
P.C. Chikezie
Department of Biochemistry, Imo State University, Owerri, Imo State, Nigeria
LiveDNA: 234.10425
ORCID: 0000-0001-5066-724X
Asian Journal of Biochemistry
Year: 2011 | Volume: 6 | Issue: 5 | Page No.: 373-383







ABSTRACT
In vitro study was carried out to investigate levels of oxidative stress indicators of sickle erythrocytes incubated in aqueous extracts of Anacardium occidentale, Psidium guajava and Terminalia catappa for 12 h. At regular time intervals of 3 h, portions of the incubation mixtures were withdrawn and spectrophotometric method was used to assay for levels of erythrocyte Malondialdehyde (MDA) and methaemoglobin (Met. Hb%). The control analysis showed that within the experimental time, erythrocyte MDA increased from 2.45±0.35 to 3.13±0.59 mmol mL-1 (p>0.05; pvalue = 0.801176). Erythrocyte MDA concentrations in the presence of the three extracts were higher than the control samples at t = 3 h (p>0.05; p value = 0.963253). Compared with the control samples at the given time (t) intervals, extract of T. catappa exhibited the highest capacity to cause reduction of erythrocyte MDA ([T. catappa] = 800 mg%; [MDA] = 2.89±0.33 mmol mL-1; t = 12 h). Erythrocyte Met. Hb% increased from 2.42±0.55 to 2.51±0.49% (p>0.05; p value = 0.995171) in the control samples within 12 h. Incubation of sickle erythrocytes with extract of (P. guajava) = 800 mg% for 9 h caused reduction of Met. Hb% from 2.49±0.49 to 2.29±0.45%; p>0.05; p = 0.983519. Extracts of A. occidentale, P. guajava and T. catappa exhibited variable capacities to hinder lipid peroxidation but did not cause corresponding reduction in erythrocyte Met. Hb%, exemplified by negative correlation between the two oxidative stress indicators in the presence of T. catappa and higher concentrations of A. occidentale and P. guajava.
PDF Abstract XML References Citation
Received: October 28, 2010;
Accepted: June 06, 2011;
Published: July 29, 2011
How to cite this article
P.C. Chikezie, 2011. Oxidative Stress Indicators of Human Sickle Erythrocytes Incubated in Aqueous Extracts of Three Medicinal Plants. Asian Journal of Biochemistry, 6: 373-383.
DOI: 10.3923/ajb.2011.373.383
URL: https://scialert.net/abstract/?doi=ajb.2011.373.383
DOI: 10.3923/ajb.2011.373.383
URL: https://scialert.net/abstract/?doi=ajb.2011.373.383
INTRODUCTION
Oxidative stress is caused by accumulation of Reactive Oxygen Species (ROS) (Shyur et al., 2005; Richards et al., 2007) produced as normal by-products of cellular metabolism (Richards et al., 2007; Breusegem and Mittler, 2008), exposure to ionic/electromagnetic radiations and some environmental pollutants (Tiwari, 2001; Aqil et al., 2006; Doss et al., 2009). These reactive species are capable of damaging diverse biomolecules and cell structures in which lipids are probably the most susceptible (Saengkhae et al., 2007; Okwu, 2007; Geetha et al., 2007) when cellular levels are not controlled by appropriate antioxidant scavenging systems (Neupane et al., 2008). Whereas erythrocyte of all genotypes are particularly sensitive to oxidative stress, when compared with normal erythrocytes, sickle erythrocytes spontaneously generate approximately twice as much superoxide (.O2-), peroxide (H2O2) and hydroxyl (.OH) radicals (Hebbel et al., 1982) and increasing evidence suggest that lipid peroxidation may be an important factor in sickle cell anaemia (Tamer et al., 2000).
