
S.M. El-Bahr
Department of Physiology, Biochemistry and Pharmacology (Biochemistry), College of Veterinary Medicine and Animal Resources, King Faisal University, Saudi Arabia
Asian Journal of Biochemistry
Year: 2014 | Volume: 9 | Issue: 1 | Page No.: 30-40







ABSTRACT
The objective of the present study was to investigate the effect of camel milk on activities and gene expression of hepatic antioxidant enzymes in rats intoxicated with carbon tetrachloride (CCl4). Therefore, twenty four rats were used in the current study. Rats were divided into four groups, the first and second groups were received water and camel's milk, respectively whereas rats of both third and fourth groups were injected with CCl4 and received water and camel's milk, respectively. After 5 weeks, liver tissues were collected for biochemical analysis of the activities and gene expression of antioxidant enzymes. Rats supplemented with camel milk alone showed no significant difference in all examined parameters compared to control rats. Liver damage and oxidative stress were evident in untreated CCl4 intoxicated rats as indicated by significant elevation of hepatic enzymes, significant elevation in thiobarbituric acid reactive substance (TBARS), significant reduction in reduced glutathione level (GSH), significant reduction in the activities of catalase (CAT), superoxide dismutase (SOD), glutathione peroxidase (GPX) and glutathione-S transferase (GST) and finally significant down-regulation of antioxidant enzymes gene expression compare to control. Administration of camel milk along with CCl4 caused amelioration in CCl4-induced effects compare to the untreated CCl4 intoxicated rats via up-regulation of antioxidant enzyme gene expression, activation of the expressed genes and increasing the availability of GSH. Conclusively, camel milk exerted its protective effect against CCl4 induced hepatic toxicity by modulating the extent of lipid peroxidation and augmenting antioxidant defense system at activity and gene expression levels.
PDF Abstract XML References Citation
Received: September 07, 2013;
Accepted: December 20, 2013;
Published: March 04, 2014
How to cite this article
S.M. El-Bahr, 2014. Camel Milk Regulates Gene Expression and Activities of Hepatic Antioxidant Enzymes in Rats Intoxicated with Carbon Tetrachloride. Asian Journal of Biochemistry, 9: 30-40.
DOI: 10.3923/ajb.2014.30.40
URL: https://scialert.net/abstract/?doi=ajb.2014.30.40
DOI: 10.3923/ajb.2014.30.40
URL: https://scialert.net/abstract/?doi=ajb.2014.30.40
INTRODUCTION
CCl4 is one of the most toxic agent to the liver tissue and its hepatotoxicity is due to trichloromethyl radical which produced during oxidative stress (Stoyanovsky and Cederbaum, 1999). These radicals stimulate the release of hepatic macrophages, kupffer cells and accompanied increase of infiltrated neutrophils and lymphocytes (Ramadori and Saile, 2004). The activated macrophages are released and contributed to liver fibrosis, inflammation and injury (Canbay et al., 2004; Saile and Ramadori, 2007) with inefficient remedy (Lee et al., 2007). Although, progress in the treatment of liver disease by chemical drugs has been reported, searching for natural drug is still ongoing (Recknagel, 1983). Currently, providing therapeutically effective natural drug for the treatment of liver diseases is the main duty of complementary and alternative medicines. Bioactive peptides and proteins contents of camel's milk are important for maintain effective bioprocesses such as digestion, growth and immune responses (Yagil et al., 1984; Korhonen and Pihlanto, 2003). In addition, camel’s milk has the advantage of being stored at room temperature for longer period than other milk (Omer and Eltinay, 2009). The most described uses of camel's milk are reported as drug against autoimmune diseases, dropsy, jaundice, spleenomegaly, tuberculosis, asthma, anemia, piles, diabetes (Rao et al., 1970) and as antimicrobial agent (El Agamy et al., 1992). In addition, camel’s milk has antitoxic effect against cadmium chloride (Al-Hashem et al., 2009; Dallak, 2009), CCl4 (Khan and Alzohairy, 2011), Cisplatin (Afifi, 2010), Paracetamol (Al-Fartosi et al., 2011), Aluminum chloride (Al-Hashem, 2009). It has been reported that, camel milk ameliorates CCl4 induced oxidative stress via regulation of antioxidant enzyme activities (Al-Fartosi et al., 2012). However, publications reported the gene expression regulation of these enzymes by camel milk are lack so far. Therefore, the aim of the present study was to investigate the effect of camel milk on oxidative damage and oxidative stress related gene expression and activities in rat liver intoxicated withCCl4.
