
D. Kalpana Deepa Priya
Department of Medical Biochemistry, Dr. ALM Post Graduate Institute of Basic Medical Sciences, University of Madras, Sekkizhar Campus, Chennai-600 113, India
R. Gayathri
Department of Medical Biochemistry, Dr. ALM Post Graduate Institute of Basic Medical Sciences, University of Madras, Sekkizhar Campus, Chennai-600 113, India
G.R. Gunassekaran
Department of Medical Biochemistry, Dr. ALM Post Graduate Institute of Basic Medical Sciences, University of Madras, Sekkizhar Campus, Chennai-600 113, India
S. Murugan
Department of Medical Biochemistry, Dr. ALM Post Graduate Institute of Basic Medical Sciences, University of Madras, Sekkizhar Campus, Chennai-600 113, India
D. Sakthisekaran
Department of Medical Biochemistry, Dr. ALM Post Graduate Institute of Basic Medical Sciences, University of Madras, Sekkizhar Campus, Chennai-600 113, India
Asian Journal of Biochemistry
Year: 2011 | Volume: 6 | Issue: 5 | Page No.: 395-405







ABSTRACT
Sulforaphane (SFN) is an organosulfur compound found in brassica vegetables. The present study was undertaken to investigate the defending role of sulforaphane at a dose of 9 μmoles/mouse/day against the cellular dysfunction in benzo(a)pyrene [B(a)P] (100 mg kg-1 b.w., i.p.) induced experimental lung carcinogenesis. The sub cellular derangements were assessed by the cytotoxic and cellular functional markers, lysosomal enzyme leakage and enzymes of nucleic acid metabolism. The concept of free radicals contributing to carcinogenesis was confirmed by evaluating thioredoxin reductase and heme oxygenase. The carcinogenic biomarkers were complemented by immunohistochemical analysis of cell proliferating nuclear antigen-Ki67 and measurement of serum carcinoembryonic antigen by ELISA. Our work proves the chemoprotective potential of sulforaphane in vivo against cellular derangements caused by B(a)P in the process of carcinogenesis.
PDF Abstract XML References Citation
Received: February 11, 2011;
Accepted: May 16, 2011;
Published: July 29, 2011
How to cite this article
D. Kalpana Deepa Priya, R. Gayathri, G.R. Gunassekaran, S. Murugan and D. Sakthisekaran, 2011. Inhibitory Effect of Sulforaphane against Benzo(a)pyrene Induced Lung Cancer by Modulation of Biochemical Signatures in Female Swiss Albino Mice. Asian Journal of Biochemistry, 6: 395-405.
DOI: 10.3923/ajb.2011.395.405
URL: https://scialert.net/abstract/?doi=ajb.2011.395.405
DOI: 10.3923/ajb.2011.395.405
URL: https://scialert.net/abstract/?doi=ajb.2011.395.405
INTRODUCTION
Recently women have been diagnosed in increasing numbers than men of Lung cancer which is often caused by exposure to cigarette smoke or other environmental carcinogens. Of the many substances present in tobacco smoke, B(a)P is considered a prototype polycyclic aromatic hydrocarbon (PAH), classic DNA damaging agent and carcinogen. It is a ubiquitous pollutant found in amounts of 10 ng per cigarette contributing about 200 ng/day for a pack-a-day smoker (Scherer et al., 2000). The process of tumorigenesis is initiated when a replication-competent cell acquires a mutation in a ‘gatekeeping’ pathway that endows it with a selective growth advantage. Many of the known gatekeepers were identified through the study of unusual families with predispositions to specific types of cancers (Knudson, 2002). It appears that the cells of solid tumors must accumulate several rate-limiting mutations in cancer genes to achieve malignant status. Each of these cells has multiple genetic abnormalities and is capable of rapidly evolving variants to combat any therapeutic onslaught (Nygren and Larsson, 2003).
Recent focus of cancer chemoprevention is on intermediate biomarkers capable of detecting early changes that can be correlated with inhibition of carcinogenic progression. Although Sulforaphane (SFN) is not a direct-acting antioxidant, there is substantial evidence that SFN acts indirectly to increase the antioxidant capacity of animal cells and their abilities to cope with oxidative stress (Dinkova-Kostova et al., 2006). It is highly effective in preventing or reducing tumor formation induced by carcinogens in animal models (Fahey et al., 2002). Much of sulforaphane’s cytoprotective enzyme induction is thought to occur via the actions of the transcription factor Nrf2 (Fahey et al., 2002).
