
P.C. Chikezie
Department of Biochemistry, Imo State University, Owerri, Imo State, Nigeria
LiveDNA: 234.10425
ORCID: 0000-0001-5066-724X
Asian Journal of Biochemistry
Year: 2011 | Volume: 6 | Issue: 1 | Page No.: 98-103







ABSTRACT
The aim of the present study was to ascertain methaemoglobin concentrations and levels of NADH-methaemoglobin reductase activity of three human erythrocyte genotypes, namely, HbAA, HbAS and HbSS. The cyanomethaemoglobin reaction was used for the determination of erythrocyte haemolysate methaemoglobin concentration. NADH-methaemoglobin reductase activity was measured by the rate of oxidation of NADH + H+ when, the erythrocyte enzyme was incubated in potassium ferrocyanide {K3Fe(CN)6}. Whereas, methaemoglobin concentrations in the three erythrocyte genotypes was in the range of 1.45±0.13 to 2.50±0.43%, in the order: HbAS<HbAA<HbSS, levels of NADH-methaemoglobin reductase activity ranged between 8.86±2.49 and 14.77±1.47 IU gHb-1, in the order of: HbSS<HbAS<HbAA. However, there was no significant difference (p<0.05) in methaemoglobin concentration and NADH-methaemoglobin reductase activity between HbAA and HbAS erythrocytes. The results showed a relationship between erythrocyte NADH-methaemoglobin reductase activity and methaemoglobin concentration.
PDF Abstract XML References Citation
How to cite this article
P.C. Chikezie, 2011. Methaemoglobin Content and NADH-methaemoglobin Reductase Activity of Three Human Erythrocyte Genotypes. Asian Journal of Biochemistry, 6: 98-103.
DOI: 10.3923/ajb.2011.98.103
URL: https://scialert.net/abstract/?doi=ajb.2011.98.103
DOI: 10.3923/ajb.2011.98.103
URL: https://scialert.net/abstract/?doi=ajb.2011.98.103
INTRODUCTION
Methaemoglobin (MHb) is an altered state of haemoglobin in which the ferrous (Fe2+) irons of heme are oxidized to the ferric (Fe3+) state (Murray et al., 2006). The ferric haem moities of MHb are unable to bind oxygen molecules. NADH-Methaemoglobin Reductase (NADH-MR) (EC: 1.6.2.2) is a 300 amino acid, membrane-bound enzyme localized in the endoplasmic reticulum of all cells (Dekker et al., 2001). This enzyme transfers electrons from NADH to cytochrome b5 via its Flavin Adenine Dinucleotide (FAD) prosthetic group (Yubisui et al., 1980). Under normal conditions, this NADH-dependent enzyme also referred to as ferricyanide reductase, NADH-diaphorase or cytochrome b5 reductase is the only system within the erythrocyte that maintains hemoglobin in its oxygen-carrying reduced state (Rockwood et al., 2003). It is noteworthy that a second form of this enzyme, which is NADPH dependent, is less active in mammals in converting MHb back to hemoglobin than is the NADH-dependent form (Smith, 1991).
Deficiencies in the reductase systems and resulting clinical manifestations have been described (Jaffee, 1981). Generally, cellular activity of NADH-MR reflects an organism’s capacity to reduce MHb and is therefore an important factor in evaluating MHb formation and etiology of methaemoglobinemia. From a toxicological standpoint, NADH-MR is the rate-limiting enzyme controlling the toxicokinetics of the reduction of MHb (Rockwood et al., 2003). In addition, previous report by Borgese et al. (1987) stated that the activity of NADH-MR is generally reduced in red cells of patients with recessive hereditary methaemoglobinemia. NADH-MR activity has been measured in a variety of non-human species (Agar and Harley, 1972; Lo and Agar, 1986; Whittington et al., 1995) and these studies have shown that species with lower NADH-MR activities convert MHb back to haemoglobin slower than do species with higher activities (Smith, 1991).
This study seeks to establish whether variability in NADH-MR activity exist amongst human erythrocyte genotypes, namely; HbAA, HbAS and HbSS, in relationship with their respective methaemoglobin contents. The findings of the present investigations will provide insights into the oxidant status of the three erythrocyte genotypes and deduce the degree of tolerance to toxic methaemoglobinogenic agents by individuals who express these genotypes.