MATERIALS AND METHODS
Chemicals and kits: Paraffin oil, CCl4 (Spectrosol® BHD chemicals ltd pool, England) and other chemicals and solvents were of highest grade commercially available. EGTA, EDTA, sucrose, Tris, butanol, mannitol, metaphosphoric and H2O2 were purchased from Sigma Chemical Co. (St. Louis, MO, USA). RIPA buffer was provided by Cayman chemical company, USA. All other chemicals were of analytical grade. Diagnostic kits for serum alanine aminotransferase (ALT) and aspartate amino transferase (AST) were purchased from ELIPSE, United diagnostic industry, UDI, Dammam, Saudi Arabia.
Camel’s milk: Camel’s milk samples were collected daily early in the morning from camel farm in Camel Research Center, King Faisal University, Al-Ahsa, Saudi Arabia. Milk was collected from camels by hand milking. The samples were collected in sterile screw bottles and kept in cool boxes until transported to the laboratory. The rats were given fresh milk (100 mL 24 h-1 cage-1) (Althnaian et al., 2013) as such without any further treatment.
Animals and treatment: A total of 24 albino rats (200-250 g) were obtained from Laboratory House of College of Veterinary Medicine and Animal Resources, King Faisal University, Al-Ahsa, Saudi Arabia and acclimated for 10 days before starting the experiment. All animals were housed in standard cages (6 rats cage-1), feeding with standard laboratory diet and tap water ad libitum. The experimental animals were housed in air-conditioned rooms at 21-23°C and 60-65% of relative humidity and kept on a 12 h light/12 h dark cycle. The animals received humane care in accordance with the Guide for the Care and Use of Laboratory Animals, published by ethics of scientific research committee of King Faisal University, Saudi Arabia.
Induction of hepatotoxicity by CCl4: Liver toxicity was induced by the intraperitoneal injection of CCl4 (1 mL kg-1 b.wt.), 1:1 diluted with paraffin oil, for two successive days of the experiment (Khan and Alzohairy, 2011).
Experimental groups and protocol: The rats were divided randomly into 4 groups comprising 6 rats in each group and fed the same diet throughout the experimental period. The experimental design is described as fellow:
| Group 1: | Rats fed only with basal diet and water and injected i/p with Paraffin oil, this group was served as control 1 group |
| Group 2: | Rats fed normal basal diet, injected i/p with Paraffin oil and treated with camel's milk (100 mL 24 h-1 cage-1) (Althnaian et al., 2013) as their sole source of drinking water, this group was served as control 2 group |
| Group 3: | Rats fed basal diet and water and intoxicated with CCl4 (1 mL kg-1 b.wt.), 1:1 diluted with paraffin oil on first two days of the experiment (Khan and Alzohairy, 2011) |
| Group 4: | Rats fed basal diet and intoxicated with CCl4 (1 mL kg-1 b.wt.), 1:1 diluted with paraffin oil on first two days of the experiment and then treated with camel's milk (100 mL 24 h-1 cage-1) as their sole source of drinking water |
Sample collection: After 5 weeks, all rats were anesthetized with diethyl ether. Blood samples were collected by cardiac puncture before incision of the abdomen; 5 mL of blood samples were collected in plain tubes, serum was collected and frozen at -30°C until the time of analysis of liver enzymes using commercial assay kits according to the manufacture instruction. The liver tissues were removed and liver fragments were immediately frozen in liquid nitrogen and stored at -80°C for molecular and biochemical analysis of antioxidant enzymes. Because of the same liver tissues of the previously published work (Althnaian et al., 2013) were used, the methodology of liver histogram was not mentioned in the current work.
Assessment of liver damage: Commercial diagnostic kits (United Diagnostic Industry, UDI, Dammam, Saudi Arabia) were used for determination of ALT (EP07-500) and AST (EP15-500) on ELIPSE full automated chemistry analyzer (Rome, Italy). The hitopathological picture of the previous work (Althnaian et al., 2013) was taken in consideration.