Specifically, sickle erythrocytes and their membrane structures are susceptible to endogenous free radical-mediated oxidative damage that correlates with the proportion of irreversibly sickled erythrocytes (Aslan et al., 2000). Furthermore, accumulation of hydrogen peroxide (H2O2) decreases the half life of erythrocytes by increasing oxidation of polyunsaturated fatty acids of membrane constituents (Nijs and De Meirleir, 2004) and can oxidize haemoglobin to methaemoglobin (Neupane et al., 2008). Methaemoglobin does not bind reversibly with oxygen. One of the toxic end products of lipid peroxidation is Malondialdehyde (MDA) (Tamer et al., 2000). Sickle erythrocytes contain increased amount of MDA and evidence of abnormal amino group cross-linking by MDA has been demonstrated in lipid extract of sickle erythrocyte membrane preparations (Dalle-Donne et al., 2006).
Erythrocytes like other biological cells are supplied with diverse protective antioxidant mechanisms in order to counteract the toxic effects of ROS (Champe et al., 2005; Forchetti et al., 2006; Chikezie, 2011a, b). Antioxidants function as modulators of cellular homeostasis including detoxification of oxyradicals and metals as well as potent free radical scavenger. Erythrocytes reduced Glutathione (GSH) is one of the major non-enzymic endogenous antioxidants, protecting tissue against ROS (Dalle-Donne et al., 2006). Other antioxidants are α-tocopherol (Muller, 2004; Zabri et al., 2008), uric acid (Glantzounis et al., 2005; Sautin and Johnson, 2008; Bowman et al., 2010), ascorbic acid (Sati et al., 2010), β-carotenoids (Lee et al., 2000) and varieties of plant secondary metabolites such as flavonoid and related polyphenolic compounds (Middleton et al., 2000; Rice-Evans, 2001; Srivastava et al., 2011). Notable erythrocyte enzymatic ROS scarvenging systems include glutathione reductase (Forchetti et al., 2006), glutathione peroxidase (Manfredini et al., 2008) and glucose-6-phosphate dehydrogenase (Champe et al., 2005; Ojo et al., 2006). Others are superoxide dismutase (Ekor et al., 2006; Saengkhae et al., 2007; Ojha et al., 2010), catalase (Goth, 1991; Pennings et al., 1999; Chandrasena et al., 2006; Shanmugarajan and Devaki, 2008), peroxiredoxins (Neumann et al., 2003; Low et al., 2007) and NADH-methaemoglobin reductase (Mallory, 2003; Chikezie, 2011b).
Varieties of xenobiotics of plant origin such as fava beans extract (Fava fava) have been reported as agents that can interfere with the redox status of human erythrocytes especially in individuals with impaired glucose-6-phosphate dehydrogenase activity (Champe et al., 2005; Ojo et al., 2006). In the same vein, scavenging activities for free radicals by natural products of plant origin have been widely reported Shyur et al. (2005), Aqil et al. (2006), Buricova and Reblova (2008), Muanda et al. (2009), Veeru et al. (2009) and Sati et al. (2010). This study ascertained erythrocyte level of oxidative stress by estimating the concentrations of oxidative stress indicators, namely, MDA and methaemoglobin of sickle erythrocytes incubated in aqueous extracts of three medicinal plants: Anacardium occidentale, Psidium guajava and Terminalia catappa.
MATERIALS AND METHODS
Collection of plant specimens: Fresh leaves samples of A. occidentale, P. guajava and T. catappa were harvested between July and August, 2010, from trees growing within the environment of Imo State University, Owerri, Nigeria. The plant specimens were identified and authenticated by Dr. F. N. Mbagwu at the Herbarium, Department of Plant Science and Biothechnology. A voucher specimen was deposited at the Herbarium for reference purposes.
Preparation of aqueous extracts of plant specimens: The samples were washed under continuous current of distilled water for 15 min and air dried at room temperature (24°C) for 60 min. The separate leaves were dried for 5 h in an oven at 60°C to become crispy and ground with ceramic mortar and pestle. To each specimen, two grams (2 g) of the pulverized sample was suspended in 100 mL of distilled water and allowed to stand for 6 h at 37°C. Aqueous extracts (2 g%) of A. occidentale, P. guajava and T. catappa leaves were obtained by simple filteration method with Whatman No. 2 filter paper. The filtrates were centrifuged at 1200xg for 5 min to remove tissue debris. The supernatants were carefully harvested with pasteur pipette into sterile test tubes and kept at 4°C in a refrigerator for at least 24 h before subsequent tests. Serial dilutions of the aqueous extracts in the order of 200, 400, 600 and 800 mg% were used for analyses.