A potential strategy for diagnosing lung cancer, the leading cause of cancer-related death, is to identify metabolic signatures (biomarkers) of the disease. Such analysis will lead to new biological insights of the disease identifying potential therapeutic targets. With this initiative in mind the present study was undertaken to analyze the protection offered by sulforaphane against B(a)P induced biochemical alterations during the process of carcinogenesis in serum and tissue samples of cancer induced and treated female Swiss Albino Mice (SAM). This will further help us to analyze the metabolic state of the disease involving cellular and sub-cellular organelle dysfunction.
MATERIALS AND METHODS
Drugs and chemicals: Benzo(a)pyrene was purchased from Sigma Chemical Company, USA. Sulforaphane (>99% purity) was purchased from LKT Laboratories (St Paul, MN). All other chemicals used were of analytical grade obtained from Sisco Research Laboratories Pvt. Ltd., Mumbai, India and Glaxo Laboratories, CDH division, Mumbai, India.
Animals: Healthy female Swiss albino mice (4-6 weeks old) weighing 15-20 g were used throughout the study (2007-2010). Mice were acclimated to laboratory condition with regular temperature control ranging from 23±2°C and were given ad libitum access to balanced diet (Gold Mohor rat feed, Ms. Hindustan Lever Ltd., Mumbai) and water. All the experiments were performed in compliance with the regulation of our institutional Animal Care and Use Committee. They were maintained in a controlled environment condition of alternative 12 h light/dark cycles. This research work on female Swiss albino mice was sanctioned and approved by our Institutional animal ethical committee (IAEC/No-02/077/2007).
Treatment schedule: The animals were divided into five groups with six mice in each group.
| Group I: | Control animals treated with corn oil (vehicle) intraperitoneally |
| Group II: | Animals were treated with Benzo(a)pyrene [B(a)P] (100 mg kg-1 b.wt., dissolved in corn oil, intraperitoneally) thrice a week to induce lung cancer. The method of cancer induction was adopted from Wattenberg et al. (1997) and given intraperitoneally thrice a week at a dose of 100 mg kg-1 b.wt. and treatment conditions were accordingly decided in the current study |
| Group III: | Animals treated with sulforaphane (9 μmoles/mouse/day) on alternate days for two weeks prior to first dose of the carcinogen and continued until the 12th week and sacrificed (Pre-treatment group) |
| Group IV: | B(a)P administered animals were treated with sulforaphane from the 12th week to the 20th week (Post-treatment group) |
| Group V: | Animals treated with sulforaphane alone as in Group III served as drug control animals |
After the respective treatment periods the animals were sacrificed and their blood, lung and liver tissues were used for the biochemical analyses. Both the organs were excised immediately and washed in ice cold saline to remove any extraneous matter and cleaned and blotted to dryness in Whatman No. 1 filter paper. A 10% homogenate of lung and liver tissues were prepared in appropriate buffers as necessitated by the protocols. Dilutions were decided based on the protein concentration. A portion of the lung tissue was immediately stored for immunohistochemical studies in 10% buffered formalin. The method of Lowry et al. (1951) was adopted for the estimation of protein content in the serum and tissue homogenates.
Biochemical markers
Enzymes of cellular integrity and cytotoxicity: The activity of alkaline phosphatase was determined using disodium phenyl phosphate as the substrate (King, 1965a) and was expressed as Fmoles of phenol liberated/min/mg protein in tissue and IU L-1 in serum. AST and ALT were assayed by the method of King (1965b) and expressed as μmoles of pyruvate liberated/min/mg protein in tissue and IU L-1 in serum. The assay of LDH was based on its ability to convert lactate to pyruvate in the presence of the coenzyme nicotinamide adenine dinucleotide (NAD+) (King, 1965c) and expressed as μmoles of pyruvate liberated/min/mg protein in tissue and U L-1 in serum. Activity of γGT was determined by the method of Orlowski and Meister (1965) using L-γ-gutamyl-p-nitroanilide as the substrate and expressed as nmoles of p-nitroaniline liberated /min/mg protein. Acid phosphatase was assayed by the method of King (1965a) and the activity was expressed as ìmoles of phenol liberated/min/mg protein under incubation conditions.