Determination of hepatic antioxidant enzymes, thiobarbituric acid reactive substances (TBARS) and reduced glutathione: One gram of liver tissues was homogenized in 5 mL of cold 20 mM HEPES buffer, pH 7.2, containing 1 mM EGTA, 210 mM mannitol and 70 mM sucrose. After centrifugation (1500xg/5 min) at 4°C, the supernatant was removed and stored frozen at -80°C until the time of analysis of superoxide dismutase (SOD). Another one gram of liver tissues was homogenized in 5 mL of cold buffer of 50 mM potassium phosphate buffer, pH 7, containing 1 mM EDTA. After centrifugation (10.000xg/15 min) at 4°C, the supernatant was removed and stored frozen at -80°C until the time of analysis of catalase (CAT), glutathione peroxidase (GPX), glutathione-S transferase (GST) and reduced glutathione (GSH). The extent of lipid peroxidation in terms of TBARS formation was measured by mixing one gram of liver tissues with RIPA buffer (Item No. 10010263, Cayman chemical company, USA). After homogenization, sonication and centrifugation (1600xg/10 min), the supernatant was removed and stored frozen at -80°C until the time of analysis. The activities of CAT (nmol min-1 g-1 tissue; Cayman Chemical Company, USA, Catalog No. 707002), GPX (nmol min-1 g-1 tissue; Cayman Chemical Company, USA, Catalog No.703102), SOD (U g-1 tissue; Cayman Chemical Company, USA, Catalog No. 706002), GST (nmol min-1 g-1 tissue; Cayman Chemical Company, USA, Catalog No. 703302) and concentrations of GSH (μM; Cayman Chemical Company, USA, Catalog No. 703002) and TBARS (μM; Cayman Chemical Company, USA, Catalog No. 10009055) were determined by ELISA reader (Absorbance Microplate Reader ELx 800TM BioTek®, USA). Results were calculated according to the manufacture instructions.
Total RNA isolation and real time RT-PCR of hepatic antioxidant enzymes: Liver tissues (approximately 1 g of tissue per sample) were immediately added to 1 mL of TriZol reagent (Invitrogen, Carlsbad, CA) and homogenized using homogenizer (Tissue Ruptor, Qiagen GmbH, Germany). One milliliter of the tissue homogenate was transferred to a microfuge tube and total RNA was extracted by adding 0.2 mL chloroform. Afterwards, samples were vortexed vigorously for 15 sec and incubated at room temperatuer for 3 min. After centrifugation (12,000 g 15 min-1) at 4°C, the aqueous phase containing RNA was transfered into new tubes. RNA was precipitated by mixing the aqueous phase with 0.5 mL isopropyl alcohol and incubated at room temperuter for 10 min. After centrifugation at 12,000 g for 10 min at 4°C, RNA pellets were washed by mixing and vortexing with 1ml of 75% ethanol. After centrifugation (7.500 g 5 min-1) at 4°C, RNA pellets were resuspended in nuclease free water (Life Technologies. USA). The purity of RNA at 260/280 OD ratio and RNA integrity was evaluated using Multi-Mode Microplate reader (SYNERGY Mx, BIO-TEK. Winooski, Vermont, USA). Only high purity samples (OD260/280 >1.8) were subjected to further manipulation. cDNA was prepared from RNA samples according to Revers Transcription System Kit (Promega, Madison, USA) by using Bio-Rad Thermal Cycler (T100TM, Foster city, California, USA). Briefly, total RNA were activated at 70°C for 10 min and 20 μL reaction mix were made of 4 μL MgCl2, 2 μL of reverse transcription 10X buffer, 2 μL of dNTP mixture (10 mM), 0.5 μL of random primers, 0.75 μL of AMV reverse transcriptase enzyme, 1ng RNA and nuclease-free water to a final volume of 20 μL. Then the reaction was incubated at 42°C for 60 min followed by incubation at 94°C for 5 min. cDNA was diluted up to 100 μL with nuclease-free water for PCR amplification. Real time RT-PCR was performed using QuantiFastTM SYBR Green PCR Master Mix kit (QIAGEN, Hilden; Germany). The 25 μL reaction for each examined gene was prepared from 12.5 μL of master mix; 2 μL forwerd primer (10 pmol); 2 μL revers primer (10 pmol); 2 μL cDNA of the sample and 6.5 μL of nuclease-free water. Cycling parameters were, 50°C for 2 min, 95°C for 15 min, 40 cycles of 95°C for 10 sec, followed by 55°C for 30 sec and 72°C for 10 sec with final melting at 95°C for 20 sec. For each gene examined, duplicate samples from each cDNA analyzed by real time RT-PCR using the Bio-Rad CFX Manager 3.0 Software of the C1000 Touch thermal cycler-CFX96 Real time PCR(BIO-RAD, Foster city, California, USA). The â-actin mRNA fragment was used as housekeeping gene to normalize the expression data. The primer sequences are described in Table 1.
