
K. Sahayaraj
Department of Advanced Zoology and Biotechnology, Crop Protection Research Centre, St. Xavier`s College (Autonomous), Palayamkottai-627002, Tamil Nadu, India
J. Shoba
Department of Advanced Zoology and Biotechnology, Crop Protection Research Centre, St. Xavier`s College (Autonomous), Palayamkottai-627002, Tamil Nadu, India
Asian Journal of Biochemistry
Year: 2012 | Volume: 7 | Issue: 3 | Page No.: 112-122







ABSTRACT
Botanical insecticides have been recommended by most of the Biointensive Integrated Pest Management (BIPM) Practitioners throughout the world. However, the target organs for such botanicals are imperative. Here, the toxicological effects of Tephrosia purpurea (Linn.) (Fabaceae) and Acalypha indica (Linn.) (Euphorbiaceae) crude extract on the mortality, reproductive organs macromolecules, mineral level in alimentary canal and detoxication enzyme level in the fat body and intestine of Dysdercus cingulatus (Fab.) (Pyrrhocoridae) was studied. Crude extract of A. indica caused more mortality than T. purpurea. Cotton leaves and seed were altered the impact of these botanicals pesticidal property. For instance, the LC50 value was higher when the pest was fed with aqueous extract treated cotton seeds than on cotton leaves. Detoxication enzymes like Aspartate Aminotransferase (AAT) and Alanine Aminotransferase (ALT) level was high in intestine than the fat body whether the insect fed with cotton leaves or cotton seeds. AAT level was enhanced by the T. purpurea extract than A. indica extract. The Na+ level high in cotton seed fed pest whereas K+ level and Ca++ level were high in cotton leaves fed animals. A. indica sprayed cotton seed induce the Na+ and K+ level in the alimentary canal. Ca++ level decreased by A. indica and T. purpurea extracts sprayed cotton seed and leaves fed pest. Treatment of T. purpurea and A. indica altered both total glucose and protein level of D. cingulatus ovary and tests. It seems that the detrimental effect of the botanical crude extract can be due to both the modulating nature of the extracts and the biochemical alteration in the respective system, alimentary canal and fat body of the D. cingulatus can be a potent candidate in integrated pest management programmes for controlling such pests.
PDF Abstract XML References Citation
Received: February 26, 2012;
Accepted: March 21, 2012;
Published: May 19, 2012
How to cite this article
K. Sahayaraj and J. Shoba, 2012. Toxic Effect of Tephrosia purpurea (Linn.) and Acalypha indica (Linn.) Aqueous Extracts Impact on the Mortality, Macromolecules, Intestinal Electrolytes and Detoxication Enzymes of Dysdercus cingulatus (Fab.). Asian Journal of Biochemistry, 7: 112-122.
DOI: 10.3923/ajb.2012.112.122
URL: https://scialert.net/abstract/?doi=ajb.2012.112.122
DOI: 10.3923/ajb.2012.112.122
URL: https://scialert.net/abstract/?doi=ajb.2012.112.122
INTRODUCTION
Due to intensive use of insecticides in arthropod control, many pest populations have developed resistance to these compounds and residual effects, respectively, resulting in worldwide efforts of finding alternative insecticides from plants or plant constituents. The botanicals were reported to be safer and were useful against crop pests by many authors. Botanical biopesticides are eco-friendly pesticides which are stored in plants as secondary metabolites. For the past 30 years, plant extracts and their bioactive molecules have been intensively utilized as an alternative pest management component because they are safe to our health and also to out environment (Seffrin et al., 2010). Available sources of indigenous plant material can possibly be used to control plant pests under field condition after screening them in the laboratory. Most of the novel bioactive principles of plants constituted by secondary metabolites like alkaloses, terpenoids, flavaonoids, phenolic compounds, organic acids and lipids (Harborne, 1998). Many of the defensive components of plants are biodegradable with non-residual effects on the biological environment; hence, an attempt has been made in the present investigation to identify plants with potential to control plant pests.