Collection of blood samples/preparation of erythrocyte haemolysate: Five milliliters (5.0 mL) of human venous blood samples of HbSS genotype were collected by venipuncture and stored in EDTA anticoagulant tubes. The blood samples were obtained between July and August, 2010, from nine (9) male volunteers (59-79 kg) between the age bracket of 21-34 year attending clinics at the Federal Medical Center (FMC), Imo State University Teaching Hospital (IMSUTH), Orlu, St. John Clinic/Medical Diagnostic Laboratories, Avigram Medical Diagnostic Laboratories and Qualitech Medical Diagnostic Laboratories. These Centers are located in Owerri, Imo State, Nigeria. The Institutional Review Board of the Department of Biochemistry, Imo State University, Owerri, Nigeria, granted approval for this study and all blood donors signed informed consent form. This study was in accordance with the ethical principles that have their origins in the Declaration of Helsinki.
The erythrocytes were washed by centrifugation method as described by Tsakiris et al. (2005). To remove platelets and leucocytes, the sediment was re-suspended in 3.0 mL of Phosphate-Buffered Saline (PBS) solution, pH 7.4 and passed through a column (3.5 cm in a 30 mL syringe) of cellulose-microcrystalline cellulose (ratio w/w 1:1) as described by Kalra et al. (1981). The eluted fraction was passed twice through a new column of cellulose-microcrystalline cellulose (ratio 1:1 w/w) to obtain erythrocyte suspension sufficiently devoid of leucocytes and platelets. The isolated erythrocytes were lysed by freezing/thawing as described by Galbraith and Watts (1980) and Kamber et al. (1984). The erythrocyte haemolysates were finally re-suspended in 1.0 mL of the buffer and stored at -70°C until analyses (Pennings et al., 1999).
Experimental design: A portion of 0.2 mL aqueous extracts of A. occidentale, P. guajava and T. catappa of increasing concentrations in the order: 200, 400, 600 and 800 mg % w/v were added to corresponding test tubes containing 0.8 mL of erythrocyte haemolysate (ratio 1:4 v/v). The incubation mixture was allowed to stand at a regulated temperature of 37°C in a water bath. At regular time intervals of 3 for 12 h, aliquots of 0.2 mL of the incubation mixture were withdrawn and used for the determinations of erythrocyte MDA and methaemoglobin concentrations.
Determinations of erythrocyte malondialdehyde and methaemoglobin concentrations: Determination of erythrocyte MDA was by method described by Tjahjani et al. (2008) with minor modifications. A mixture of 20% Trichloracetic Acid (TCA) and 0.67% Thiobarbituric Acid (TBA) in a ratio of 2:1 was added into a test tube. A volume of 0.2 mL of erythrocyte haemolysate was introduced in the mixture and boiled for 10 min in a water bath. After cooling to 24°C, the mixture was centrifuged at 3,000xg for 10 min. The absorbance of supernatant was read with a spectrophotometer (SPECTRONIC 20, Labtech-Digital Blood Analyzer®) at maximum wavelength (λmax) = 532 nm. The values of absorbance of the samples were converted to MDA concentrations using the MDA standard curve (Schemuk et al., 2002). Determination of methaemoglobin content of erythrocyte haemolysate was by modification of the method of Evelyn and Malloy (1938), as described by Akomopong et al. (2000).
Statistical analysis: The results were expressed in terms of arithmetic Mean±Standard Deviation (SD). The correlation coefficients between the results were determined with Microsoft Office Excel, 2007 version and data were analyzed by Student’s t-test as described by Pearson and Hartley (1966). Values of p<0.05 were considered statistically significant.