Statistical analysis: All data was presented as Mean±Standard error of mean by using student-ttest. All tests were performed using computer package of the statistical analysis system (SAS, 2002). The relative gene expression of target genes in comparison to the β-actin reference gene was calculated using the Bio-Rad CFX Manager 3.0 Software of the C1000 Touch thermal cycler-CFX96 Real time PCR(BIO-RAD, Foster city, California, USA).
| Table 1: | Details giving primer sequences and expected product size for the genes amplified |
 | |
| SOD: Superoxide dismutase, CAT: Catalase, GPX: Glutathione peroxidase, GST: Glutathione-S transferase | |
RESULTS
Assessment of liver function: The activities of AST and ALT were estimated in serum samples as the liver function biomarkers. These results are given in Table 2. The CCl4 treatment markedly affected the liver specific enzymes. It was found that a significant (p<0.05) increase in serum AST(130.8±01.7 U L-1) and ALT (47.4±4.0 U L-1) activities of CCl4 treated rats compare to control (118.9±06.0 and 29.2±0.1 U L-1), respectively. This result suggests that these hepatic biomarkers are elevated in the serum due to release of the enzymes from damaged liver. However a significant decrease (p<0.05) was observed in the respective serum activities of rats given Camel milk+CCl4 (125.6±2.0 and 34.7± 1.5 U L-1) compared with CCl4 treated group(130.8±01.7 and 47.4±4.0 U L-1), respectively. As this work is a continuation to the previous published work (Althnaian et al., 2013), inclusion of histopathological picture in this study was not necessary because the same liver samples were used. The previous study had demonstrated that, liver of CCl4-intoxicated rats showed massive fatty change and centrilobular necrosis in most cases. Hepatitis characterized by mononuclear cells infiltration mostly macrophages and lymphocytes around central veins and in portal areas was also noticed in most cases of CCl4 intoxicated rats. In addition, the liver of CCl4-intoxicated rats and treated with camel milk exhibited clear hepatic recovery characterized by a complete regeneration of hepatocytes and the hepatic tissue appeared more or less normal in most cases.
Effect of treatments on hepatic lipid peroxidation: Results in Table 3 shows that the TBARS level was significantly (p<0.05) increased in the liver of Ccl4 intoxicated rats (33.3±1.01 μM) compare to the control (28.3±0.93 μM).
| Table 2: | Effect of administration of CCl4 and/or camel milk for five weeks on serum biomarker enzymes activities of AST (U L-1) and ALT (U L-1) |
 | |
| ALT: Alanine aminotransferase, AST: Aspartate amino transferase. Values are expressed as Mean±SE, n = 6 for each group; Significance was calculated at p<0.05. *Significant as compared to control animals. **significant as compared to CCl4 treated animals | |
| Table 3: | Effect of administration of CCl4 and/or camel milk for five weeks on levels of TBARS (μM) and reduced glutathione (μM) and activities of CAT (nmol min-1 g-1 tissue), SOD (U g-1 tissue), GPX (nmol min-1 g-1 tissue) and GST (nmol min-1 g-1 tissue) in rats liver tissue |
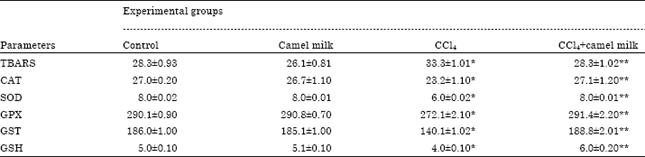 | |
| TBARS: Thiobarbituric acid reactive substance, CAT: Catalase, SOD: Superoxide dismutase, GPX: Glutathione peroxidase, GST: Glutathione-S transferase and GSH: Reduced glutathione. Values are expressed as Mean±SE, n = 6 for each group. Significance was calculated at p<0.05, *significant as compared to control animals, **significant as compared to CCl4 treated animals | |
Treatment of CCl4 intoxicated rats with camel milk caused significant (p<0.05) decrease in TBARS level (28.3±1.02 μM) of liver tissue compared to that in liver from untreated CCl4 intoxicated rats (33.3±1.01 μM). Administration of camel milk only did not affect significantly (p>0.05) the level of TABARS in rat liver compare to the control.
Effect of treatments on GSH and hepatic antioxidant enzyme activities: GSH concentration and antioxidant enzyme activities (CAT, SOD, GPX and GST) are shown in Table 3. The concentration of GSH was reduced significantly (p<0.05) in CCl4 intoxicated rats (4.0±0.10 μM) compare to the control (5.0±0.10 μM). However, camel milk administration reserves GSH values in CCl4 intoxicated rats (6.0±0.20 μM) compare to untreated CCl4 intoxicated rats (4.0±0.10 μM) and control (5.0±0.10 μM). All the antioxidant enzyme activities were reduced significantly (p<0.05) in CCl4 intoxicated rats (CAT: 23.2±1.10 nmol min-1 g-1 tissue; SOD: 6.0±0.02 U g-1 tissue; GPX: 272.1±2.10 nmol min-1 g-1 tissueand GST: 140.1±1.02 nmol min-1 g-1 tissue) compare to the control (27.0±0.20; 8.0±0.02; 290.1±0.90; 186.0±1.00), respectively. However all activities of these enzymes showed a significant (p<0.05) recovery in response to camel milk administration to CCl4 intoxicated rats (27.1±1.20; 8.0±0.01; 291.4±2.20; 188.8±2.01), respectively compare to untreated CCl4 intoxicated rats (23.2±1.10; 6.0±0.02; 272.1±2.10; 140.1±1.02), respectively. The activities of hepatic antioxidant enzymes of rats treated only with camel milk were comparable to that of the control (Table 3).
