
Kuzhandaivel Hemalatha
K.S.R. College of Engineering, K.S.R. Kalvi Nagar, Tiruchengode-637 215, Tamil Nadu, India
Selvaraj Venkatesan
Measurements Laboratory, Gujarat Tea Processors and Packers Ltd., Vipul State, Khokhra, Ahmadabad-380008, Gujarat, India
Asian Journal of Biochemistry
Year: 2011 | Volume: 6 | Issue: 5 | Page No.: 384-394







ABSTRACT
Tea soils are enriched with more amount of Fe and the soils of tea plantations are continuously saturated with water for several weeks. This leads to depletion in oxygen, causes reduction in oxidation process and enhances the reduction process. During reduction process all the Fe3+ ions are reduced to Fe2+ which is the more favored form for plant uptake. This increase in uptake of Fe2+ ion may cause changes in the biochemical constituents and disturbs the enzyme activity. So the necessity occurs to study the impact of excess iron on biochemical parameters and enzyme activity in tea plant. The study was conducted at nursery level using one year old plant of clone UPASI 9 under green house condition. The chlorophyll and polyphenols content increased with the addition of smaller amount of iron, a sharp decline was noted with increase in iron dosage. This is proved by the negative and significant correlation coefficient obtained between soil applied Fe and leaf polyphenols (r = -0.801 p = 0.05). The activity of invertase was higher even at higher dosage whereas the amylase activity decreases with increase in iron concentration. It was found that the glutamate synthase activity and amino acid content decreases sharply with increase in iron addition. The decrease in glutamate synthase activity and amino acid content proves that excess iron influence the nitrogen metabolism leading to lack of nitrogen assimilation in tea plant which ultimately reflects on productivity. Even though iron increases the activity of enzymes like amylase, invertase and biochemical parameters to certain extent, at higher concentration it was found harmful to plants leading to plant death.
PDF Abstract XML References Citation
Received: April 29, 2011;
Accepted: June 22, 2011;
Published: July 29, 2011
How to cite this article
Kuzhandaivel Hemalatha and Selvaraj Venkatesan, 2011. Impact of Iron Toxicity on Certain Enzymes and Biochemical Parameters of Tea. Asian Journal of Biochemistry, 6: 384-394.
DOI: 10.3923/ajb.2011.384.394
URL: https://scialert.net/abstract/?doi=ajb.2011.384.394
DOI: 10.3923/ajb.2011.384.394
URL: https://scialert.net/abstract/?doi=ajb.2011.384.394
INTRODUCTION
Tea (Camellia sinensis), belongs to Theaceae family is cultivated in thirty one countries (Madhu et al., 2011). In India, tea is grown over an area of 5.78 lakh ha mainly under rain fed sloping land conditions (Madhu et al., 2011). Tea drinking is associated with the reduction of serum cholesterol, prevention of low density lipoprotein oxidation, decreased risk of cardiovascular disease and cancer (Chung et al., 2003). In tea, there are other compounds which are beneficial to human health like fluoride, caffeine and essential minerals (Cabrera et al., 2003). Iron is one of such essential mineral for plants required for biological redox system (Asad and Rafique, 2000) and an essential component of many enzymes, plays important role in the physiological and biochemical processes of plants. It acts as a co-factor of key enzymes involved in plant hormone synthesis and participates in numerous electron transfer reactions (Kerkeb and Connolly, 2006). The perennial plants like tea have to face great difference in iron availability in the environment because of their immobility and either starvation or excess of this element is believed to generate oxidative stress (Abdel-Kader, 2007) and is responsible for several nutritional disorders affecting the physiology of plants (Becker and Asch, 2005). Oxidative stress due to excessive production of Reactive Oxygen Species (ROS) viz., superoxide radical (O2¯), hydrogen peroxide (H2O2) and hydroxyl radical (OH¯) is a common consequence of most biotic and abiotic stresses (Polle and Rennenberg, 1993). The toxicity of reactive oxygen species depends on the presence of a fentons catalyst such as iron or copper ions, giving rise to extremely reactive OH¯ radical in the presence of H2O2 and O2¯ (Chen and Schopfer, 1999). The reactive toxic oxygen species causes damage to DNA, proteins, lipids, chlorophyll and almost every other organic constituent of living cell (Becana et al., 1998). Plants protect cell and sub cellular systems from the cytotoxic effects of these reactive oxygen species with antioxidant enzymes and metabolites such as carotenoids, ascorbic acid etc. (Alscher et al., 1997). Although iron is present in both Fe2+ and Fe3+ forms in soils, it is taken up by plants only in Fe2+ form. The available iron content of tea soils varies from 50 to 100 mg kg-1. Since oxygen diffuses in air 103 to 104 times faster than in water or water saturated soils, oxygen is depleted rapidly by the respiration of soil microorganisms and plant roots in waterlogged soils and also in continuously wetted soils. With the depletion in oxygen, Fe3+ can act as electron acceptor for microbial respiration and subsequently reduced to Fe2+ (Becker and Asch, 2005). The soils of tea plantations are continuously saturated with water for several weeks and therefore the danger of iron toxicity is not eliminated. Even though the toxic effect of iron is not generally reported from tea gardens, it is worthwhile to generate data on the same aspect. In the previous attempt, influence of Fe toxicity on various other nutrients was studied by Hemalatha et al. (2006). The main objective of the current study was to document the influence of iron toxicity on the biochemical parameters like chlorophyll, carotenoids, amino acids and polyphenols and the activities of enzymes like amylase, invertase, aspartate aminotransferase and glutamate synthase of tea leaves.