RESULTS
The control analysis showed that within the limit of experimental time 0-12 h, erythrocyte MDA concentration increased from 2.45±0.35 to 3.13±0.59 mmol mL-1 (Fig. 1-3). Also, incubation of sickle erythrocytes in aqueous extracts of A. occidentale, P. guajava and T. catappa, showed increasing erythrocyte MDA concentrations with progression of incubation time.
However, compared with the corresponding control samples at every given time (t) interval, when t>3 h, erythrocytes suspended in the three aqueous extracts exhibited decreased MDA concentrations (p>0.05; p value = 0.963253). Erythrocyte MDA concentrations in the presence of the three extracts were generally higher than the control samples at t = 3 h. Specifically, when the erythrocytes were incubated for 12 h in aqueous extract of A. occidentale, MDA concentration was 2.92±0.25 mmol mL-1 (control = 3.13±0.59 mmol mL-1; p> 0.05; p value = 0.9776), representing a decrease of 6.71% of MDA concentration within the period of incubation (Fig. 1).
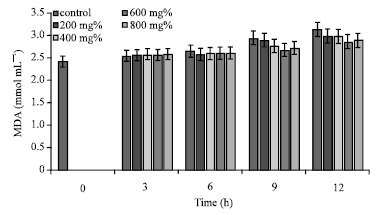 | |
| Fig. 1: | MDA concentrations of sickle erythrocytes incubated in aqueous extract of A. occidentale |
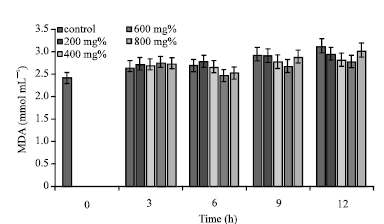 | |
| Fig. 2: | MDA concentrations of sickle erythrocytes incubated in aqueous extract of P. guajava |
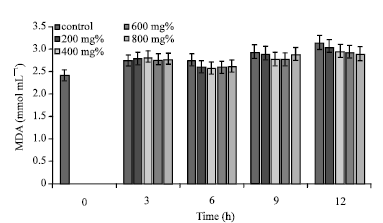 | |
| Fig. 3: | MDA concentrations of sickle erythrocytes incubated in aqueous extract of T. catappa |
Whereas in the control sample, peak concentaration of erythrocyte MDA concentration was 3.13±0.59 mmol mL-1 (t = 12 h), erythrocytes incubated in aqueous extracts of P. guajava and T. catappa, exhibited peak MDA concentration in the following specifications: ([P. guajava] = 800 mg%, [MDA] = 3.04±0.38 mmol mL-1; [T. catappa] = 200 mg%, [MDA] = 3.04±0.23 mmol mL-1) (Fig. 2).
At t = 12 h, aqueous extract of T. catappa, exhibited the highest capacity to cause the reduction of erythrocyte MDA concentration ([T. catappa] = 800 mg%; [MDA] = 2.89±0.33 mmol mL-1), representing 7.66% reduction in erythrocyte MDA concentration (Fig. 3). Notably, Fig. 3 showed that in a concentration dependent manner, aqueous extract of T. catappa caused decreased erythrocyte MDA concentrations between incubation time of 9 and 12 h. The levels of erythrocyte MDA in the presence of 800 mg% concentration of A. occidentale and P. guajava were elevated compared to lower concentrations of the same extracts.
Erythrocyte MDA concentrations in the presence of 800 mg% of A. occidentale and P. guajava (r = 0.845082) showed a higher positive correlation than between A. occidentale and T. catappa (r = 0.746903). The MDA contents of sickle erythrocytes incubated in the four experimental concentrations of A. occidentale and P. guajava exhibited low positive correlation at t = 3 h; (r = 0.271896).