Effect of treatments on gene expression of antioxidant enzymes: The mRNA expression of SOD, CAT, GPX and GST were measured by real time PCR (Fig. 1). The results showed that, the expression of all of the antioxidant enzymes were reduced significantly (p<0.05) in CCl4 intoxicated rats compare to the control. However, the expression of all of the antioxidant enzymes were increased significantly (p<0.05) in CCl4 intoxicated rats treated with camel milk compare to untreated CCl4 intoxicated rats. The expression of hepatic antioxidant enzymes of rats treated only with camel milk was comparable to that of the control (Fig. 1).
DISCUSSION
In the current study serum liver biochemical markers, AST and ALT activities were increased significantly (p<0.05) in rats treated with CCl4 compare to control. This increase in serum levels of liver function biomarkers might be attributed to the liver damage as these enzymes are cytoplasmic in location and its released into circulation indicated damage to cell membrane (Zimmerman et al., 1965; Brent and Rumack, 1993; Recknagel et al., 1989). Similar results (Trible et al., 1987; Wang et al., 1997; Mehmetcik et al., 2008; Arici and Cetin, 2011; Althnaian et al., 2013) demonstrated that, ALT and AST enzymes activities were significantly increased following CCl4 administration. The hepatotoxic effectof CCl4 is due to active metabolite Ccl3 which has the ability to eliminate hydrogen from fatty acids and initiating the lipid peroxidation, hepatocytes injury and liver damage (Forni et al., 1983; Park et al., 2005). However, in the current study, camel milk was found to ameliorate (p<0.05) the leakage of AST and ALT induced by CCl4 treatment in rats. This suggests a membrane stabilizing activity of camel milk because it has been accepted that, serum levels of transaminases might be return to normal whenever healing of hepatic parenchyma and hepatocytes regeneration have been achieved (Thabrew et al., 1987). Supporting to the present findings, several reports (Khan and Alzohairy, 2011; Hamad et al., 2011; Al-Fartosi et al., 2012; Althnaian et al., 2013) demonstrated the hepatoprotective effect of camel milk.
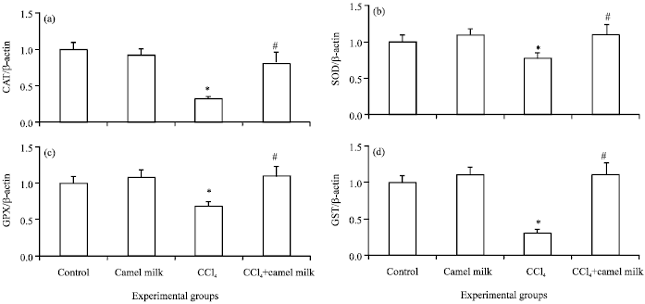 | |
| Fig. 1(a-d): | Real time PCR analysis of mRNA gene expression of hepatic catalase, (a) CAT, (b) Superoxide dismutase, SOD, (c) Glutathione peroxidase, GPX and (d) Glutathione-S transferase, GST of control non-treated, camel milk treated, CCl4 treated and CCl4+camel milk treated rats. Liver tissues were homogenized in TriZol reagent and RNA was extracted by chloroform. After preparation of cDNA, real time RT-PCR was performed using QuantiFastTM SYBR Green PCR Master Mix kit. The relative gene expression of target genes in comparison to the β-actin reference gene was calculated using the Bio-Rad CFX Manager 3.0 Software of the C1000 Touch thermal cycler-CFX96 Real time PCR. Values are expressed as mean±SEM. *Values are significantly different (p<0.5) compare to control. #Values are significantly different (p<0.5) compare to CCl4 intoxicated group |
These studies elucidated that, the ameliorative effect of camel milk against CCl4-induced oxidative stress in rats was attributed to the antioxidant effect of camel milk contents such as vitamins (A, B2, C and E) and trace elements (Yousef, 2004). The efficient hepatoprotective drugs can be defined as, drug which able to reduce the harmful effect or restore the normal hepatic function that has been distributed by toxin. Camel milk reduced the levels of liver enzymes that has been elevated as results of CCl4 toxicity. This indicated that, the structural integrity of hepatocytes cell membrane has been protected and/or damaged hepatocytes have been regenerated (Palanivel et al., 2008).