MATERIALS AND METHODS
Pot grown tea plants (one year old) of the clone UPASI-9 were used for this study with six treatments including an untreated control. The pot was filled with 4 kg of soil to which ferrous sulphate was added externally so as to have 50, 500, 750, 1000, 3000 and 5000 mg of Fe per kg of soil. The experiment was conducted in triplicate. The plants were monitored everyday and the visual symptoms when noticed were recorded. The experiment was continued for a period of 90 days and thereafter the plant was uprooted and separated into root, stem and leaf. The samples were analyzed for nutrients, enzyme activities and biochemical parameters.
Glutamate synthase assay: Exactly 0.5 g of plant material was homogenized with 5 mL of 100 mM phosphate buffer (pH 7.5) containing 1 mM disodium EDTA, 1 mM dithioerythrietol and 1% polyvinyl pyrolidone. It was centrifuged at 10,000 g for 30 min at 4°C. The supernatant was collected and used as enzyme source. The activity was assayed spectrophotometrically as described by VandeCasteele et al. (1975).
Amylase and invertase: The reducing sugar released was estimated by Nelson-Somogyi method (Somogyi, 1952).
Aspartate amino transferase: Aspartate amino transferase catalyses the following reaction:
The aspartate amino transferase was assayed by the method developed by Bergmeyer and Brent (1974).
Biochemical parameters: Leaf samples of known quantity weighed in a mortar was ground with sufficient quantities of chilled methanol. It was filtered and made up to 50 mL in a volumetric flask-using methanol. This solution was diluted five times with methanol and used for the estimation of chlorophyll A, B and Carotenoids (Wellburn, 1994) by measuring the absorbance at 470, 653 and 666 nm using UV-VIS spectrophotometer (GBC 918).
About 0.5 g of leaf sample was weighed and ground with ethyl alcohol. The contents were filtered and the filtrate was made up to 50 mL with ethyl alcohol. This alcoholic extract was used for the estimation of polyphenols (Dev-Choudhury and Goswami, 1983) and amino acids (Moore and Stein, 1948) and the absorbance was recorded at 700 and 570 nm using UV-VIS Spectrophotometer (GBC 918).
Statistical analysis: Statistical analysis was carried out by the standard method (Gomez and Gomez, 1984). The correlation coefficient was worked out using Special Purpose Software for Statistics (SPSS) 12.1 for windows.