Erythrocyte Met. Hb% increased from 2.42±0.55 to 2.51±0.49% in the control sample within the duration of 12 h (Fig. 4-6). Although at t = 3 h, aqueous extract of A. occidentale (except 800 mg%) caused increased Met. Hb%, the values were not significantly different (p>0.05) from the control samples. Further increases in incubation time (t>3) engendered comparatively declining levels of erythrocyte Met. Hb% except at t = 12 h (Fig. 4). Although aqueous extract of [A. occidentale] = 200 mg% caused decreased erythrocyte Met. Hb% from 2.49±0.49 to 2.34±0.65% within 6 h of incubation, the value increased to 2.55±0.43% at t = 12 h (Fig. 4).
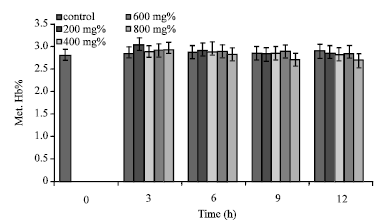
Incubation of sickle erythrocytes in aqueous extract of [P. guajava] = 800 mg% for 9 h caused the reduction of Met. Hb% from 2.49±0.49 to 2.29±0.45%. At [P. guajava] = 600 mg%, erythrocyte Met. Hb% gave value of 2.55±0.43% (t = 3 h) which was not significantly (p>0.05; pvalue = 0.996121) higher than the control value (Fig. 5). Also, in the presence of aqueous extract of [P. guajava] = 200 mg%, value of erythrocyte Met. Hb%, 2.46±0.43%; at t = 9 h, was not significantly different (p>0.05; p = 0.999778) from the control sample.
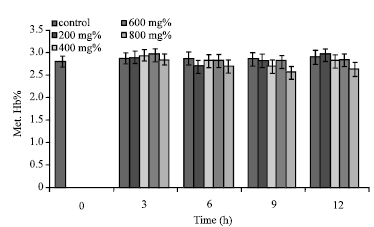 | |
| Fig. 4: | Met. Hb% of sickle erythrocytes incubated in aqueous extract of A. occidentale |
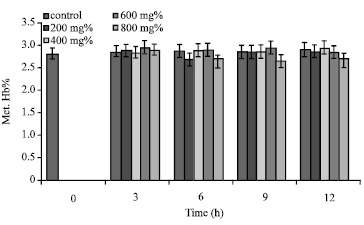 | |
| Fig. 5: | Met. Hb% of sickle erythrocytes incubated in aqueous extract of P. guajava |
 | |
| Fig. 6: | Met. Hb% of sickle erythrocytes incubated in aqueous extract of T. catappa |
In addition, 200 mg% of T. catappa caused increased erythrocyte Met. Hb% within the experimental period of 6 h (Fig. 6). Generally, aqueous extract of T. catappa exhibited low capacity to cause reduction in erythrocyte Met. Hb%. Statistical evalution shows that erythrocyte Met. Hb% in the presence of 600 mg% of P. guajava and T. catappa displayed high positive correlation (r = 0.965535). Also, 800 mg% of A. occidentale and P. guajava exhibited positive correlation (r = 0.878868) with respect to erythrocyte Met. Hb%.
| Table 1: | Correlation between changes in MDA concentration and Met. Hb% of sickle erythrocyte incubated in aqueous extracts of A. occidentale, P. guajava and T. catappa for 12 h |
 | |
From the data presented in Table 1, erythrocyte Met. Hb% and MDA concentration showed significant positive correlation in the presence of aqueous extracts of 200 mg% A. occidentale and 400 mg% P. guajava. Notably, erythrocytes incubated in aqueous extracts of T. catappa and concentrations of A. occidentale and P. guajava>400 mg% showed negative correlation.