Aqueous or crude extracts or oil of number of plants have been used for the management of economically important pests world-wide (Haque et al., 2002; Rahman et al., 2003; Leatemia and Isman, 2004a, b; Bobadilla et al., 2005; Khalequzzaman and Sultana, 2006; Deepa and Bai, 2010; Ebadollahi, 2011; Silva et al., 2012). In particular, aqueous suspension of roots and leaves of Tephrosia purpurea (Del.) Pers. (Leguminosae) reduced the egg laying capacity of the insects (Ghei, 2001; Srivastava and Gupta, 2007). It also has nematicidal (Kimenju et al., 2008) activity. Anti-mosquito (Daniel et al., 1995; Govindarajan et al., 2008; Nazar et al., 2009; Sakthivadivel and Daniel, 2008) and molluscicidal activity (Sharma et al., 2009) of Acalypha indica (Linn.) = (A. ciliate Wall) (Euphorbiaceae) have already been reported.
Red cotton bug, Dysdercus cingulatus (Fab.) (Hemiptera: Pyrrhocoridae) has piercing and sucking type of mouthpart and it causes damages to leaves, seeds, developing fruits and flower bud and also transmitting fungi into the leaves, immature lint and seeds of cotton (Ahmed and Schaefler, 1987; Yasuda, 1992). Dysdercus cingulatus nymphs and adults infest cotton, lady’s finger, sambhal, hollyhock etc. and it is difficult to manage, because of its high mobility and having many alternative host plants. Previously, it was reported that extracts of Catharanthus roseus G. Don, Parthenium hysterophorus Don and Nephrolepis exaltata Schot caused morphological change in D. cingulatus (Rajendran and Gopalan, 1980). Impact of neem extracts (Gawande and Burkhade, 1989) and neem-based commercial pesticide on molting and vitellogenesis of D. cingulatus was recorded (Abraham and Ambika, 1979; Pandey and Tiwari, 2011). Ovicidal and ovipositional effects of Pedalium murex Linn on Dysdercus spp. (Gahukar, 1995) and D. cingulatus (Sahayaraj et al., 2006) was available in the literature. Very recently Dhembare and Gholap (2010) screened insecticidal activity of 13 plants against this pest.
Poor farmers prefer to spray aqueous extracts of the locally available plants rather to use high coast bioactive principles of the same plant available in the market. Both Tephrosia purpurea and Acalypha indica are endemic and even present in the road side. Detoxication enzymes play critical roles in insect physiology: inactivating ingested foreign toxin materials and save the animal for growth and development. Hence, it is imperative to know how crude extracts causing mortality and what are the biochemical changes takes place in the intestine while an insect feed botanicals treated plant parts. Enormous publications were available about the impact of plant extracts against both defoliators and sucking pests (Kotkar et al., 2002; Akhtar and Isman, 2004; Leatemia and Isman, 2004a, b; Deepa and Bai, 2010; Ebadollahi, 2011; Silva et al., 2012). However, very limited research has studies the actual mechanism by which the plant extracts cause mortality or kill the insects. We provided new information about the biochemical pathway modulated by two indigenous plants, T. purpurea and A. indica against a cosmopolitan insect D. cingulatus for the first time. In additional how detoxication enzymes involved in the insect exposed to crude plant extract has been discussed. Nothing is known about the toxicity and physiology of several of botanical insecticides to D. cingulatus. Kept the existing lacuna in the mind, the objective of the present investigation was to evaluate the impact of T. purpura and A. indica aqueous extracts on D. cingulatus adults mortality, total glucose and protein level in the reproductive systems; sodium, potassium and calcium levels in alimentary canal and detoxication enzymes (Aspartate Aminotransferase-AAT and Alanine Aminotransferase-ALT) activities in whole alimentary canal and fat body of D. cingulatus.
MATERIALS AND METHODS
Collection and extraction of plant materials: Whole plants (except root) of T. purpurea and A. indica were collected during November 2006 from St. Xavier's College campus, Palayamkottai, Tirunelveli District, Tamil Nadu, India. The collected plant materials identified using standard volumes of Flora of the presidency of Madras and Flora of the Tamil Nadu Carnatic and confirmed with the help of experts. The collected aerial plant materials were washed, shade dried for two weeks. One kilograms of areal plant material was powdered using an electric blender and 200 g powder was soaked in 500 mL boiled water for overnight with occasional shaking. Then the content was filtered using Whatman No.1 filter paper and the filtrate was concentrated using a rotary vacuum evaporator at 40°C until further use. All experiments were carried out in Crop Protection Research Centre from November 2006 to October 2007.
