HT1B receptors in a dose dependent manner (Table 1). A significant decrease in the density of these receptors was seen with 0.2 mg (45%), 0.5 mg (72%) and 1.0 mg (70%) of sumatriptan treatment (Fig. 1). No further decrease was observed with increasing doses of sumatriptan from 2 to 16 mg. The affinity of [3H]5-HT to 5-HT1B receptors was increased significantly with 0.5 and 1 mg dose, as evident from the decreased Kd values. However, no further change in the affinity was observed with further increase in the dose of sumatriptan (2 to 16 mg kg-1 b.wt.).
| Table 1: | In vivo effect of Sumatriptan administration for 7 days on cerebellar 5-HT1B receptors in rat brain |
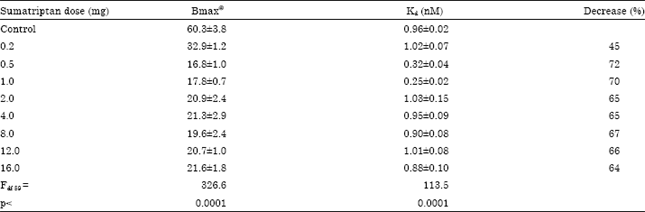 | |
| @ f mole mg-1 protein, Values are mean and SD of 3-4 experiments, each assayed in duplicate. The density of 5-HT1B receptors was estimated using [3H]5-HT, as described, in membranes obtained from cerebellum of rats treated with different doses (0.2-16 mg kg-1 b.wt.) of sumatriptan for 7 days | |
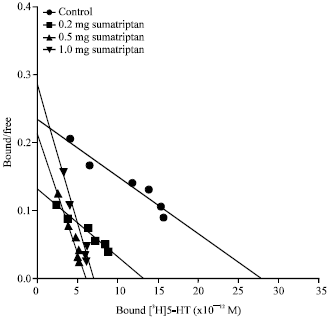 | |
| Fig. 1: | Scatchard plot of [3H]5-HT binding to 5-HT1B receptors in cerebellar membranes of rats Administered (i.p) with 0.2, 0.5 and 1.0 mg kg-1 b.wt. of sumatriptan 7 days and control rats. Values are mean of 3-4 experiments, each assayed in duplicate |
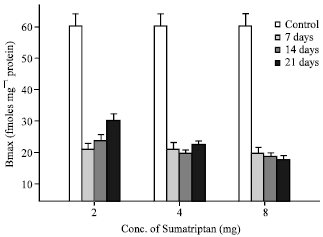 | |
| Fig. 2: | 5-HT1B receptor density was estimated, as described, in cerebellar membranes of rats treated with various doses (2, 4 and 8 mg kg-1 body wt, i.p.) of sumatriptan for 7, 14 and 21 days, Values are mean and SD of 3-4 experiments, each assayed in duplicate, *the percentage of downregulation of receptor with 2 mg kg-1 b.wt. dose, was significantly different with duration (Fdf23 = 34.9; p<0.0001) |
Chronic sumatriptan exposure: Rats were also treated with 2, 4 and 8 mg kg-1 b.wt. of sumatriptan for varied duration (7, 14 and 21 days). When compared to control levels there was a significant decrease in the density of 5-HT1B receptors with 2, 4 and 8 mg kg-1 doses, when rats were administered for 7, 14 and 21 days (Fig. 2). However, with 2.0 mg kg-1 dose, there was a significant (Fdf23 = 34.9; p<0.0001) up regulation of 5-HT1B receptors at 21 days of exposure, when compared to 7 and 14 days of exposure (Table 2). There was however, no change in the receptor affinity.
| Table 2: | Effect of various doses and durations of sumatriptan administration on cerebellar 5-HT1B receptors in rat brain |
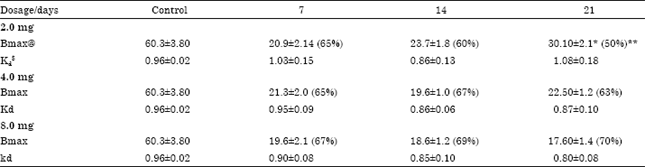 | |
| @ f moles mg-1 protein , $ nM, Values are mean and SD of 3-4 experiments, each assayed in duplicate, The density of 5-HT1B receptors was estimated using [3H]5-HT, as described, in the cerebellar membranes obtained from rats treated with different doses (2, 4, 8 mg kg-1 b.wt.) of sumatriptan for 7, 14 and 21 days, *Significance compared with 7 days (Fdf 23 = 34.9, p<0.0001), **% downregulation compared to control levels | |
This trend, however, was not observed with higher doses (4 and 8 mg kg-1), where there was no significant change in the receptor density after 14 and 21 days of sumatriptan treatment, when compared to 7 days. Drug displacement studies revealed that sumatriptan has a high affinity towards 5-HT1B receptors with a Ki of 9.4±0.9 nM and an IC50 of 28.7±1.4 nM. When compared to sumatriptan the affinity for other agents was in the order of TFMPP (Ki = 29.5±2.1 nM)>mCPP (Ki = 108.0±15.1 nM)>pindolol (Ki = 148.0±17.6 nM)>5-CT (Ki = 272.0±2.6 nM).