DISCUSSION
The increasing erythrocyte MDA concentrations of the control samples with time were obvious reflection of production and accumulation of ROS, engendered by normal metabolic processes in these cells. Tamer et al. (2000) had earlier reported that sickle erythrocytes generate ROS spontaneously and contain high level of MDA, a by-product of lipid peroxidation. It is worthwhile to note that several abnormalies associated with sickle erythrocyte are inextricably connected with the relatively high level of oxidative stressors in this erythrocyte genotype (Manfredini et al., 2008; Repka and Hebbel, 1991; Dhalla et al., 2000). From the results showed in Fig. 1-3, the increasing erythrocyte MDA concentrations of the control and test samples was an indication of time dependent progression of lipid peroxidation in both samples. However, relatively lower MDA concentrations of the test samples compared to the control samples at the given time intervals revealed that lipid peroxidation was retarded by the three plant extracts in connection with their corresponding concentrations. The present findings are in concord with the report of Lam et al. (2007). They noted that γ-irradation induced oxidative stress indicators were significantly reduced in rats after orally administered with Aloe vera. Prevoius findings have attributed the capacities of Zingiber officinale, Aloe vera (L.) and Rheum rhabarbarum (L.) extracts to reduce lipid peroxidation to the presence of phenolic compounds in these plants (Aaviksaar et al., 2003; Lam et al., 2007). Also, membrane protective activities of diverse plant extracts have been atrributed to their antioxidant content and capacity to impede membrane lipid peroxidation (Hu and Kitts, 2000; Okpuzor et al., 2008; Buricova and Reblova, 2008; Mudgal et al., 2010). However, since 800 mg% extract concentration of A. occidentale and P. guajava did not cause the anticipated level of hinderance to lipid peroxidation (r = 0.845082), it is envisaged that higher concentrations of the two plant extracts may promote it. It is worthwhile to mention that Paiva and Russell (1999) had prevoiusly reported that antioxidant activity of carotenoids (including β-carotene) may exhibit adverse effects when present in high dose.
The present result showed that Met. Hb% of sickle erythrocyte was significantly (p<0.05; p = 0.0489323) higher than normal physiologic concentration (Met. Hb% = 1.50) as reported by Tietz (1976) and Chikezie (2009) for HbAA erythrocyte genotype. Noteworthy, the level of erythrocyte methaemoglobin reported here is comparable to those presented else where (Van Kuijk et al., 1987; Chikezie, 2009; Chikezie, 2011b). The primary reason for the relatively high concentration of oxidized haemoglobin is also connected with excessive production and accumulation of ROS compared with other human erythrocyte genotypes (Van Kuijk et al., 1987). The moderate reduction in Met. Hb% in erythrocyte incubated in aqueous extracts of A. occidentale, P. guajava and T. catappa is attributed to their antioxidant activity as earlier discussed. However, previous reports stated that the association of certain methaemoglobinopathies such as HbMBoston, HbMIwate, HbMHydepark and HbMHammersmith with sickle erythrocytes contributed to insignificant reduction in erythrocyte Met. Hb% (Mayes, 1983). These variant haemoglobin molecules are noted for tendency towards spontaneous oxidation in vivo and resistant to enzymic and non-enzymic reduction mechanisms.
CONCLUSION
Based on the levels of oxidative stress indicators in the present study, aqueous extracts of A. occidentale, P. guajava and T. catappa exhibited variable capacities to hinder lipid peroxidation but did not cause corresponding reduction in erythrocyte Met. Hb%, exemplified by negative correlation between the two oxidative stress indicators in the presence of T. catappa and higher concentrations of A. occidentale and P. guajava.
REFERENCES
- Aaviksaar, A., M. Haga, K. Kuzina, T. Pussa, A. Raal and G. Tsoupras, 2003. Hydroxystilbenes in the roots of Rheum rhaponticum. Proc. Estonian Acad. Sci. Chem., 52: 99-107.
Direct Link - Akomopong, T., N. Ghori and K. Haldar, 2000. In vitro activity of riboflavin against the human malaria parasite Plasmodium falciparum. Antimicrobial Agents Chemothera, 44: 88-96.
Direct Link - Aqil, F., I. Ahmad and Z. Mehmood, 2006. Antioxidant and free radical scavenging properties of twelve traditionally used Indian medicinal plants. Turk. J. Biol., 30: 177-183.