RESULTS AND DISCUSSION
The plants which received 5000 mg Fe kg-1 soil, developed toxicity symptoms on 9th day after imposing treatments. The bottom most leaf showed the symptom first. Intermittent yellowing on leaf surface was noted during the initial stages in the bottom most leaf and at later stages it turned into a coppery red. This could be because the excess Fe2+ may catalyse the generation of active oxygen species such as super oxide, hydroxyl radical and H2O2 as follows:
This reaction is greatly influenced when Fe is abundant and iron itself can be a part of highly reactive per-ferryl radicals (Halliwell and Gutteridge, 1984). Free radicals are responsible for the damage caused by iron toxicity (Thengbai and Goodman, 2000). These observations are in line with the observation made by Yamanouchi and Yoshida (1981), in rice plant where the symptoms are particularly developed in older leaves due to higher transpiration rate. Similar kind of rusty brown observation was noted in rice by Fairhurst and Witt (2002). The plants established on soil containing 3000 and 1000 mg Fe kg-1 also showed toxicity symptoms on 37th and 41st day, respectively. The symptoms appeared little later on (44th and 90th) on the plants established with 750 and 500 mg Fe Kg-1. Although the lower part of leaves was showing toxicity symptoms continuously throughout the study, no casualty was noted. A mild defoliation was noted after loosing the entire green color. The upper most leaves of the tea plant established in soil containing above 500 mg Fe kg-1 showed reddish brown spots on upper epidermis. The surviving plants were uprooted and separated into root, stem and leaf on 90th day after experimentation and analyzed for Fe and the results are given in Fig. 1. From the Fig. 1a, it is analysed that the content of iron with respect to the soil application was found to be more in root than in stem and leaf similarly from the Fig. 1b it is noted that the content of iron in soil increases with increase in the external application of iron. This was proved by the positive and significant correlation coefficient worked out between soil applied iron and the iron content in various plant parts. The iron content in root, stem, leaf and soil with respect to the external addition of soil applied iron were r = 0.942, 0.996, 0.990 and 0.937, respectively is highly significant at p = 0.01 (Table 2). The leaves are analyzed for biochemical parameters and some enzyme activities. No visible toxicity symptoms were appeared on the plants planted in soil having 50 mg Fe kg-1, while it was noticed at 500 mg Fe treatment. Hence, the Fe content observed in leaf, stem and root was taken as the limits of toxicity. Accordingly 1140, 570 and 547 mg kg-1 was considered as the critical toxicity limit of leaf, stem and root of tea plant.
Biochemical parameters: The polyphenols content decreased sharply with respect to the addition of increased iron dosage. The plants which received more than 750 mg kg-1 exhibited significantly (r = -0.801 p = 0.05) lower amount (below 160 g kg-1) of polyphenols when compared to that of control (Fig. 2). This observation confirms our earlier findings that high concentration of Fe may result in localized formation of Fe polyphenols complex (Venkatesan et al., 2005, 2006).
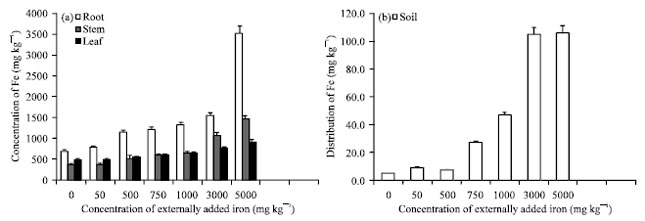 | |
| Fig. 1 (a-b): | (a) Absorption and distribution of iron to various plant parts and (b) levels of available iron in soil after 120 days of external addition. The error bars represent the relative standard deviation |
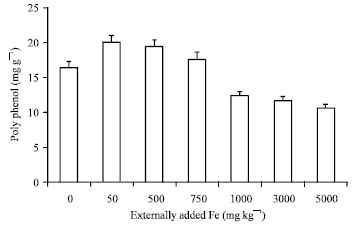 | |
| Fig. 2: | Influence of externally added iron on poly phenol content in leaf. The error bars represent the relative standard deviation |
| Table 1: | Correlation coefficient worked out between externally added iron, enzymes and biochemical parameters in green leaves |
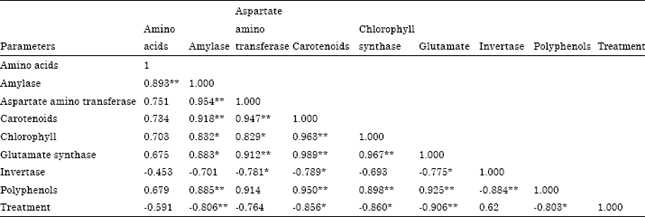 | |
| *Significant at 5% level. **Significant at 1% level | |
| Table 2: | Correlation coefficient worked out between externally added iron and iron content in various plant parts |
 | |
| *Significant at 5% level. **Significant at 1% level | |
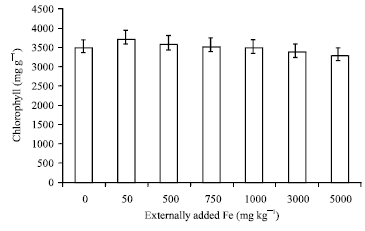 | |
| Fig. 3: | Influence of externally added iron on chlorophyll content in leaf. The error bars represent the relative standard deviation |
This is proved by the negative correlation coefficient obtained between soil applied Fe and leaf polyphenols (r = -0.803; p = 0.05) (Table 1). The chlorophyll content estimated in tea leaves also showed the same trend as that of polyphenols (Fig. 3) (r = -0.860; p = 0.05) (Table 1). This is because excess of Fe must have catalyzed the generation of active O2 species or free radicals which could have eventually oxidized the chlorophyll and subsequently lead to a decrease in chlorophyll content (Monteiro and Winterbourn, 1988) at higher treatment.