The significant increase of TBARS in liver of untreated CCl4 intoxicated rats suggests enhanced peroxidation leading to tissue damage and the failure of the antioxidant mechanisms in preventing of excessive free radicals (Romero-Alvira and Roche, 1996). This confirmed by the results of the current study which reported a significant decrease of both GSH content and the activities of antioxidant enzymes (SOD, GPX, CAT, GST) in CCl4 intoxicated rats compare to control which impose in elevation of TBARS levels. These findings were found to be in agreement with other previous studies in rats liver intoxicated with CCl4 (Yousef, 2004; Aranda et al., 2010; Al-Fartosi et al., 2012; Bona et al., 2012; Pirinccioglu et al., 2012). The significant decrease of TBARS in liver of rats intoxicated with CCl4 and treated with camel milk suggests the protective effect of camel milk against CCl4-induced oxidative stress. This protective effect of camel milk is due to contents of essential vitamins and minerals (Barbagallo et al., 1999).
GSH plays an important role in antioxidant system of the body. It maintains normal structure and function of the cells via redox and detoxification reaction. In the present study a significant decrease of GSH values were observed in CCl4 intoxicated rats. The decrease of GSH is perhaps due to direst requisition of GSH by GPX to scavenge free radicals that formed during the metabolism of CCl4. Similar results were reported in rats intoxicated with CCl4 (Bona et al., 2012). The ameliorative effect of camel milk against CCl4 induced toxic effect on GSH level was attributed to the ability of camel milk to scavenge free radicals and restore the antioxidant status which underlined by increasing the activities and gene expression of antioxidant enzymes as discussed below. Camel milk restored the GSH values in rat liver intoxicated with aluminum chloride (Al-Hashem, 2009), cadmium chloride (Al-Hashem et al., 2009) and alcohol (Darwish et al., 2012).
The oxidative stress refers to the imbalance between prooxidants and antioxidants in biological systems. Therefore, the significant increase in lipid peroxidation could be due to significant reduction in the activities of enzymatic antioxidants such as CAT, SOD, GPX and GST as well as non- enzymatic antioxidants such as GSH in the liver of CCl4-intoxicated rats, as compared to the control. Superoxide radical is converted to H2O2 by SOD. Furthermore, H2O2 is transferred to molecular oxygen and water by CAT and GPX. Therefore, SOD, CAT, GPX and GST constitute the principal components of the antioxidant system and their deficiencies leads to oxidative stress. Therefore, the significant reduction in these enzyme activities in liver of CCl4 intoxicated rats (Table 3) could be responsible for increased lipid peroxidation observed as reflected on high level of TBARS along with low level of GSH in these animals compared to the control during CCl4-induced hepatotoxicity. Significant reductions in GPX, SOD and CAT have been reported in CCl4-intoxicated rat liver (Palanivel et al., 2008; Bigoniya and Rana, 2010), kidney (Adewole et al., 2007; Ganie et al., 2011) and lung (Ganie et al., 2011). Previous study has reported that camel milk is a potent inducer of these detoxifying enzymes and thereby prevents the toxicity induced by aluminum chloride (Al-Hashem, 2009), cadmium chloride (Al-Hashem et al., 2009; Dallak, 2009) and alcohol (Darwish et al., 2012).
At gene expression level, the current findings indicated that, CCl4 induced down regulation of all examined antioxidant enzymes (SOD, CAT, GPX and GST) gene expression. Similar results (Chen et al., 2013) was obtained in CCl4-induced liver fibrosis in mice. The present findings also reported that, camel milk augmented the antioxidant status via up-regulation of CAT, SOD, GPX and GST gene expression (Fig. 1). Similar results (Afifi, 2010) demonstrated that camel milk regulated gene expression of these antioxidant enzymes in lung tissues of rats intoxicated with cisplain. The author studied gene expression of antioxidant enzymes in lung tissues of rats by conventional PCR. The conjugation of reactive xenobiotic metabolites with GSH as mentioned above is an important step in detoxification mechanism. This conjugation process is mediated by GST. CCl4 depleted the GSH through conjugation process (Table 3). The down regulation of GST gene expression in CCl4 intoxicated rats compare to control as presented in the current study (Fig. 1c) could limit the ability of hepatic tissues to conjugate the reactive metabolites which alleviated by Camel milk.
CONCLUSION
Camel milk may exert its protective effect against CCl4 induced hepatic toxicity by modulating the extent of lipid peroxidation and augmenting antioxidant defense system at activity and gene expression levels.
REFERENCES
- Adewole, S.O., A.A. Salako, O.W. Doherty and T. Naicker, 2007. Effect of melatonin on carbon tetrachloride-induced kidney injury in Wistar rats. Afr. J. Biomed. Res., 10: 153-164.