Direct Link - Bowman, G.L., J. Shannon, B. Frei, J.A. Kaye and J.F. Quinn, 2010. Uric acid as a CNS antioxidant. J. Alzheimers Dis., 19: 1331-1336.
PubMedDirect Link - Buricova, L. and Z. Reblova, 2008. Czech medicinal plants as possible sources of antioxidants. Czech J. Food Sci., 26: 132-138.
Direct Link - Chandrasena, L.G., S. Chackrewarthy, P. Teckla, M.J. Perera and D. de Silva, 2006. Erythrocyte antioxidant enzymes in patients with cataract. Ann. Clin. Lab. Sci., 36: 201-204.
Direct Link - Chikezie, P.C., 2009. Comparative methaemoglobin concentrations of three erythrocyte genotypes (HbAA, HbAS and HbSS) of male participants administered with five antimalarial drugs. Afr. J. Biochem. Res., 3: 266-271.
Direct Link - Chikezie, P.C., 2011. Comparative in vitro osmotic stability of three human erythrocyte genotypes in the presence of quinine and chloroquine phosphate. Asian J. Biochem., 6: 55-64.
CrossRefDirect Link - Chikezie, P.C., 2011. Methaemoglobin concentration and NADH-methaemoglobin reductaseactivity of three human erythrocyte genotypes. Asian J. Biochem., 6: 98-103.
CrossRefDirect Link - Dalle-Donne, I., R. Rossi, R. Colombo, D. Giustarini and A. Milzani, 2006. Biomarkers of oxidative damage in human disease. Clin. Chem., 52: 601-623.
CrossRefDirect Link - Dhalla, N.S., A.B. Elmoselhi, T. Hata and N. Makino, 2000. Status of myocardial antioxidants in ischemia-reperfusion injury. Cardiovasc. Res., 47: 446-456.
CrossRefDirect Link - Doss, A., A.P.A. Doss and R. Dhanabalan, 2009. In vitro antioxidant properties of certain indigenous medicinal plants from Western Ghats of India. Internet J. Nutri. Wellness, Vol. 7.
Direct Link - Ekor, M., G.K.A. Adepoju and A.A. Epoyun, 2006. Protective effect of the methanolic leaf extract of Persea americana (avocado) against paracetamol-induced acute hepatotoxicity in rats. Int. J. Pharmacol., 2: 416-420.
CrossRefDirect Link - Galbraith, D.A. and D.C. Watts, 1980. Changes in some cytoplasmic enzymes from red cells fractionated into age groups by centrifugation in Ficoll/Triosil gradients. Comparison of normal humans and patients with Duchenne muscular dystrophy. Biochem. J., 191: 63-70.
Direct Link - Geetha, A., M.D. Lakshmi Priya, S. Annie Jeyachristy and R. Surendran, 2007. Level of oxidative stress in the red blood cells of patients with liver cirrhosis. Indian J. Med. Res., 126: 204-210.
PubMed - Glantzounis, G.K., E.C. Tsimoyiannis, A.M. Kappas and D.A. Galaris, 2005. Uric acid and oxidative stress. Curr. Pharm. Des., 11: 4145-4151.
PubMedDirect Link - Góth, L., 1991. A simple method for determination of serum catalase activity and revision of reference range. Clin. Chim. Acta, 196: 143-151.
CrossRefPubMedDirect Link - Hebbel, R.P., J.W. Eaton, N. Balasingam and M.H. Steinberg, 1982. Spontaneous oxygen radical generation by sickle erythrocytes. J. Clin. Invest., 70: 1253-1259.
CrossRefDirect Link - Hu, C. and D.D. Kitts, 2000. Studies on the antioxidant activity of Echinacea root extract. J. Agric. Food Chem., 48: 1466-1472.
CrossRefPubMedDirect Link - Kalra, V.K., S.C. Sikka and G.S. Sethi, 1981. Transport of amino acids in gamma-glutamyl transpeptidase-implanted human erythrocytes. J. Biol. Chem., 256: 5567-5571.