Preparation of stock solutions: Thirty grams of the crude extract was taken in a standard measuring flask and dissolved in 100 mL of water. To this mixture, 0.1 mL of Tween 80 (0.05%) added as an emulsifier. This mixture was made up to 500 mL using distilled water to prepare the stock solution which was considered to be a 6% solution. From the stock solution, 250 mL was placed in a beaker and used as 6% test solution and the remaining solution was diluted with water up to 500 mL and was considered as 3% solution. This sequential method was used to prepare the other following test concentrations, 1.5, 0.75 and 0.385%. A mixture of 0.1 mL of Tween 80 was made up to 100 mL in a standard measuring flask by adding distilled water to serve as the control solution. The cotton seeds (50 g) and leaves (30 g) were separately soaked in 100 mL crude extracts of the plants separately over night, then shade dried for 5-10 min and provided to the D. cingulatus as food.
Establishment of insect culture: Dysdercus cingulatus nymphs were collected from cotton fields in and around Palayamkottai, India. Dysdercus cingulatus nymphs were fed with water soaked cotton seeds in the laboratory at 28±2°C, 11±1 h photoperiod and 65-70% RH. Adults were released into the oviposition chambers for egg laying and provided with cotton leaves along with cotton seeds. Eggs were collected, kept separately and newly hatched nymphs were maintained on water soaked cotton seeds. Freshly molted adult were used for the experiments.
Toxicological bioassay: Experiments were carried out in plastic vials (400 mL capacity). Wet filter paper (6 cm2) was placed in each plastic vials to avoid early drying of the cotton seeds (variety SVPR-2). For experimental category 10 cotton seeds dipped in different concentrations of the plant extract for over night, shade dried for 5 min and kept inside the vials. A single adult D. cingulatus (starved for 6 h prior to exposure to diet) was introduced individually. In another experiment, a fresh leaf disc (4 cm diam) of cotton (variety SVPR-2) was used for D. cingulatus adults. For toxicological bioassay, the leaf discs dipped in crude extracts at 6, 3, 1.5, 0.75 and 0.385% concentrations, were screened. Leaf discs or cotton seeds treated with water were used as a negative control. Ten replicates were maintained for each treatment with five nymphs per replicate. The experiment was conducted under laboratory conditions as mentioned above. We assessed adult mortality of red cotton bug one, two, three and four days after exposure to the plant extract with counting dead specimen. The results were adjusted with Abbott's formula for corrected mortality and calculated the LC50 (Finney, 1971). After 96 h, live D. cingulatus were collected (3 and 6% plant extracts treated insects only) and used for the biochemical analyses.
Preparation of gut, fat body and reproductive system samples: Since the LC30-LC30 values were<2%, we chosen 2% for the further experiment. In another experiment, 2% plant extract was prepared, treated to cotton leaves and seeds and provided to the adult male and female (10 each) red cotton bug continuously for four days. Then, the alive insects were used for the estimation of enzymes, minerals and macromolecule. Individuals were anaesthetized with cotton soaked in chloroform and the entire reproductive tract (male and female), fat body and digestive systems were dissected out in ice cold insect Ringer’s solution (NaCl2-6.5 g; KCl2-0.25 g; CaCl2-0.25 g; Na2CO3-0.25 g in 1000 mL distilled water). The malpighian tubules, adhering tissue and gut contents were removed carefully from the alimentary canal.
Enzyme estimation: Enzyme extracts of fat body and whole alimentary canal were prepared by the method of Applebaum (1964) and Applebaum et al. (1961). The gut and fat body weighed and body part was homogenized for 3 min at 4°C in ice cold citrate-phosphate buffer (pH 6.8) (0.1 M Citric acid and 0.1 M Sodium citrate) using a tissue homogenizer with a Teflon pestle. Homogenized gut sections were suspended in buffer and diluted into 4 mL. The homogenate was centrifuged at 8000 rpm for 15 min and supernatant was used as enzyme source. The Aspartate (AAT) and Alanine Aminotransferase (ALAT) were quantified using L-aspartic acid and L-Alanine used as standard, respectively (Bergmeyer and Bernt, 1965).