Lysosomal markers and nucleic acid enzymes: The activity of cathepsinD was estimated by the method of Biber et al. (1981) using haemoglobin as the substrate and expressed as μmoles of tyrosine released/min/mg protein. βD-Glucuronidase activity was assayed by the method of Kawai and Anno (1971) and expressed as μmoles of p-nitrophenol liberated/min/mg protein.
5’-Nucleotidase was assayed by the method of Luly et al. (1992) and expressed as ìmoles of inorganic phosphorus liberated/min/mg protein. ADA activity was measured by the spectrophotometric method described by Guisti and Galanti (1974) and expressed as international unit (IU L-1) in serum and ìmoles of NH3 liberated/mgprotein/hr in tissue. XO was assayed by the method of Fried and Fried (1966) and expressed as U mg-1 protein (1 unit corresponds to the change of 0.1 OD/min by 1 mL of the enzyme).
Oxidative stress markers and carcinogenic markers: Thioredoxin reductase activity was assessed by the methodology adopted by Holmgren (1977) and expressed as U mg-1 protein (1unit corresponds to1 μmole of TNB formed/min). The activity of heme oxygenase was determined by the method of Tenhunen et al. (1969) and expressed as μmoles of bilirubin formed/min/mg protein based on its millimolar extinction coefficient -27.7. The activity assay of aryl hydrocarbon hydroxylase was modified from the method of Buening et al. (1981) and expressed as pmoles of phenolic metabolite formed/min/mg protein in serum. Assay of Amylase activity in serum and tissue was carried out by the method of Nelson (1944) and expressed as ìmoles of maltose liberated/mg protein in tissue and IU L-1 in serum.
Carcinoembryonic antigen (CEA): Quantitative estimation of CEA was based on the solid phase enzyme linked immunosorbent assay. CEA was assayed using UBI Magiwell enzyme immunoassay kit. The values were expressed as ng mL-1 serum.
Immunohistochemistry for Ki67: Immuno Histochemical analysis of Ki67 expression was carried out on paraffin sections of lung tissue using the B-SAP universal staining kit (Span diagnostic ltd, Surat, India). Initially, the sections were de-paraffinised in xylene and dehydrated in ethanol. After antigen retrieval the slides were incubated for 5 min with blocking solution (10% normal goat serum) at room temperature. Then the sections were incubated for 4 h with mouse monoclonal anti-Ki67 antibody (Sigma chemicals). Then washed with PBS and subsequently incubated with biotinylated secondary link antibody for 30 min at room temperature. Finally treated with Diaminobenzidine chromogen for 15 min and then washed with deionised water, counter stained with hematoxylin and mounted. Photographs were taken using Nikon microscope ECLIPSE E 400, model 115, Japan.
Statistical analysis: Data are presented as the Mean±Standard deviation (SD).One-way Analysis of Variance (ANOVA) was used to detect the significant changes between the groups. The student Least Significant Difference (LSD) method was used to compare the means of different groups and the significance was denoted by ‘P’ value. A commercial software SPSS version 10.0 was employed to find out the statistical significance between the groups and p<0.05 was considered statistically significant.
RESULTS
Figure 1a represents the effect of sulforaphane on the activities of marker enzymes such as aspartate Aminotransaminase (AST), alanine Aminotransaminase (ALT), Acid Phosphatase (ACP) and Alkaline Phosphatase (ALP) in serum of control and experimental groups. A profound increase was noticed in the activities of all serum maker enzymes signifying p<0.001; p<0.01; p<0.01; p<0.001 for AST, ALT, ACP and ALP respectively in group II animals. On treatment with sulforaphane, a significant reduction (p<0.001) in the activities of the above mentioned enzymes were observed except for ACP signifying p<0.05 in group III and IV animals.
The activities of serum Aryl Hydrocarbon Hydoxylase (AHH) and adenosine deaminase were significantly increased (p<0.001) in the group II induced animals and decreased (p<0.01; p<0.05) respectively on treatment with sulforaphane as shown in Fig. 1b. In tissue a significance of p<0.001 for ADA was observed on treatment with sulforaphane in group III and IV animals.. Interestingly amylase was found to be decreased (p<0.001) in the serum but increased (p<0.001) in tissue of cancer induced animals and increased (p<0.01 and p<0.05) in serum in group III and IV animals treated with sulforaphane. A reverse effect was seen (p<0.001) in tissue of pre-and post-treated animals. Drug control animals showed no significance when compared with control animals.