CrossRefDirect Link - Al-Fartosi, K.G., A. Majid, M.A. Auda and M.H. Hussein, 2012. The role of camel's milk against some oxidant-antioxidant markers of male rats treated with CCl4. Int. J. Res. Pharmaceut. Biomed. Sci., 3: 385-389.
Direct Link - Al-Fartosi, K.G., O.S. Khuon and H.I. Al-Tae, 2011. Protective role of camel's milk against paracetamol induced hepatotoxicity in male rats. Int. J. Res. Pharmaceut. Biomed. Sci., 2: 1795-1799.
Direct Link - Al-Hashem, F., 2009. Camel milk protects against aluminium chloride-induced toxicity in the liver and kidney of white albino rats. Am. J. Biochem. Biotechnol., 5: 98-108.
Direct Link - Aranda, M., C.D. Albendea, F. Lostale, L. Lopez-Pingarron and L. Fuentes-Broto et al., 2010. In vivo hepatic oxidative stress because of carbon tetrachloride toxicity: Protection by melatonin and pinoline. J. Pineal Res., 49: 78-85.
CrossRefPubMedDirect Link - Arici, O.F. and N. Cetin, 2011. Protective role of ghrelin against carbon tetrachloride (CCl4)-induced coagulation disturbances in rats. Regul. Pept., 166: 139-142.
CrossRefPubMedDirect Link - Barbagallo, M., L.J. Dominguez, M.R. Tagliamonte, L.M. Resnick and G. Paolisso, 1999. Effects of vitamin E and glutathione on glucose metabolism: Role of magnesium. Hypertension, 34: 1002-1006.
CrossRefPubMedDirect Link - Bigoniya, P. and A.C. Rana, 2010. Protective effect of Wrightia tinctoria bark triterpenoidal fraction on carbon tetrachloride-induced acute rat liver toxicity. Iran. J. Pharmacol. Therapeut., 9: 55-62.
Direct Link - Bona, S., L.I. Filippin, F.C. Di Naso, C. de David and B. Valiatti et al., 2012. Effect of antioxidant treatment on fibrogenesis in rats with carbon tetrachloride-induced cirrhosis. ISRN Gastroenterol.
CrossRefDirect Link - Brent, J.A. and B.H. Rumack, 1993. Role of free radicals in toxic hepatic injury II. Are free radicals the cause of toxin-induced liver injury? J. Toxicol.: Clin. Toxicol., 31: 173-196.
CrossRefPubMedDirect Link - Canbay, A., A. Feldstein, E. Baskin-Bey, S.F. Bronk and G.J. Gores, 2004. The caspase inhibitor IDN-6556 attenuates hepatic injury and fibrosis in the bile duct ligated mouse. J. Pharm. Exp. Ther., 3: 1191-1196.
PubMed - Chen, S., L. Zou, L. Li and T. Wu, 2013. The protective effect of Glycyrrhetinic acid on carbon tetrachloride-induced chronic liver fibrosis in mice via upregulation of Nrf2. PLoS ONE, Vol. 8.
CrossRef - Dallak, M., 2009. Camel's milk protects against cadmium chloride-induced hypochromic microcytic anemia and oxidative stress in red blood cells of white albino rats. Am. J. Pharmacol. Toxicol., 4: 136-143.
Direct Link - Darwish, H.A., N.R. Abd Raboh and A. Mahdy, 2012. Camel's milk alleviates alcohol-induced liver injury in rats. Food Chem. Toxicol., 50: 1377-1383.
CrossRefPubMedDirect Link - El Agamy, E.S.I., R. Ruppanner, A. Ismail, C.P. Champagne and R. Assaf, 1992. Antibacterial and antiviral activity of camel milk protective proteins. J. Dairy Res., 59: 169-175.
CrossRefDirect Link - Ganie, S.A., E. Haq, A. Hamid, Y. Qurishi and Z. Mahmood et al., 2011. Carbon tetrachloride induced kidney and lung tissue damages and antioxidant activities of the aqueous rhizome extract of Podophyllum hexandrum. BMC Complement Altern. Med., Vol. 11.
CrossRefDirect Link - Hamad, E.M., E.A. Abdel-Rahim and E.A. Romeih, 2011. Beneficial effect of camel milk on liver and kidneys function in diabetic sprague-dawley rats. Int. J. Dairy Sci., 6: 190-197.
CrossRefDirect Link - Khan, A.A. and M.A. Alzohairy, 2011. Hepatoprotective effects of camel milk against CCl4-induced hepatotoxicity in rats. Asian J. Biochem., 6: 171-180.