Cation estimation: Cation equanimity was estimated using emission flame photometry (Pye SP90A spectrophotometer). Gut sample of 5-10 μL (depending on the availability) was diluted immediately after collection in 2 mL of distilled water and measured against 100 μM standards. The interference of sodium on the potassium value was calculated (Dow, 1981). Calcium content was estimated calorimetrically using standard procedure (Verma et al., 1957).
Total glucose and protein analysis: Reproductive organs extracts were used for the total glucose and protein quantification. The glucose was determined by the glucose oxidase method (Harold, 1988). Total protein content was estimated using bovine serum albumin as standard (Lowry et al., 1951).
Statistical analysis: The results were expressed in Mean±SE. Individual value of control categories of both seed and leaves fed D. cingulatus separately were compared with their respective feed treated with A. indica and T. purpurea separately by student ‘t’ test using SPSS (version 13) and the results were expressed at p<0.01 to p<0.05.
RESULTS
Adult mortality was significantly influenced by day after treatment (p<0.001), substance (p<0.01) and their concentration (p<0.05). Significantly the highest mortality was caused by A. indica treated cotton leaves fed group (77.4±0.5%), than cotton seeds fed group (49.5±1.9%). Significantly the lowest mortality caused T. purpurea (40.4±2.1%). We determined significantly the lowest mortality first day after treatment (41.7±2.2%) and the highest mortality third day after treatment (71.3±1.7%). The LC50 value was low for A. indica extract than for T. purpurea extract provided by cotton leaves and seeds. Both extracts of these plants exhibited strong dose dependent insecticidal activity.
Mineral elements: The Na+, K+ and Ca++ levels recorded from intestine of the pest and presented in Table 2. Na+ and K+ and Ca++ level was higher while the pest fed with cotton seed and leaves respectively. Na+ level significantly (p<0.05) decreased and increased (p<0.05) by A. indica and T. purpurea extracts treated cotton seed. A. indica extract treated cotton leaves highly reduced (p<0.01) K+ and Ca++ level (Table 2).
Glucose and protein in Reproductive system: Feeding T. purpurea extract treated cotton leaves increased the glucose (p<0.005) and protein (p<0.05) level in the ovary and testes than cotton seed fed category (Table 3). Glucose level was high while D. cingulatus fed with cotton leaves treated with A. indica extract. Ovary protein and glucose level did not changed by A. indica treated cotton leaves or seed fed insect. However, A. indica treated cotton seed and leaf fed insect testes glucose and protein level increased (Table 3). D. cingulatus ovary total glucose and protein level decreased by A. indica and T. purpurea extracts treated cotton seed and leaves.
| Table 1: | Toxic effect of Tephrosia purpurea and Acalypha indica aqueous extracts on LC50 parameters of adult Dysdercus cingulatus after 96 h of treatment |
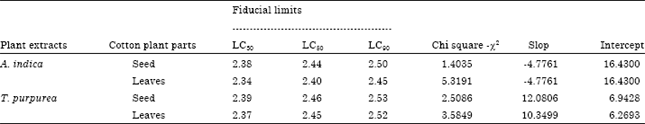 | |
| Table 2: | Impact of plant aqueous extracts on the intestinal electrolytes levels of (μg mg-1) of D. cingulatus |
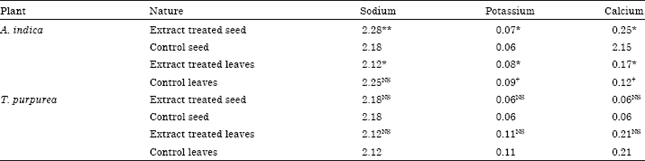 | |
| *: Significance (p<0.05), **(p<0.005) between control with experimental insects, +: Significance (5%) between control leaves with experimental insects, NS: Insignificant at 5% level | |
| Table 3: | Impact of plant extracts on total glucose and protein level of reproductive system of (μg mg-1) of D. cingulatus |
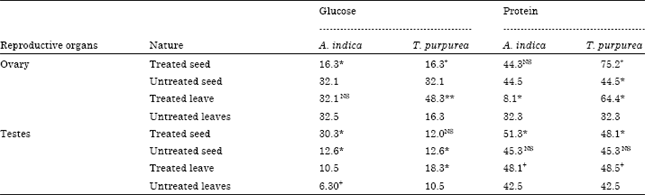 | |
| *: Significance (p<0.05), **(p<0.005) between control with experimental insects, +: Significance (5%) between control leaves with experimental insects and NS: Insignificant at 5% level | |