Figure 1c illustrates the alteration in the activity of carcinogenic marker (γGT) in serum, lung, liver and 5’nucleotidase activity in liver of experimental animals. In group II animals γGT was profoundly increased signifying with p<0.001 in serum and lung and with p<0.01 in liver and similar observation was made with 5’nucleotidase in liver of group II animals. Pre-treatment with sulforaphane in group III animals decreased the activity of γGT signifying (p<0.001) in the serum and lung and (p<0.01) in liver. Group IV animals showed a similar decline (p<0.05) for γGT in serum and both tissues. A considerable reduction with p<0.05 and p<0.01 for 5’ nucleotidase was observed in the group III and IV animals groups respectively. The sulforaphane alone treated animals (group V) did not show any significant changes when compared with control animals (group I).
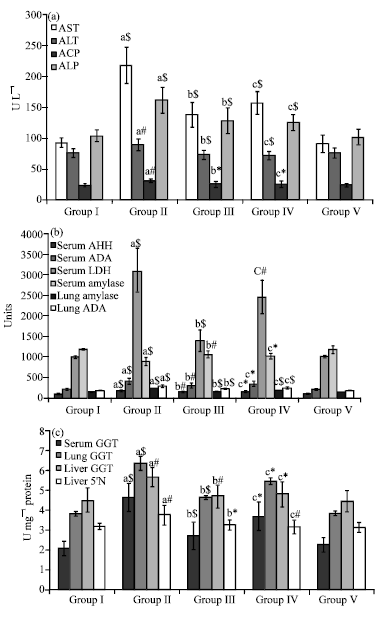 | |
| Fig. 1: | Effect of sulforaphane on the (a) serum and (b) and (c) carcinogenic biomarkers of control and experimental mice. Values are expressed as Mean±SD (n = 6); a: as compared with group I; b and c: as compared with group II; *p<0.05; #p<0.01; $p<0.001; NS = Not significant |
Figure 2a describes the modulations in the activities of liver function assessment enzymes. These include AST, ALT and ALP. In group II animals, the increased activities of ALT and AST (p<0.01) along with ALP (p<0.001), were found to be decreased significantly (p<0.05) for ALT and (p<0.001) for AST and ALP in group III and IV animals as compared to group I animals. Group V animals did not show any significant change when compared with control animals (group I).
Figure 2b illustrates the alteration in the activities of cytotoxic markers-LDH and ACP in lung and liver. LDH was profoundly increased signifying with p<0.001 in both tissues and acid phosphatase was increased with p<0.001 in lung and p<0.05 in liver in group II cancer induced animals. Their decrease was significant in the pre-treated and post-treated group signifying p<0.001 for LDH in both tissues. A reduction signifying p<0.001 on pre-treatment and p<0.01 on post-treatment for ACP in lung and p<0.05 in liver of group III and IV sulforaphane treated animals was observed was observed. The drug control group showed no significant changes.
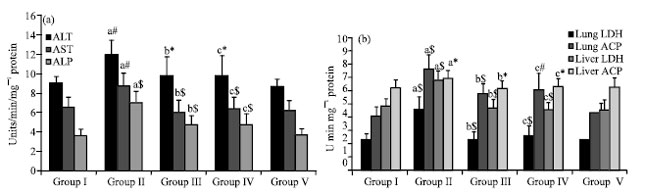 | |
| Fig. 2: | Effect of sulforaphane on the tissue (a) functional and (b) cytotoxic markers of control and experimental mice. Values are expressed as Mean±SD (n = 6); a: as compared with group I; b and c: as compared with group II; *p<0.05; #p<0.01; $p<0.001; NS = Not significant |
Figure 3a explains the effect of sulforaphane on the lysosomal markers (Cathepsin D and βD-glucuronidase) and nucleic acid enzymes, 5’Nucleotidase(5’N) and Xanthine Oxidase (XO) in Lung tissue of control and experimental animals. A significant increase in βD-glucuronidase (p<0.001) and cathepsin D (p< 0.05) was observed in group II carcinogen administered animals when compared with controls. Likewise a radical decrease (p<0.01) for βD-glucuronidase was found in group III and IV animals. A noticeable increase in the group III (p<0.05) and group IV (p<0.001) for cathepsin D was noticed. A significant increase in the activities of 5’N and XO (p<0.001) were observed in group II induced animals. On pre-treatment with sulforaphane the activities decreased (p<0.001). There was a slight difference in the alleviation in the group IV animals for these enzymes representing a significance of p<0.05 for 5’N and p<0.01 for XO. Drug control animals showed no significance when compared with control animals.