MATERIALS AND METHODS
Selection of volunteers/collection of blood samples: Forty three healthy male (61-73 kg) participants of confirmed HbAA (15), HbAS (15) and HbSS (13) genotypes between the ages of 20-28 years enrolled for this study between the months of February and May, 2009. Five milliliters of human venous blood of HbAA, HbAS and HbSS genotypes were obtained from participants by venipuncture and stored in EDTA anticoagulant tubes. Blood samples of HbSS genotype were from patients attending clinics at the Federal Medical Center (FMC), Imo State University Teaching Hospital (IMSUTH), Orlu, St. John Clinic/Medical Diagnostic Laboratories, Avigram Medical Diagnostic Laboratories and Qualitech Medical Diagnostic Laboratories. These centers are located in Owerri, Imo State, Nigeria.
Ethics: The institutional review board of the Department of Biochemistry, University of Port Harcourt, Port Harcourt, Nigeria, granted approval for this study and all participants involved signed an informed consent form. This conducted study was in accordance with the ethical principles that have their origins in the Declaration of Helsinki. Individuals drawn were from Imo State University, Owerri, Nigeria and environs. The research protocols were in collaboration with registered and specialized clinics and medical laboratories.
Preparation of erythrocyte haemolysate: The erythrocytes were washed by methods as described by Tsakiris et al. (2005) and Chikezie (2009). Within 2 h of collection of blood samples, portions of 1.0 mL of the samples were introduced into centrifuge test tubes containing 3.0 mL of buffer solution pH = 7.4: 250 mM tris-HCl (Tris-HCl)/140 mM NaCl/1.0 mM MgCl2/10 mM glucose). The erythrocytes were separated from plasma by centrifugation at 1200x g for 10 min, washed three times by three similar centrifugations with the buffer solution. The erythrocytes re-suspended in 1.0 mL of this buffer were stored at 4°C. The washed erythrocytes were lysed by freezing/thawing as described by Galbraith and Watts (1980) and Kamber et al. (1984). The erythrocyte haemolysate was used for the determination of NADH-methaemoglobin reductase activity and methaemoglobin concentration.
Determination of methaemoglobin concentration of erythrocyte lysate: Determination of methaemoglobin content of red cell lysate was by modification of the method of Evelyn and Malloy (1938), as described by Akomopong et al. (2000) and Chikezie (2009). A total of 400 μL of 0.5 M Phosphate buffer (pH 6.5) was added to 600 μL of the cell lysate and the mixture was centrifuged at 16,000x g for 5 min to sediment debris. A total of 700 μL of the supernatant fraction was used to measure the absorbance at λmax = 630 nm (the absorbance maximum for methaemoglobin) and the reading was recorded as SI. A total of 50 μL of 10 g% KCN was added and after 5 min at room temperature (24°C), a second reading (S2) was recorded. KCN converts methaemoglobin to cyanomethaemoglobin, which does not absorb at 630 nm; hence, the difference between absorbance readings S1 and S2 represents the absorbance due to methaemoglobin.
To measure total hemoglobin levels, all of the hemoglobin was converted to methaemoglobin, the absorbance of the sample at λmax = 630 nm was recorded and then KCN was added to form cyanomethaemoglobin. Specifically, 70 μL of the supernatant fraction was diluted 10-fold into 600 μL of 0.1 M phosphate buffer (pH = 6.5). Next, 30 μL of freshly prepared 20 g% K3Fe(CN)6 was added and incubated for 5 min at room temperature (24°C) and an initial reading (T1) was recorded. A total of 50 μL of 10% KCN was subsequently added and a second reading (T2) was recorded. The percent methaemoglobin in the sample was calculated as:
[100(S1-S2)]/[10(T1-T2)] |
Determination of erythrocytes haemolysate haemoglobin concentration: A modified method (Baure, 1980) described by Chikezie et al. (2009), based on cyanomethaemoglobin reaction was used for the determination of haemolysate haemoglobin concentration. The expressed values were in g dL-1. A 0.05 mL portion of human red blood cell haemolysate was added to 4.95 mL of Drabkin reagent (100 mg NaCN and 300 mg K4Fe(CN)6 L-1). The mixture was left to stand for 10 min at room temperature and absorbance read at λmax = 540 nm against a blank. The absorbance was used to evaluate for haemolysate haemoglobin concentration by comparing the values with the standards.
Determination of nadh-methaemoglobin reductase activity of erythrocyte lysate: The enzyme activity represents the rate of oxidation of NADH and is described as follows:
NADH-MR activity of erythrocyte lysate was assayed according to the method of Board et al. (1990). A mixture of 0.2 mL tris-HCl/EDTA buffer pH = 8.0, 0.2 mL NADH and 4.35 mL of distilled water was introduced into a test tube and incubated for 10 min at 30°C. The whole content was transferred into a cuvette and the reaction above started by adding 0.2 mL of K3Fe (CN)6/0.05 mL erythrocyte haemolysate. The increase in absorbance of the medium was followed spectrophotometrically at λmax = 340 nm min-1 for 10 min at 30°C against a blank solution.