| Table 4: | Impact of Tephrosia purpurea and Acalypha indica aqueous extracts on the detoxication enzyme level (μg mg-1) of D. cingulatus fat body and intestine |
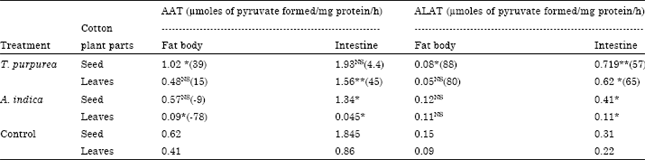 | |
| Statistical analysis between control and plant extracts treated pests (seeds and leaves separately), significance are expressed at p<0.05* and p<0.005**, specific activities as nmoles of substrate conjugated min-1 mg-1 protein | |
Fat body and alimentary canal detoxication enzymes: In control category, Aspartate (AAT) and Alanine Aminotransferase (ALAT) levels high in intestine (p<0.005) and fat body (p<0.05) of D. cingulatus, respectively. Application of T. purpurea increased AAT level both in fat body and intestine. T. purpurea extract treated cotton seed increased and decreased ALAT level in intestine and fat body, respectively. Whereas, while cotton seed treated with A. indica provided to the pest, AAT level increased in intestine and fat body (Table 4).
DISCUSSION
Present results shows that A. indica and T. purpurea crude extracts exhibited oral toxicity to the red cotton bug. Results also reveals that crude extracts caused does dependent and time-dependent mortality to the pest. The small variations in LC50 values of both insecticidal plants with respect to time of exposure and type of food by which the extracts incorporated suggest a rapid toxic action. Almost immediately following the feeding of T. purpurea and A. indica treated cotton leaves and seeds, insect movement decreased (or) spasmodic movements and feeding practically ceased or decreased, faecal elimination, thus confirming the acute toxicity of these plants to the red cotton bug. In general, the results showed the effectiveness of tested plants as insecticidal plants against the red cotton bug and effect may be due to joint action of different toxic principles in mixture that influenced different sites of action physiologically and biochemical resulting in quicker mortality. Canavanine showed significant action on the fertility of Dysdercus koenigii (Koul, 1984). Ovicidal and ovipositional effect of Pedalium murex Linn on D. cingulatus was also studied by Sahayaraj et al. (2006).
This study is the first in a series aiming to record the mineral elements level in the red cotton bug in relation to crude plant extracts ingestion through seed or leaves of the cotton plant. Mineral elements are found in most plant-feeding insects because they are constituents of plants. Calcium ion plays an important role in the regulation of muscle contraction, cell motility and activity of the nervous system. Sodium, potassium and calcium level was high in cotton seed and leaves respectively fed pest reveals that part of the plant determine the level of mineral elements in insects. In T. purpurea extracts treated cotton seed fed insect, the sodium level significantly blacked, at the same time while the extract incorporated in to the cotton leaves, sodium level slightly increased (p<0.05), because the bioactive of this plant selectively block the sodium channels in the slow inactivated state. An opposite trend was observed for A. indica extracts. Type of feed such as cotton seed and leaves were also influence the electrolyte level of D. cingulatus. For instance, both Na+, Cl¯ level was higher while D. cingulatus fed with cotton seed. Similarly K+ and Ca2+ level was higher when D. cingulatus fed with cotton leaves. But incorporation of A. indica and T. purpurea bioactive principles further reduced these electrolytes. It shows that tested plants interfere with the physiology of D. cingulatus.