Figure 3b depicts the modulation of oxidative stress markers-Thioredoxin Reductase (TRxR) and Haem Oxygenase (HO) associated with carcinogenesis. The activities of these enzymes were found opposing in the tumor induced animals where a significant decline was noticed with TRxR (p<0.01) and increase with HO (p<0.001).A similar opposing effect in the reverse order was seen on treatment with sulforaphane in the pre-(p<0.05) and post-treated(p<0.01) group for both enzymes.
Figure 3c shows the effect of sulforaphane on the expression of the fetal antigen Carcino Embryonic Antigen (CEA) in the serum of control and experimental mice. A significant increase in the expression of CEA in serum of cancer bearing group II animals when compared with control group I animals was detected. On treatment with sulforaphane this was decreased in group III and group IV animals when compared with group II animals. No significant changes were noticed in group V animals when compared with group I control animals.
Figure 4 represents the microphotographs of lung sections of the different experimental groups showing immunoreactivity to the cell proliferating nuclear antigen-Ki67. Group II slide has 4+ degree of positive reactivity when compared to the 2+ score in the post-treated group. The pre-treatment group III and drug control group IV showed no reactivity as seen in group I control slide.
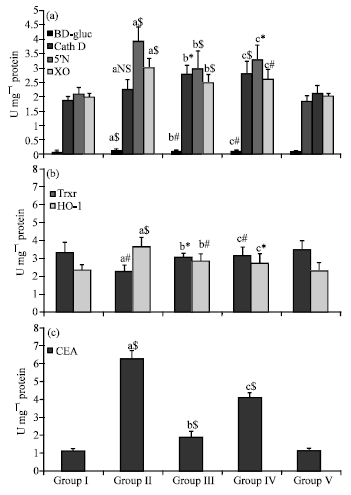 | |
| Fig. 3: | Effect of sulforaphane on (a) lysosomal enzymes, nucleic acid enzymes, (b) oxidative stress markers and (c) serum CEA levels of control and experimental animals. Values are expressed as Mean±SD (n = 6); a: as compared with group I; b and c: as compared with group II; *p<0.05; #p<0.01; $p<0.001; NS = Not significant |
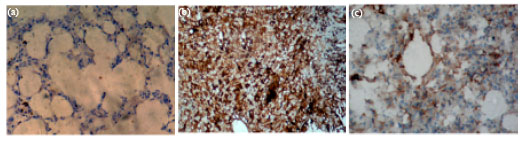 | |
| Fig. 4: | Immunoreactivity to Ki 67 in control and experimental animals. (a) Group I: Control animals showing absence of immunoreactivity for Ki67 (200X), (b) Group II: B(a)P treated animals showing a high gradation of 4+ positivity confined to the nucleus (200X) and (c) Group IV: Sulforaphane post-treated animals with highly reduced immunoreactivity graded 2+ positivity for Ki67 (200X) |
DISCUSSION
The present study reflects the biochemical response of a transformed tissue involving various cellular derangements by experimental induction of lung carcinogenesis in mice. The aspartate and alanine aminotransferases (AST and ALT) are important enzymes of liver whose activities are related with the amino acid homeostasis. Their elevation in serum has been attributed to the damaged structural integrity of hepatic cell membrane as reported earlier (Gupta et al., 2006). (Chidambara Murthy et al., 2005). In this study, we found that exposure to B(a)P brings about the hepatocellular damage due to oxidative stress leading to increased enzyme activity which is evident from a progressive increase in the serum levels of ALP, AST and ALT. AST being a key mitochondrial enzyme, its increase in the serum levels would indicate excessive mitochondrial proliferation (Hassanein, 2004). Treatment with SFN in group III and IV significantly reduced the elevated levels of the enzymes towards their respective normal values indicating rehabilitation of plasma membrane and as well as repair of hepatic tissue damage induced by B(a)P.