Calculation of Enzyme Activity: The expression below was used to evaluate erythrocyte NADH-MR activity in international unit per gram haemoglobin (IU gHb-1):
Where:
| EA | = | Enzyme activity in IU gHb-1 |
| [Hb] | = | Haemolysate haemoglobin concentration (g dL-1) |
| O.D min-1 | = | Change min-1 in absorbance at 340 nm |
| Σ | = | Millimolar extinction coefficient = 6.22, in reaction in which 1 mole of NADH + H+ is oxidized |
| VC | = | Cuvette volume (total assay volume) = 1.0 mL |
| VH | = | Volume of haemolysate in the reaction system (0.05 mL) |
Statistical analysis: The experiments were designed in a completely randomized method and data collected were analyzed by the analysis of variance procedure while treatment means were separated by the Least Significance Difference (LSD) incorporated in the Statistical Analysis System (SAS) package of 9.1 version.
RESULTS AND DISCUSSION
The Mean±SD methaemoglobin concentration, expressed as percentage (Met.Hb%) of total haemoglobin concentration of three erythrocyte genotypes (HbAA, HbAS and HbSS) was in the order: HbAS<HbAA<HbSS. Whereas, there was no significant difference (p>0.05) in Met.Hb% between HbAA and HbAS erythrocytes, level of methaemoglobin in sickle cell genotype exhibited significantly higher (p<0.05) concentration compared with the HbAA and HbAS genotypes (Table 1).
The three human erythrocyte genotypes showed varied levels of NADH-MR activity and were in the order: HbSS<HbAS<HbAA. Similar to methaemoglobin concentrations, there was no significant difference (p>0.05) in NADH-MR activity between HbAA and HbAS erythrocytes (Table 1). Therefore, there was an inverse relationship between Met.Hb% and NADH-MR activity amongst the three human erythrocytes genotypes.
Previous reports have associated the dysfunctional erythrocytes (HbSS genotype) with raised level of oxidized haemoglobin (Van Kiujk et al., 1987; Chikezie, 2009). They noted that the primary reason for the relatively raised concentration of oxidized haemoglobin (methaemoglobin) in HbSS erythrocytes was higher production of superoxide ion by these erythrocytes compared to those of HbAA and HbAS erythrocytes. Specifically, sickle erythrocytes spontaneously generate approximately twice as much superoxide, peroxide and hydroxyl radical when compared with normal erythrocytes. In addition, HbSS erythrocytes contain increased amount of malondialdehyde, a by-product of lipid peroxidation and evidence of abnormal amino group cross-linking by malondialdehyde has been demonstrated in lipid extract of HbSS erythrocytes membranes (Das and Nair, 1980). This phenomenon confirms the susceptibilty of sickle erythrocytes to endogenous free radical-mediated oxidative damage that correlates with the proportion of irreversible sickled erythrocytes (Rice-Evans et al., 1986).
The relative lower activity of NADH-MR of sickle erythrocytes compared with the HbAA and HbAS erythrocytes suggest an inverse relationship between the methaemoglobin concentration of human erythrocytes and NADH-MR activity. In concord with this finding, previous reports by Agar and Harley (1972), Lo and Agar (1986) and Whittington et al. (1995) showed that the capacity of non human species to revert oxidized haemoglobin-Fe3+ (MHb) to the reduced form haemoglobin-Fe2+ is inextricable connected with the NADH-MR activity of the corresponding erythrocytes. In addition, Geetha et al. (2007) averred that high level of methaemoglobin concentration in cirrhotic erythrocytes might be partly due to low level of methaemoglobin reduction engendered by low level of NADH-MR activity.
| Table 1: | Methaemoglobin concentration (Met.Hb%) and NADH-methaemoglobin reductase activity of male erythrocyte haemolysate |
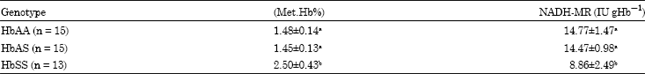 | |
| Means±SD in the column with the same letter are not significantly different at p<0.05 according to LSD. n = No. of blood samples | |
Therefore, this study suggests that persons who express the sickle erythrocytes will exhibit more susceptibilty and slower rate of recovery when challenged with agents that causes toxic methaemoglobinemia. This suggestion agrees with the findings of Daly et al. (1983), who reported that several members of a family who showed decreased activities of erythrocyte NADH-cytochrome b5 reductase, predisposed them to the development of clinically significant methaemoglobinaemia when administered with oxidant drugs.