Between the two plant extracts, T. purpurea showed high impact on the glucose and protein level of D. cingulatus. Earlier reports were showed that neem products inhibit the reproductive system (Koul, 1984) and vitellogenesis (Abraham and Ambika, 1979) of Dysdercus spp.. Thus was also several impaired embryogenesis will damage to trophocytes in D. koenigii (Koul, 1984) reduction in yolk material affecting choriogenesis and irregular shape of oocytes leading to complete atrophy of various and blockage of oogenesis. In the present study both T. purpurea and A. indica reduce protein content in female reproductive system evident for the reduction of yolk material in oocytes. Later it was reported that vapor of Acorus calamus Linn. reduced the fecundity of D. cingulatus. It is further believed that evident increased in the biochemical concentration in D. cingulatus is an index for the xenobiotic stress caused by both T. purpurea and A. indica. Glucose is stored in a polymeric form, glycogen that can be readily degraded on demand to be used as a glycolytic fuel (Steele, 1982). It appears that the various secondary metabolites of both T. purpurea and A. indica responsible for the physiological stresses are instrumental in reducing the biochemical constituents and metabolic disturbances. High protein and glycogen in the plant extracts treated D. cingulatus reproductive organs could be attributed to the presence of water-soluble phenolic compounds in cotton leaves treated leaves and seeds which make the plants less attractive to pests and interfere with their reproduction. Impact of neem derived pesticides i.e., neem gold on moulting and vitellogenesis of D. cingulatus were observed (Sahayaraj et al., 2006). Sahayaraj and Agnul (2004) also reported that crude plant extracts increase the whole body glycogen content of S. litura. In this study, in order to tolerate the impact of T. purpurea and A. indica toxicants, D. cingulatus increased the glycogen and protein level in the reproductive systems, because adults spent most of their energy for reproductive behaviour and oviposition.
The higher levels of free amino acids observed in the present investigation (Table 1) support this assumption. This indicates the crucial role for aminotransferases in protein synthesis in red cotton bug. The role of amino transferases in manipulation of various metabolic events like glucogenesis, gluconeogenesis, biological oxidations, histolysis and histogenesis (Reddy et al., 1992) by elevating the levels of amino transferases needs to be ascertained. The insect fat body is an organ analogous which carries out a variety of different metabolic activities comparable to mammalian liver. It is the place of intense biosynthetic activity throughout the insect life and is the main source for the hemolymph proteins.
The presence of Aspartate (AAT) and Alanine (ALAT) aminotransferase are formerly called as Glutamic Oxaloacetic Transaminase (GOT) and Glutamic pyruvate Transaminase (GPT). The presence of Aspartate (AAT) and Alanine (ALAT) aminotransferase activity was detected in haemolymph, silkgland, muscle and fat body of silkworm as reported in earlier investigations (Reddy et al., 1992). Plant extracts caused an elevation in the activity levels of both AAT and ALAT in intestine and fat body of the red cotton bug (Table 2), indicating the increased turnover of amino acids and glutamate formation during metamorphosis in red cotton bug. The most common detoxication enzyme systems are esterases, glutathione S-transferases and P450 monooxygenases (Caballero et al., 2008). Yerasi and Chitra (2000) studied the impact of annonain on Aspartate and Alanine Aminotransferase levels of Spodoptera litura. Here again, GSTs play an important role in detoxification, as has been shown for the detoxification of glucosinolates (Wheeler et al., 2001) or furocoumarins (Yu, 1996; Arora et al., 2008). However, in control categories, SGOT level was well pronounced in intestine, whereas SGPT level pronounced in the fat body of D. cingulatus adults. However, when the insect feed with A. indica and T. purpurea extracts treated cotton seeds, the enzyme level have increased indicates animal secretes more amount of enzyme to nullify the impact of the secondary metabolites of the plants. In contrast, cotton leaves treatment, in addition to increasing the enzyme level, A. indica reduces the level of detoxication enzymes (Table 2). Studies of Sahayaraj and Antony (2006) revealed that the tested five plants reduced both GST and GPT activities of S. litura indicating that the insect is susceptible to these plant products and GPT is one among the important detoxification enzymes. Further, they proposed that GST play a key role in phase II of enzymic detoxication whereas, GPT is not involved in the detoxication process in S. litura. As observed in the intestine, both the GOT and GPT levels was reduced from control category to T. purpurea treated cotton leaves and seeds fed D. cingulatus. In general, D. cingulatus showed more level of these enzymes. Similarly in A. indica treated cotton leaves and seed fed D. cingulatus fat body, both GOT and GPT. Furthermore, the results strongly suggest the inter conversion between glycogen and trehalose in plant extract treatment.