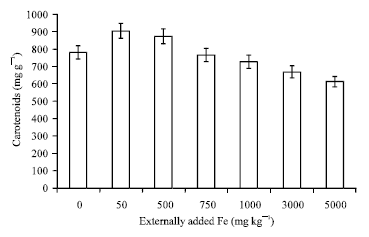 | |
| Fig. 4: | Influence of externally added iron on carotenoid content in leaf. The error bars represent the relative standard deviation |
In the study conducted by Arunachalam et al. (2009) on the impact of iron concentration on the plant physiology reported the decrease in chlorophyll content with increased iron concentration. The Carotenoids content also followed the same trend as that of chlorophyll (Fig. 4) (r = -0.856; p = 0.05) (Table 1). This uniformity in trend is due to the uniformity of pigment synthesis which may be due to the equivalent synthetic activity of chlorophyll and Carotenoids (Wilkinson and Ohki, 1988) and moreover carotenoids act as one of the important metabolite which helps in protecting the cellular and sub cellular system from the cytotoxic effects of these reactive oxygen species (Agarwal et al., 2006) by quenching the singlet oxygen (Prasad and Bisht, 2011). But with decrease in carotenoids content the possibility of protecting the plant cellular systems is in vein which leads to the damage of organic constituent of the living cell. Chlorophyll and carotenoids showed a negative and significant correlation co-efficient (r = -0.860 p = 0.05; r = -0.856 p = 0.05) (Table 1) with soil applied Fe.
Enzyme activity
Amylase: Fe has some influence on amylase activity which is an enzyme participating in carbohydrate metabolism. The activity is linear upto 500 mg Fe kg-1 above which there was a sharp decline. The hydrolysis of starch into maltose is carried out in the presence of carbohydrase enzyme amylase as catalyst. The activity is measured in the form of reducing sugar. The decrease in activity with increase in the concentration of Fe is due to fact that Fe forms complex with carbohydrate compounds (Hofner, 1970) like maltose (Fig. 5). A negative and significant (r = -0.806; p = 0.01) (Table 1) correlation coefficient exihibited between soil applied Fe and amylase activity proved that the enzyme activity will be highly disturbed with increase in the externally added Fe.
Invertase: Invertase which is also a carbohydrase enzyme, catalyse sucrose into glucose and fructose which is measured in the form of reducing sugar. The result is little contrary to amylase activity. The percentage of reducing sugar formed has been increased upto 3000 mg Fe and then decreased at 5000 mg. At 50 and 500 mg treatment the percentage of reducing sugar formation was almost equal. A slight increase was noted with 750 mg again at 1000 and 3000 mg treatment a drastic increase was noted, suddenly at higher treatment ie., at 5000 mg the activity was totally reduced (Fig. 6).
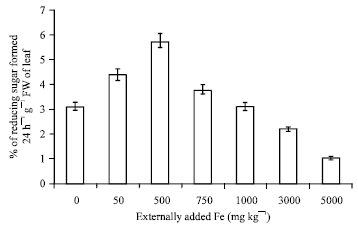 | |
| Fig. 5: | Influence of externally added iron on amylase activity in leaf. The error bars represent the relative standard deviation |
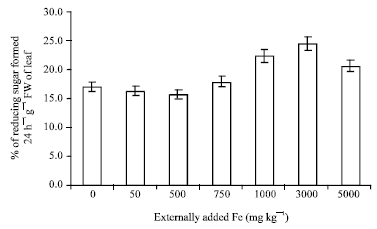 | |
| Fig. 6: | Influence of externally added iron on invertase activity in leaf. The error bars represent the relative standard deviation |
As mentioned elsewhere the reduction in the formation of reducing sugar above 3000 mg Fe may be due to the formation of Fe complex with reducing sugar (Hofner, 1970). Though the activity was reduced at 5000 mg treatment the statistical data proved that it was not significant (Table 1).