DISCUSSION
In view of the potential pathophysiological and therapeutic implications, presynaptic 5-HT receptors have been extensively studied by molecular biological and pharmacological techniques. Studies have established that the terminal auto receptors controlling 5-HT release in the rat brain are of 5-HT1B types. The 5-HT1B receptors were originally thought to exist only in rodents. Later they have been characterized and found to be identical with 5-HT1Dβ receptors found in higher species (Hoyer et al., 1994). 5-HT1D receptors (5-HT1Dα)) have 63% overall structural homology and 77% amino acid sequence homology with 5-HT1B (5-HT1Dβ) receptors. Rat brain has very low levels of 5-HT1D mRNA. Similar to 5-HT1A receptors, mRNA for both 5-HT1B and 5-HT1D subtypes are found in Dorsal Raphe Nucleus (DRN), suggesting that non-5HT1A receptors like 5-HT1B receptors, with very small amount of 5-HT1D component, exist in DRN (Stamford et al., 2000). Both 5-HT1Dα and 5-HT1Dβ receptors have also been shown to exhibit high affinity for sumatriptan (Weinshank et al., 1992). In the rat dura mater, 5-HT1Dα and 5-HT1Dβ receptors mediate the inhibitory effects of sumatriptan, suggesting the role of 5-HT1B receptors in the mechanism of action of sumatriptan. This finding was further supported by the report that showed lack of sumatriptan activity in the 5-HT1B receptors knockout mice (Yu et al., 1996). The 5-HT1B receptors in rats and 5-HT1D receptors in guinea pigs have been thought to be the relevant targets of sumatriptan within the dura mater (Matsubara et al., 1991). Pre junctional action of 5-HT1 receptors on trigemino vascular axons but not the post junctional action, like constriction of vascular smooth muscle, mediates the effect of sumatriptan (Moskowitz et al., 1991). Inhibitory 5-HT1B receptors have been shown to be present in cat brain and 5-HT1B receptors mediated inhibition has been suggested to be the most potent action in the inhibition of trigemino vascular nociceptive traffic (Goadsby and Classey, 2003).
Since in rats 5-HT1B receptors are the targets of sumatriptan, the dose and duration dependent in vivo effects of sumatriptan on the density of cerebellar 5-HT1B receptors were studied in rat brain. The present study demonstrated a dose dependent down regulation of 5-HT1B receptors in the cerebellum at lower doses of sumatriptan. A dose of 1 mg was sufficient to down regulate nearly 70% of the receptors. Interestingly, no further down regulation of 5-HT1B receptors was observed with increasing dose. However, with increasing duration of exposure, a significant reduction in the down regulation of 5-HT1B receptors (from 65% to 50%) was seen at a dose of 2 mg. However, this trend was not seen at 4 and 8 mg kg-1 b.wt. doses. The significant decrease in Kd values is concomitant with significant decrease in the density of 5-HT1B receptors at 0.5 and 1.0 mg kg-1 dose of sumatriptan. However, the affinity was same as that of control with further increase in dosage. It was observed that the affinity of [3H5-HT to 5-HT1B receptors increases with increase in downregulation.
As there was no further increase in downregulation with increasing dosage, the affinity was not altered. These findings suggest that administration of lower doses (2 mg) for a moderate period (2 weeks) would be critical for a significant downregulation of cerebellar 5-HT1B receptors, which may be involved in the therapeutic action of sumatriptan and higher doses for a longer period may be required for the prophylactic activity.
After intra vascular administration of sumatriptan, rats Johnson et al. (2001) have shown that sumatriptan has high affinity agonistic activity at 5-HT1D/B receptors, though less than Zolmitriptan. This weaker functional 5-HT1B receptor agonistic potency of sumatriptan when compared to Zolmitriptan is however, not due to its low concentration in the brain (Johnson et al., 2001). In a recent study, a significant decrease (40%) in 5-HT1B/1D receptor mRNA in trigeminal ganglion was reported after 14 days of treatment with sumatriptan (7). Furthermore, the drug displacement study done in our laboratory has revealed that sumatriptan has a high affinity (Ki = 9.4 nM) to 5-HT1B receptors in rat cerebellum, when compared to TFMPP, a 5-HT1B agonist, mCPP, a 5-HT2B/2C agonist, pindolol, a 5-HT1A/1B antagonist and 5-CT, a 5-HT1A/1B/1D agonist.
In another study in rats, the dose relation and complete inhibition of plasma protein extravasation in the duramater was found to be similar with both sumatriptan and electriptan, when injected intra venously (Gupta et al., 2000). This suggests that the effectiveness of this drug, which displays both high affinity and selectivity to 5-HT1B/1D receptors, may indicate the importance of this specific 5-HT1-receptor subtype in the pathogenesis of migraine. Experiments with new 5-HT1B and 5-HT1D subtype selective ligands are required to confirm these findings. It is note worthy that 5-HT1B receptors also sub serve the role of heteroreceptors. Future studies are needed to replicate findings in various brain regions using more specific radioligands.