CONCLUSION
Plants offer a wide array of bioactive compounds for use as insecticides of these compounds can use local labour and simple technology and if these metabolites are sufficiently toxic, as ecological sound and it should be possible to develop culturally acceptable and inexpensive insecticides. We conclude that A. indica and T. purpurea can be used as a potent insecticide since the concentrations (LC90) used to kill the pest is around 2.5%. Application of these botanicals by foliar spray (leaf dip method) caused more mortality than soil application (seed dip method). In addition to the insecticidal activity, tested plants also showed to altered intestinal electrolytes, energy source (glucose and total protein) of reproductive organs and elevate detoxication enzyme levels to overcome the impact of the plant extracts of D. cingulatus. It is imperative to isolate and identify the biomolecules responsible for these activities and incorporate them in IPM. The best prospect for biopesticides is to use them in organized IPM programs that combine other pest control tactics in a manner consistent with local needs and constraints. These plants are widely distributed and easy grown. Furthermore, the extraction method is simple and cost-effective and the application techniques could be relatively easily designed for on-farm use.
ACKNOWLEDGMENTS
We are grateful to anonymous reviewers provided helpful comments on a previous version of this manuscript. KSR is very grateful to the Ministry of Earth Science, Govt. of India (Ref. No. 14/23/2008-ERS/RE) for the financial assistant.
REFERENCES
- Ahmed, I. and C.W. Schaefler, 1987. Food plant and feeding biology of the Pyrrhocoroidea (Hemiptera). Phytophaga, 1: 75-92.
Direct Link - Akhtar, Y. and M.B. Isman, 2004. Comparative growth inhibitory and antifeedant effects of plant extracts and pure allelochemicals on four phytophagous insect species. J. Applied Entomol., 128: 32-38.
CrossRefDirect Link - Applebaum, S.W., 1964. The action pattern and physiological role of Tenebrio larval amylase. J. Insect Physiol., 10: 897-906.
CrossRef - Applebaum, S.W., M. Jankovic and Y. Birk, 1961. Studies on the mid gut amylase activity of Tenebrio molitor L. larvae. J. Insect Physio., 7: 100-108.
Direct Link - Arora, G.K., P.J. Rup and S.K. Sohal, 2008. Influence of coumarin on the enzymatic activity in nymphs of Lipaphis erysimi (Kalt.). Allelopathy J., 22: 221-229.
Direct Link - Deepa, M.A. and V.N. Bai, 2010. Bioinsecticidal compounds of celastraceae-the spindle tree family. Int. J. Botany, 6: 220-227.
CrossRefDirect Link - Bobadilla, M., F.M. Zavala, G.Z. Sisniegas, J. Mostacero and L. Taramona, 2005. Evaluacion larvicida de suspensiones acuosas de Annona muricata Linnaeus (guanabana) sobre Aedes aegypti Linnaeus (Diptera, Culicidae). Revista Peruana De Biologia, 12: 15-152.
Direct Link - Caballero, C.J., Lopez-Olguin1, M. Ruiz1, F. Ortego and P. Castanera1, 2008. Antifeedant activity and effects of terpenoids on detoxication enzymes of the beet armyworm, Spodoptera exigua (Hubner). Span. J. Agri. Res., 6: 177-184.
Direct Link - Dow, J.A.T., 1981. Ion and water transport in locust alimentary canal: Evidence from in vivo electrochemical gradients. J. Exp. Bio., 93: 167-179.
Direct Link - Ebadollahi, A., 2011. Iranian plant essential oils as sources of natural insecticide agents. Int. J. Biol. Chem., 53: 266-290.
CrossRefDirect Link - Govindarajan, M., A. Jebanesan, T. Pushpanathan and K. Samidurai, 2008. Studies on effect of Acalypha indica L. (Euphorbiaceae) leaf extracts on the malarial vector, Anopheles stephensi Liston (Diptera:Culicidae). Prasito. Res., 103: 691-695.
CrossRefPubMedDirect Link - Khalequzzaman, M. and S. Sultana, 2006. Insecticidal activity of Annona squamosa L. seed extracts against the red flour beetle, Tribolium castaneum (HERBST). J. Bio-Sci., 14: 107-112.
CrossRefDirect Link - Kimenju, J.W., A.M. Kagundu, J.H. Nderitu, F. Mambala, G.K. Mutua and G.M. Kariuki, 2008. Incorporation of green manure plants into bean cropping systems contribute to root-knot nematode suppression. Asian J. Plant Sci., 7: 404-408.