Establishing the activities of some DNA turnover enzymes and free radical metabolizing enzymes in cancerous tissues seems to be of particular importance to elucidate possible relations between cancer and free radical metabolisms. In several studies, ADA was found increased in cancerous tissue and cells compared to non-cancerous ones (Camici et al., 1990) and this coincides well with present results which reflects an accelerated purine turnover and high salvage pathway activity of nucleic acid metabolism associated with tissue hyper proliferation in B(a)P induced pulmonary pathogenesis. Also a high XO activity recorded in group II B(a)P induced animals is due to increased conversion of Xanthine dehydrogenase to oxidase (Sushma et al., 2006) . This may be an attempt to lower salvage pathway activity which is vital for rapid DNA synthesis in cancer cells. The treatment with SFN had therapeutic consequences in B(a)P induced tumorigenesis with a concordant decrease of the enzymes, reflecting a decreased purine turnover and hence the proliferation rate in group III and IV animals.
A significant increased activity of amylase in the tissue (Lung) might be due to its increased local production. Likely an increase in the serum was not observed in the cancer induced group. This might be due to the enzyme modification caused by the toxic metabolites formed during B(a)P activation leading to its functional impairment. The marker enzymes such as AHH, ADA, GGT, 5’-ND and LDH are specific indicators of lung damage (Tessitore et al., 1994).The increase in their activities may be due to the increased tumor incidence (Ravichandran and Ramanibai, 2008). Also damage to plasma membrane causes the leakage of these enzymes from the cytosol to the blood stream (Shanmugarajan et al., 2008). Lactate dehydrogenase is recognized as a potential tumor marker in assessing the progression of the proliferating malignant cells due to its increased turnover. Gamma glutamyl transpeptidase (γ-GT) is not only useful in diagnosis but also has prognostic value in malignancies such as lung cancer and malignant melanoma. Its increase in group II animals was to compensate the decreased GSH levels and an inhibitory effect was posed by SFN in group III and IV animals which could be due to its stimulatory effect on the GSH biosynthesis, hence reduced the depletion of GSH from the cells by a autocrine positive feed back loop mechanism.
Intracellular Acid Phosphatase (ACP) and βD-glucuronidase (βGR) are largely confined to lysosomes. Their activities correlate and primarily respond to cellular injury and inflammation (Kumar et al., 2005). Their increase in serum indicate an enhanced Golgi activity and peroxidation in lysosomal membranes after exposure to B(a)P causing membrane lysis leading to enzyme leakage. Cathepsin D, an endopeptidase was found recently to be an important regulator of apoptosis acting as a direct activator of caspase 3 and 9 (Minarowska et al., 2007). In the current observation, its mild release was noticed in the cancer induced group II animals due to increased lipid peroxidation by B(a)P. This response was further boosted upon treatment with SFN in group III and IV indicating the selectivity of the phytochemical in initiating apoptosis and further support the emerging picture of Cathepsin D as an important mediator of programmed cell death.
Thioredoxin reductases (TrxRs) play an important role in multiple cellular events such as ROS detoxification, oxidoreductase activities and cytokine effects (Yodoi, 2000) related to carcinogenesis including cell proliferation, apoptosis and cell signaling. HO-1 displays antioxidant, anti-apoptotic and anti-inflammatory effects and appears to have a complex role in angiogenesis (Prawan et al., 2005). In present study both enzymes were slightly elevated as a means of compensatory adaptation to the depleted antioxidant enzymes and especially GSH (Priya et al., 2011) in B(a)P treated mice. Increased levels of TrxR have been reported in many different tumors with a correlation to malignancy and poor prognosis (Kahlos et al., 2001). On treatment with sulforaphane, TrxR activity was still increased due to the formation of more SFN-GSH conjugate as reported earlier by Zhang and Callaway (2002). This acts as a driving force for the formation of reduced glutathione. The antioxidant action of hemeoxygenase is favored by the breakdown of heme proteins thus reducing the lipid peroxidation and the production of ROS in SFN treated animals. Carcinoembryonic antigen is a cell surface glycoprotein expressed in fetal tissues and is transcriptionally silent in adults and its elevated levels suggest that CEA could play an important role in cancer progression (Jessup et al., 1999). The observed significantly high levels of CEA in cancer bearing animals indicated the progression of tumor growth. A decrease in CEA level, upon treatment is associated with a better survival as seen in group III and IV animals. Ki-67 protein is an excellent marker for determining proliferating cells in human and animal neoplasm (Ozaki et al., 2007). In the present study, the Ki-67 labeling index increased in group II animals indicating cell proliferation as it is expressed in all phases of cell cycle except in the resting G0 and early G1 phases. It has been previously reported to be strongly associated with the percentage of growth fraction (Petrowsky et al., 2001) and poor prognosis in pulmonary adenocarcinomas. The confirmation of adenocarcinoma by B(a)P was complemented by CEA levels and degree of Ki67 positivity.