CrossRefDirect Link - Korhonen, H. and A. Pihlanto, 2003. Food-derived bioactive peptides: Opportunities for designing future foods. Curr. Pharm. Des., 9: 1297-1308.
PubMedDirect Link - Lee, C.H., S.W. Park, Y.S. Kim, S.S. Kang, J.A. Kim, S.H. Lee and S.M. Lee, 2007. Protective mechanism of glycyrrhizin on acute liver injury induced by carbon tetrachloride in mice. Biol. Pharmaceut. Bull., 30: 1898-1904.
CrossRefPubMedDirect Link - Omer, R.H. and A.H. Eltinay, 2009. Changes in chemical composition of Camel's raw milk during storage. Pak. J. Nutr., 8: 607-610.
CrossRefDirect Link - Palanivel, M.G., B. Rajkapoor, R. Senthil Kumar, J.W. Einstein and E.P. Kumar et al., 2008. Hepatoprotective and antioxidant effect of Pisonia aculeata L. against CCl4-induced hepatic damage in rats. Scientia Pharmaceutica, 76: 203-215.
CrossRefDirect Link - Park, W.H., S.K. Lee and C.H. Kim, 2005. A Korean herbal medicine, Panax notoginseng, prevents liver fibrosis and hepatic microvascular dysfunction in rats. Life Sci., 76: 1675-1690.
CrossRefPubMedDirect Link - Pirinccioglu, M., G. Kizil, M. Kizil, G. Ozdemir, Z. Kanay and M.A. Ketani, 2012. Protective effect of Okuzgozu (Vitis vinifera L. cv.) grape juice against carbon tetrachloride induced oxidative stress in rats. Food Funct., 3: 668-673.
CrossRefDirect Link - Recknagel, R.O., E.A. Glende, Jr., J.A. Dolak and R.L. Waller, 1989. Mechanisms of carbon tetrachloride toxicity. Pharmacol. Ther., 43: 139-154.
CrossRefPubMedDirect Link - Romero-Alvira, D. and E. Roche, 1996. High blood pressure, oxygen radicals and antioxidants: Etiological relationships. Med. Hyphothesis, 46: 414-420.
CrossRefPubMedDirect Link - Saile, B. and G. Ramadori, 2007. Inflammation, damage repair and liver fibrosis-role of cytokines and different cell types. Z. Gastroenterol., 45: 77-86.
PubMed - Stoyanovsky, D.A. and A.I. Cederbaum, 1999. Metabolism of carbon tetrachloride to trichloromethyl radical: An ESR and HPLC-EC study. Chem. Res. Toxicol., 12: 730-736.
CrossRefPubMedDirect Link - Thabrew, M.I., P.D.T.M. Joice and W. Rajatissa, 1987. A comparative study of the efficacy of Pavetta indica and Osbeckia octandra in the treatment of liver dysfunction. Planta Medica, 53: 239-241.
CrossRefPubMedDirect Link - Trible, D.L., T.Y. Aw and D.P. Jones, 1987. The pathophysiological significance of lipid peroxidation in oxidative cell injury. Hepatology, 7: 377-386.
Direct Link - Wang, P.Y., T. Kaneko, H. Tsukada, M. Nakano, T. Nakajima and A. Sato, 1997. Time courses of hepatic injuries induced by chloroform and by carbon tetrachloride: Comparison of biochemical and histopathological changes. Arch. Toxicol., 71: 638-645.
PubMed - Yagil, R., A. Saran and Z. Etzion, 1984. Camel's milk: For drinking only? Comp. Biochem. Physiol., 78: 363-366.
Direct Link - Yousef, M.I., 2004. Aluminum-induced changes in hemato-biochemical parameters, lipid peroxidation and enzyme activities of male rabbits: Protective role of ascorbic acid. Toxicology, 199: 47-57.
CrossRefDirect Link - Zimmerman, H.J., Y. Kodera and M. West, 1965. Effects of carbon tetrachloride poisoning on the plasma levels of cytoplasmic and mitochondrial enzymes in animals with nutritional fatty metamorphosis. J. Lab. Clin. Med., 66: 324-333.
PubMedDirect Link - Ramadori, G. and B. Saile, 2004. Portal tract fibrogenesis in the liver. Lab. Invest., 84: 153-159.
PubMed - Recknagel, R.O., 1983. Carbon tetrachloride hepatotoxicity: Status quo and future prospects. Trends Pharmacol. Sci., 4: 129-131.
CrossRefDirect Link - Althnaian, T., I., Albokhadaim and S.M. El-Bahr, 2013. Biochemical and histopathological study in rats intoxicated with carbontetrachloride and treated with camel milk. Springer Plus, Vol. 2.
CrossRefDirect Link