CrossRefDirect Link - Kotkar, H.M., P.S. Mendki, S.V. Sadan, S.R. Jha, S.M. Upasani and V.L. Maheshwari, 2002. Antimicrobial and pesticidal activity of partially purified flavonoids of Annona squamosa. Pest Manag. Sci., 58: 33-37.
CrossRefDirect Link - Koul, O., 1984. I-Interaction with the development of red cotton bugs. Entomo. Exp. Application, 36: 85-88.
CrossRefDirect Link - Leatemia, J.A. and M.B. Isman, 2004. Insecticidal activity of crude seed extracts of Annona spp., Lansium domesticum and Sandoricum koetjape against lepidopteran larvae. Phytoparasitica, 32: 30-37.
CrossRefDirect Link - Leatemia, J.A. and M.B. Isman, 2004. Toxicity and antifeedant activity of crude seed extracts of Annona squamosa (Annonaceae) against lepidopteran pests and natural enemies. Int. J. Trop. Insect Sci., 24: 150-158.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Nazar, S., S. Ravikumar, G.P. Williams, M. Syed Ali and P. Suganthi, 2009. Screening of Indian coastal plant extracts for larvicidal activity of Culex quinquefasciatus. Indian J. Sci. Technol., 2: 24-27.
Direct Link - Haque, N.M.M., M.F. Rabbi, A.N.M.R.H. Karim, Mainul and S.K. Biswas, 2002. Chemical methods of leaf extraction of bankalmi, Polygonum hydropiper for controlling rice hispa beetles, Dicladispa armiger (Olivier) (Coleoptera: Chrysomelidae) in Bangladesh. J. Biological Sci., 2: 782-784.
CrossRefDirect Link - Pandey, J.P. and R.K. Tiwari, 2011. Neem based insecticides interaction with development and fecundity of red cotton bug, Dysdercus cingulatus Fab. Int. J. Agric. Res., 6: 335-346.
CrossRefDirect Link - Rahman, M.A., M.A. Taleb and M.M. Biswas, 2003. Evaluation of botanical product as grain protectant against grain weevil, Sitophilus granarious (L.) on wheat. Asian J. Plant Sci., 2: 501-504.
CrossRefDirect Link - Rajendran, B. and M. Gopalan, 1980. Juvenile hormone like activity of certain plant extracts on Dysdercus cingulatus Fab. Indian J. Agric. Sci., 50: 781-784.
Direct Link - Sakthivadivel, M. and T. Daniel, 2008. Evaluation of certain insecticidal plants for the control of vector mosquitoes viz. Culex quinquefasciatus, Anopheles stephensi and Aedes aegypti. Applied Entomo. Zool., 43: 57-63.
CrossRefDirect Link - Seffrin, R.D.C., I. Shikano, Y. Akhtar and M.B. Isman, 2010. Effects of crude seed extracts of Annona atemoya and Annona squamosa L. against the cabbage looper, Trichoplusia ni in the laboratory and greenhouse. Crop Prot., 29: 20-24.
CrossRefDirect Link - Silva, L.B., Z.F. Xavier, C.B. Silva, O. Faccenda, A.C.S. Candido and M.T.L.P. Peres, 2012. Insecticidal effects of Croton urucurana extracts and crude resin on Dysdercus maurus (Hemiptera: Pyrrocoridae). J. Entomol., 9: 98-106.
CrossRefDirect Link - Srivastava, M. and L. Gupta, 2007. Effect of formulations of Solanum surratense (Family: Solanaceae) an Indian desert plant on oviposition by the pulse beetle Callosobruchus chinensis Linn. Afr. J. Agric. Res., 2: 552-554.
Direct Link - Verma, M.R., V.M. Bhuchar and K.J. Therattil, 1957. Direct chelatometric method for the estimation of calcium or magnesium in the presence of phosphate. Nature, 179: 1244-1244.
CrossRefDirect Link - Yerasi, B. and K.C. Chitra, 2000. Response of aspartate and alanine aminotransferase to sublethal concentration of annonain by Spodoptera litura (fab.). Indian J. Entomol., 62: 367-370.
Direct Link