Prevention of dural vasodilation involves activation of trigeminal Aδ-fibers, whereas prevention of dural extravasation involves activation of trigeminal C-fibers. So, the differential effect of sumatriptan may be due to different receptor subtypes involved and to their location and species studied. Taken together, the desensitization of 5-HT1B receptors in rat brain, which are identical to 5-HT1D receptors in human brain, may underlie the mechanism of antimigraine properties of sumatriptan, wherein the neurogenic inflammation on terminals of trigeminal neurons in dural vessels may be inhibited.
ACKNOWLEDGMENTS
Authors are grateful to NIMHANS, Bangalore, India, for the research fellowship and Kamineni Institute of Medical Sciences for constant support and encouragement.
REFERENCES
- Peroutka, S.J., 1990. The pharmacology of current anti-migraine drugs. Headache: J. Head Face Pain, 30: 5-11.
CrossRef - Burstein, R. and M. Jakubowski, 2004. Analgesic triptan action in an animal model of intracranial pain: A race against the development of central sensitization. Ann. Neurol., 55: 27-36.
CrossRef - Weinshank, R.L., J.M. Zgombick, M.J. Macchi, T.A. Branchek and P.R. Hartig, 1992. Human serotonin 1D receptor is encoded by a subfamily of two distinct genes: 5-HT1Dα and 5-HT1Dβ. Proc. Natl. Acad. Sci., 89: 3630-3634.
Direct Link - Moskowitz, M.A., 1992. Neurogenic versus vascular mechanisms of sumatriptan and ergot alkaloids in migraine. Trends Pharm. Sci., 13: 307-311.
CrossRefDirect Link - Waeber, C. and M.A. Moskowitz, 1995. [3H] sumatriptan labels both 5-HT1D and 5-HT1F receptor binding sites in the guinea pig brain: An autoradiographic study. Naunyn-Schmiedeberg's Arch. Pharmacol., 352: 263-275.
CrossRefDirect Link - Reuter, U., S. Salomone, G.W. Ickenstein and C. Waeber, 2004. Effects of chronic sumatriptan and zolmitriptan treatment on 5-HT1 receptor expression and function in rats. Cephalalgia, 24: 398-407.
CrossRefDirect Link - Peroutka, S.J. and B.G. McCarthy, 1989. Sumatriptan (GR 43175) interacts selectively with 5-HT 1B and 5-HT 1D binding sites. Eur. J. Pharm., 163: 133-136.
CrossRefDirect Link - Mota, A., A. Bento, A. Penalva, M. Pombo and C. Dieguez, 1995. Role of the serotonin receptor subtype 5-HT1D on basal and stimulated growth hormone secretion. J. Clin. Endocrinol. Metab., 80: 1973-1977.
Direct Link - Facchinetti, F., R.E. Nappi, G. Sances, L. Fioroni, G. Nappi and A.R. Genazzani, 1994. The neuroendocrine effects of sumatriptan, a specific ligand for 5-HT 1-like receptors. Clin. Endocrinol., 40: 211-214.
CrossRef - Subhash, M.N., B.N. Srinivas, K.Y. Vinod and S. Jagadeesh, 1998. Inactivation of 5-HT1A and [3H] 5-HT binding sites by N-ethoxycarbonyl-2-ethoxy-1, 2-dihydroquinoline (EEDQ) in rat brain. Neurochem. Res., 23: 1321-1326.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Stamford, J.A., C. Davidson, D.P. McLaughlin and S.E. Hopwood, 2000. Control of dorsal raphe 5-HT function by multiple 5-HT 1 autoreceptors: Parallel purposes or pointless plurality? Trends Neurosci., 23: 459-465.
CrossRef - Yu, X.J., C. Waeber, N. Castanon, K. Scearce and R. Hen et al., 1996. 5-carboxamide-tryptamine, CP-122-288 and dihydroergot-amine but not sumatriptan, CP-93-129 and serotonin-5-o-carboxymethyl-glycyl-tyrosinamide block dural plasma protein extravasation in knockout mice that lack 5-hydroxytryptamine 1B receptors. Mol. Pharamacol., 49: 761-765.
- Matsubara, T., M.A. Moskowitz and B. Byun, 1991. CP-93,129, a potent and selective 5‐HT1B receptor agonist blocks neurogenic plasma extravasation within rat but not guinea‐pig dura mater. Br. J. Pharmacol., 104: 3-4.
CrossRefDirect Link - Goadsby, P.J. and J.D. Classey, 2003. Evidence for serotonin (5-HT)1B, 5-HT 1D and 5-HT 1F receptor inhibitory effects on trigeminal neurons with craniovascular input. Neuroscience, 122: 491-498.
CrossRefDirect Link - Johnson, D.E., H. Rollema, A.W. Schmidt and A.D. McHarg, 2001. Serotonergic effects and extracellular brain levels of eletriptan, zolmitriptan and sumatriptan in rat brain. Eur. J. Pharmacol., 425: 203-210.
CrossRefDirect Link