REFERENCES
- Akomopong, T., N. Ghori and K. Haldar, 2000. In vitro activity of riboflavin against the human malaria parasite Plasmodium falciparum. Antimicrobial Agents Chemothera, 44: 88-96.
Direct Link - Board, P., M. Coggan, P. Johnston, V. Ross, T. Suzuki and G. Webb, 1990. Genetic heterogeneity of the human glutathione transferases; a complex of gene families. Pharmacol. Thera., 48: 357-369.
PubMed - Borgese, N., G. Pietrini and S. Gaetani, 1987. Concentration of NADH-cytochrome b5 reductase in erythrocytes of normal and methemoglobinemic individuals measured with a quantitative radioimmunoblotting assay. J. Clin. Invest., 80: 1296-1302.
Direct Link - Chikezie, P.C., 2009. Comparative methaemoglobin concentrations of three erythrocyte genotypes (HbAA, HbAS and HbSS) of male participants administered with five antimalarial drugs. Afr. J. Biochem. Res., 3: 266-271.
Direct Link - Chikezie, P.C., A.A. Uwakwe and C.C. Monago, 2009. Glutathione S-transferase activity of erythrocyte genotypes HbAA, HbAS and HbSS in male volunteers administered with fansidar and quinine. Afr. J. Biochem. Res., 3: 210-214.
Direct Link - Daly, J.S., D.E. Hultquist and D.L. Rucknagel, 1983. Phenazopyridine induced methaemoglobinaemia associated with decreased activity of erythrocyte cytochrome b5 reductase. J. Med. Genet., 20: 307-309.
CrossRefDirect Link - Das, S.K. and R.C. Nair, 1980. Superoxide dismutase, gluthatione peroxidase, catalase and lipid peroxidation of normal and sickled erythrocytes. Br. J. Haematol., 44: 87-92.
CrossRef - Dekker, J., M.H. Eppink, R. van Zwieten, T. Rijk and A.F. Remacha et al., 2001. Seven new mutations in the nicotinamide adenine dinucleotide reduced-cytochrome b5 reductase gene leading to methemoglobinemia type I. Blood, 97: 1106-1114.
Direct Link - Galbraith, D.A. and D.C. Watts, 1980. Changes in some cytoplasmic enzymes from red cells fractionated into age groups by centrifugation in Ficoll/Triosil gradients. Comparison of normal humans and patients with Duchenne muscular dystrophy. Biochem. J., 191: 63-70.
Direct Link - Geetha, A., M.D. Lakshmi Priya, S. Annie Jeyachristy and R. Surendran, 2007. Level of oxidative stress in the red blood cells of patients with liver cirrhosis. Indian J. Med. Res., 126: 204-210.
PubMed - Kamher, E., A. Poyiagi and G. Deliconstantinos, 1984. Modifications in the activities of membrane-bound enzymes during in vivo ageing of human and rabbit erythrocytes. Comp. Biochem. Physiol. B: Comp. Biochem., 77: 95-99.
CrossRefDirect Link - Lo, S.C.L. and N.S. Agar, 1986. NADH-methemoglobin reductase activity in the erythrocytes of newborn and adult mammals. Experientia, 42: 1264-1265.
CrossRef - Rice-Evans, C., S.C. Omorphos and E. Baysal, 1986. Sickle cell membranes and oxidative damage. Biochem. J., 237: 265-269.
CrossRefPubMedDirect Link - Rockwood, G.A., K.R. Armstrong and S.I. Baskin, 2003. Species comparison of methemoglobin reductase. Exp. Biol. Med., 228: 79-83.
PubMedDirect Link - Tsakiris, S., A. Giannoulia-Karantana, I. Simintzi and K.H. Schulpis, 2005. The effect of aspartame metabolites on human erythrocyte membrane acetylcholinesterase activity. Pharmacol. Res., 53: 1-5.
CrossRefPubMedDirect Link - Van Kuijk, F.J.G.M., A. Sevanian, G.J. Handelman and E.A. Dratz, 1987. A new role of phospholipase A2: Protection of membranes from lipid peroxidation damage. Trends Biochem. Sci., 12: 31-34.
CrossRef - Whittington, A.T., A.L. Parkinson, P.B.S. Spencer, G. Grigg and L. Hinds et al., 1995. Comparative study of the antioxidant defence systems in the erythrocytes of Australian marsupials and monotremes. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol., 110: 267-272.
PubMed - Yubisui, T., M. Takeshita and Y. Yoneyama, 1980. Reduction of methaemoglobin through flavin at the physiological concentration by NADPH-flavin reductase of human erythrocytes. J. Biochem., 87: 1715-1720.
Direct Link