Glutamate synthase and amino acid: The trend observed in glutamine synthase and amino acids are almost the same. The activities of both these enzymes were high with 50 mg Fe and then decreased sharply (Fig. 7, 8). It is known that the assimilation of NH4+ into amino acids occurs via joint action of glutamine synthetase and glutamate synthase. Glutamate synthase catalyses the transfer of amide group from glutamine to alpha-ketoglutarate to yield two molecules of glutamate. This reaction is agreed to be the primary route of nitrogen assimilation in plants (Temple et al., 1998). Hence, when the activity decreased the formation of amino acid is also decreased which leads to lack of nitrogen assimilation in plants.
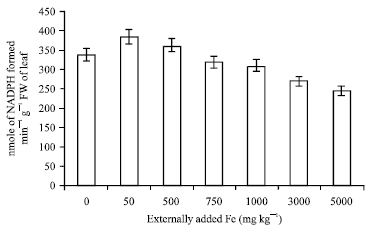 | |
| Fig. 7: | Influence of externally added iron on glutamate synthase activity in leaf. The error bars represent the relative standard deviation |
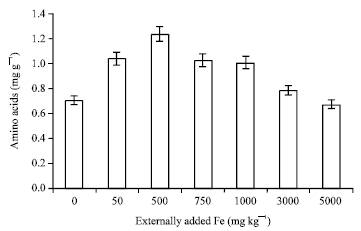 | |
| Fig. 8: | Influence of externally added iron on amino acid content in leaf. The error bars represent the relative standard deviation |
A correlation coefficient worked out between soil applied iron and glutamate synthase activity was negative and significant (r = -0.906; p = 0.01) (Table 1) whereas the correlation coefficient for amino acid content was negative with increase in the soil applied iron but it was not significant.
Aspartate amino transferase: Aspartate aminotransferase catalyses the following reaction:
It was formerly called Glutamic Oxaloacetic Transaminase (GOT). It catalyses the transamination of L-aspartate and 2-oxoglutarate to oxaloacetate and glutamate (Raja et al., 2011). The regeneration of 2-oxglutarate is necessary for ammonia assimilation through glutamate dehydrogenase (Sadasivam and Manickam, 1996). Similarly, aspartate is an important nitrogen transport compound in plants (Temple et al., 1998). The activity of aspartate aminotransferase increased till 500 mg Fe kg-1 and then decreased with increase in Fe concentration (Fig. 9). When the activity decreased the formation of 2-oxoglutarate and aspartate would be reduced. Hence, increase in Fe concentration is expected to reduce the nitrogen metabolism in tea plants. The activity of this enzyme showed a negative and significant correlation with applied iron (r = -0.764; p = 0.05).
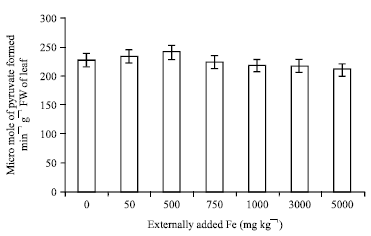 | |
| Fig. 9: | Influence of externally added iron on aspartate amino transferase activity in leaf. The error bars represent the relative standard deviation |
CONCLUSION
From the above study it is concluded that the iron concentration decreases the concentration of pigments like chlorophyll and carotenoid. The polyphenol content which plays a vital role on the quality of made tea was also affected by the localized formation of Fe-Polyphenol complex. So the quality of the made tea will be affected with the excess Fe concentration. The enzymatic activity of amylase, invertase, aspartate amino transferase and glutamate synthase was also drastically affected by the excess Fe concentration. Overall it is concluded that the excess Fe concentration leads to the plant mortality.
ACKNOWLEDGMENTS
The authors thank Dr. N. Muraleedharan, Adviser and Dr. P. Mohan Kumar, Director, UPASI Tea Research Foundation for constant encouragement throughout the study and critically evaluating the manuscript. The financial assistance provided by the Tea Board, Government of India under X Five Year Plan is gratefully acknowledged.
REFERENCES
- Agarwal, S., R.K. Sairam, R.C. Meena, A. Tyagi and G.C. Srivastava, 2006. Effect of excess and deficient levels of iron and copper on oxidative stress and antioxidant enzymes activity in wheat. J. Plant Sci., 1: 86-97.
CrossRefDirect Link - Arunachalam, R., K. Paulkumar, A.J.A. Ranjitsingh and G. Annadurai, 2009. Environmental assessment due to air pollution near iron smelting industry. J. Environ. Sci. Technol., 2: 179-186.
CrossRefDirect Link - Asad, A. and R. Rafique, 2000. Effect of zinc, copper, iron, manganese and boron on the yield and yield components of wheat crop in tehsil Peshawar. Pak. J. Biol. Sci., 3: 1615-1620.