CONCLUSION
In summary, we were able to analyze different biochemical signatures in serum and tissues of mouse model of lung cancer using different biochemical methodologies. The biomarkers analyzed in the present study have their metabolic origins in various biochemical pathways which are important for the assessment of the risk involved in the process of carcinogenesis. The importance of this study is that it establishes the feasibility of using such biomarkers to detect the initiation of lung carcinogenesis which involves various cellular dysfunctions in the target and non-target tissues.
ACKNOWLEDGMENTS
One of the authors Ms. D. Kalpana Deepa Priya, wishes to gratefully acknowledge the support given by the University Grants Commission, New Delhi, in the form of UGC scholarship under the UGC XI Plan Scheme of Research Fellowship in Sciences for Meritorious Students.
REFERENCES
- Scherer, G, S. Frank, K. Riedel, I. Meger-Kossien and T. Renner, 2000. Biomonitoring of exposure to polycyclic aromatic hydrocarbons of nonoccupationally exposed persons. Cancer Epidemiol. Biomarkers Prev., 9: 373-380.
Direct Link - Dinkova-Kostova, A.T., S.N. Jenkins, J.W. Fahey, L. Ye and S.L. Wehage et al., 2006. Protection against UV-light-induced skin carcinogenesis in SKH-1 high-risk mice by sulforaphane-containing broccoli sprout extracts. Cancer Lett., 240: 243-252.
PubMed - Fahey, J.W., X. Haristoy, P.M. Dolan, T.W. Kensler and I. Scholtus et al., 2002. Sulforaphane inhibits extracellular, intracellular and antibiotic-resistant strains of Helicobacter pylori and prevents benzo[α]pyrene-induced stomach tumors. Proc. Natl. Acad. Sci. USA., 99: 7610-7615.
Direct Link - Wattenberg, L.W., T.S. Wiedmann, R.D. Estensen, C.L. Zimmerman, V.E. Steele and G.J. Kelloff, 1997. Chemoprevention of pulmonary carcinogenesis by aerosolized budesonide in female A/J mice. Cancer Res., 57: 5489-5492.
PubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Orlowski, M. and A. Meister, 1965. Isolation of gamma-glutamyl transpeptidase from hog kidney. J. Biol. Chem., 240: 338-347.
PubMedDirect Link - Biber, J., B. Stieger, W. Haase and H. Murer, 1981. A high yield preparation for rat kidney brush border membranes different behaviour of lysosomal markers. Biochim. Biophys. Acta., 647: 169-176.
CrossRef - Kawai, Y. and K. Anno, 1971. Mucopolysaccharide-degrading enzymes from the liver of the squid, Ommastrephes sloani pacificus I. Hyaluronidase. Biochim. Biophys. Acta-(BBA) Enzymol., 242: 428-436.
CrossRefDirect Link - Luly, P., O. Barnabei and E. Tria, 1972. Hormonal control in vitro of plasma membrane bound (Na+-K+)-ATPase of rat liver. Biochimica et Biophysica Acta (BBA)-Biomembranes, 282: 447-452.
CrossRefDirect Link - Holmgren, A., 1977. Bovine thioredoxin system: Purification of thioredoxin reductase from calf liver. J. Biol. Chem., 252: 4600-4606.
PubMed - Tenhunen, R., H.S. Marver and R. Schmid, 1969. Microsomal heme oxygenase. J. Biol. Chem., 244: 6388-6394.
Direct Link - Buening, M.K., R.L. Chang, M.T. Huang, J.G. Fortner, A.W. Wood and A.H. Conney 1981. Activation and inhibition of benzo(α)pyrene and aflatoxin B1 metabolism in human liver microsomes by naturally occurring flavanoids. Cancer Res., 41: 67-72.
PubMed - Nelson, N., 1944. A photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem., 153: 375-380.
CrossRefDirect Link - Gupta, A.K., H. Chitme, S.K. Dass and N. Misra, 2006. Hepatoprotective activity of Rauwolfia serpentina rhizome in paracetamol intoxicated rats. J. Pharmacol. Toxicol., 1: 82-88.