CrossRefDirect Link - Becana, M., J.F. Moran and I. Iturbe-Ormaetxe, 1998. Iron dependant oxygen free radical generation in plants subjected to environmental stresses: Toxicity and antioxidants. Plant Soil, 201: 137-147.
CrossRef - Becker, M. and F. Asch, 2005. Iron toxicity in rice-condition and management concepts. J. Plant Nutr. Soil Sci., 168: 558-573.
Direct Link - Cabrera, C., R.Gimenez and M.C. Lopez, 2003. Determination of tea components with antioxidant activity. J. Agric. Food Chem., 51: 4427-4435.
CrossRefDirect Link - Chung, F.L., J. Schwartz, C.R. Herzog and Y.M. Yang, 2003. Tea and cancer prevention: Studies in animals and humans. J. Nutr., 133: 3268S-3274S.
PubMedDirect Link - Chen, S.X. and P. Schopfer, 1999. Hydroxyl-radical production in physiological reactions: A novels function of peroxidase. Eur. J. Biochem., 260: 726-735.
CrossRefPubMedDirect Link - Abdel-Kader, D.Z.E.A., 2007. Role of nitric oxide on iron homeostasis, chlorophyll biosynthesis and antioxidants system in two wheat cultivars. Am. J. Plant Physiol., 2: 237-250.
CrossRefDirect Link - Halliwell, B. and J.M. Gutteridge, 1984. Oxygen toxicity, oxygen radicals, transition metals and disease. Biochem. J., 219: 1-14.
PubMed - Hemalatha, K.V., S. Venkatesan and S. Jayaganesh, 2006. Characterisation of iron toxicity and its influence on nutrient uptake in tea. J. Plantation Crops, 34: 268-272.
Direct Link - Kerkeb, L. and E. Connolly, 2006. Iron transport and metabolism in plants. Genet. Eng., 27: 119-140.
PubMedDirect Link - Madhu, M., D.C. Sahoo, V.N. Sharda and A.K. Sikka, 2011. Rainwater-use efficiency of tea (Camellia sinensis (L.)) under different conservation measures in the high hills of South India. Applied Geogr., 31: 450-455.
CrossRef - Monteiro, H.P. and C.C. Winterbourn, 1988. The superoxide-dependant transfer of iron from ferritin to transferring and lactoferrin. Biochem. J., 256: 923-928.
Direct Link - Prasad, K. and G. Bisht, 2011. Evaluation of nutritive, Antioxidant and mineral composition of Pavetta indica Linn. leaves. Res. J. Phytochem., 5: 54-59.
CrossRefDirect Link - Raja, M.M.M., A. Raja, M.M. Imran, A.M.I. Santha and A. Devasena, 2011. Enzymes application in diagonostic approach. Biotechnology, 10: 51-59.
Direct Link - Temple, S.J., C.P. Vance and J.S. Gantt, 1998. Glutamate synthase and nitrogen assimilation. Trends Plant Sci., 3: 51-56.
CrossRefDirect Link - Thengbai, P. and B.A. Goodman, 2000. Free radicals generation and post anoxic injury in rice grown in an iron-toxic soil. Proceedings of 9th International Symposium on Iron Nutrition and Interactions in Plants, July 20-25, Stuttgart, Germany, pp: 1887-1900.
CrossRefDirect Link - VandeCasteele, J.P., J. Lemal and M. Coudest, 1975. Pathways and regulation of glutamate synthesis in a Corynebacteriurn sp. overproducing glutamate. J. Gen. Microbiol., 90: 178-180.
PubMedDirect Link - Venkatesan, S., S. Murugesan, V.K. Senthur-Pandian and M.N.K. Ganapathy, 2005. Impact of sources and doses of potassium on biochemical and greenleaf parameters of tea. Food Chem., 90: 535-539.
CrossRef - Wellburn, A.R., 1994. The spectral determination of chlorophyll a and chlorophyll b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol., 144: 307-313.
Direct Link - Wilkinson, R.E. and K. Ohki, 1988. Influence of manganese deficiency and toxicity on isoprenoid syntheses. Plant Physiol., 87: 841-846.
PubMedDirect Link - Alscher, R.G., J.L. Donahue and C.L. Cramer, 1997. Reactive oxygen species and antioxidants: Relationship in green cells. Physiol. Plant., 100: 224-233.
CrossRefDirect Link