CrossRefDirect Link - Chidambara Murthy, K.N., J. Rajesha, A. Vanitha, M.M. Swamy and G.A. Ravishankar, 2005. Protective effect of Dunaliella salina. A marine micro alga, against carbon tetrachloride-induced hepatotoxicity in rats. Hepatol. Res., 33: 313-319.
PubMedDirect Link - Hassanein, T., 2004. Mitochondrial dysfunction in liver disease and organ transplantation. Mitochondrion, 4: 609-620.
CrossRefDirect Link - Sushma, N.J., U. Sivaiah, N.J. Suraj and K.J. Rao, 2006. Aluminium acetate induced oxidative stress in brain of albino mice. J. Pharmacol. Toxicol., 1: 579-584.
CrossRefDirect Link - Camici, M., M.G. Tozzi, S. Allegrin, A. Del Corso and O. Sanfilippo et al., 1990. Purine salvage enzyme activities in normal and neoplastic human tissues. Cancer Biochem. Biophys., 11: 201-209.
PubMed - Tessitore, L., P. Costelli and M.F. Baccino, 1994. Pharmacological interference with tissue hypercatabolism in tumour-bearing rats. Biochem. J., 299: 71-78.
PubMed - Ravichandran, K. and R. Ramanibai, 2008. Protective effect of flavanoidal fraction of Indigofera tinctoria on benzo (α) pyrene induced lung carcinogenicity in swiss albino mice. Int. J. Cancer Res., 4: 71-80.
CrossRefDirect Link - Shanmugarajan, T.S., M. Arunsundar, I. Somasundaram, D. Sivaraman, E. Krishnakumar and V. Ravichandran, 2008. Ameliorative effect of Ficus hispida Linn. Leaf extract on cyclophosphamide-induced oxidative hepatic injury in rats. J. Pharmacol. Toxicol., 3: 363-372.
CrossRefDirect Link - Kumar, M.H., K.S. Radha and S.C. Gajaria, 2005. Algal diets reverse diethyl nitrosamine (DEN) induced hepatocarcinoma in rats. Int. J. Cancer Res., 1: 41-46.
CrossRefDirect Link - Minarowska, A., L. Miarowski, A. Karwowska and M. Gacko, 2007. Regulatory role of cathepsin D in programmed cell death. Folia Histochem. Cytobiol., 45: 159-163.
Direct Link - Yodoi, J., 2000. Evolution of thioredoxin and redox signaling research: Viewpoint. Antioxid. Redox. Signal., 2: 629-630.
PubMed - Prawan, A., J.K. Kundu and Y.J. Surh, 2005. Molecular basis of heme oxygenase-1 induction: Implications for chemoprevention and chemoprotection. Antioxid. Redox. Signal., 7: 1688-1703.
PubMed - Priya, D.K., R. Gayathri, G. Gunassekaran, S. Murugan and D. Sakthisekaran, 2011. Chemopreventive role of sulforaphane by upholding the GSH Redox cycle in pre- and post-initiation phases of experimental lung carcinogenesis. Asian Pacific J. Cancer Prev., 12: 103-110.
PubMed - Kahlos, K., Y. Soini, M. Saily, P. Koistinen and S. Kakko et al., 2001. Up-regulation of thioredoxin and thioredoxin reductase in human malignant pleural mesothelioma. Int. J. Cancer, 95: 198-204.
PubMed - Zhang, Y. and E.C. Callaway, 2002. High cellular accumulation of sulforaphane, a dietary anticarcinogen, is followed by rapid transporter-mediated export as a glutathione conjugate. Biochem. J., 364: 301-307.
PubMed - Jessup, J.M., S. Ishii, T. Mitzol, K.H. Edmiston and Y. Shoji, 1999. Carcinoembryonic antigen facilitates experimental metastasis through a mechanism that does not involve adhesion to liver cells. Clin. Exp. Metastasis., 17: 481-488.
PubMed - Ozaki, K., T. Yamagami, K. Nomura and I. Narama, 2007. Prognostic significance of surgical margin, Ki-67 and cyclin D1 protein expression in grade II canine cutaneous mast cell tumor. J. Vet. Med. Sci., 69: 1117-1121.
PubMed - Petrowsky, H., I. Sturm, O. Graubitz, D.A. Kooby and E. Staib-Sebler et al., 2001. Relevance of Ki-67 antigen expression and K-ras mutation in colorectal liver metastases. Eur. J. Surg. Oncol., 27: 80-87.
PubMed