PubMedDirect Link - Kamher, E., A. Poyiagi and G. Deliconstantinos, 1984. Modifications in the activities of membrane-bound enzymes during in vivo ageing of human and rabbit erythrocytes. Comp. Biochem. Physiol. B: Comp. Biochem., 77: 95-99.
CrossRefDirect Link - Lam, R.Y.Y., A.Y.H. Woo, P.S. Leung and C.H.K. Cheng, 2007. Antioxidant actions of phenolic compounds found in dietary plants on low-density lipoprotein and erythrocytes in vitro. J. Am. Coll. Nutr., 26: 233-242.
Direct Link - Lee, J., S. Jiang, N. Levine and R. Watson, 2000. Carotenoid supplementation reduces erythema in human skin after simulated solar radiation exposure. PSEM, 223: 170-174.
PubMed - Low, F.M., M.B. Hampton, A.V. Peskin and C.C. Winterbourn, 2007. Peroxiredoxin 2 functions as a noncatalytic scavenger of low-level hydrogen peroxide in the erythrocyte. Blood, 109: 2611-2617.
PubMedDirect Link - Manfredini, V., L.L. Lazzaretti, I.H. Griebeler, A.P. Santin and A.D.V. Brandao et al., 2008. Blood antioxidant parameters in sickle cell anemia patients in steady state. J. Nat. Med. Assoc., 100: 897-902.
PubMedDirect Link - Middleton Jr. E., C. Kandaswami and T.C. Theoharides, 2000. The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharmacol. Rev., 52: 673-751.
PubMedDirect Link - Muanda, F., D. Koné, A. Dicko, R. Soulimani and C. Younos, 2011. Phytochemical composition and antioxidant capacity of three Malian medicinal plant parts. Evidence-Based Complementary Altern. Med., Vol. 2011.
CrossRefDirect Link - Mudgal, V., N. Madaan, A. Mudgal and S. Mishra, 2010. Dietary polyphenols and human health. Asian J. Biochem., 5: 154-162.
CrossRefDirect Link - Muller, S., 2004. Redox and antioxidant systems of the malaria parasite Plasmodium falciparum. Mol. Microbiol., 53: 1291-1305.
CrossRef - Neumann, C.A., D.S. Krause, C.V. Carman, S. Das and D.P. Dubey et al., 2003. Essential role for the peroxiredoxin Prdx1 in erythrocyte antioxidant defence and tumour suppression. Nat., 424: 561-565.
CrossRefDirect Link - Neupane, D.P., S. Majhi, L. Chandra, S. Rijal and N. Baral, 2008. Erythrocyte glutathione status in human visceral leishmaniasis. Indian J. Clin. Biochem., 23: 95-97.
CrossRefDirect Link - Nijs, J. and K. De Meirleir, 2004. Oxidative stress might reduce essential fatty acids in erythrocyte membranes of chronic fatigue syndrome patients. Nutr. Neurosci., 7: 251-253.
Direct Link - Ojha, R., R. Prasad, N. Manzoor and L.A. Khan, 2010. Vitamin C modulates oxidative stress related enzyme activities in Candida albicans. Turk. J. Biochem., 35: 35-40.
Direct Link - Okpuzor, J., O. Adebesin, H. Ogbunugafor and I. Amadi, 2008. The potential of medicinal plants in sickle cell disease control: A review. Int. J. Biomed. Health Sci., 4: 47-55.
Direct Link - Paiva, S.A. and R.M. Russell, 1999. β-carotene and other carotenoids as antioxidants. J. Am. Coll. Nutr., 18: 426-433.
PubMedDirect Link - Pandey, A., R.K. Gupta and R. Srivastava, 2011. Curcumin-the yellow magic. Asian J. Applied Sci., 4: 343-354.
CrossRefDirect Link - Pennings, H.J., P.J. Borm, C.T. Evelo and E.F. Wouters, 1999. Changes in levels of catalase and glutathione in erythrocytes of patients with stable asthma, treated with beclomethasone dipropionate. Eur. Respir. J., 13: 1260-1266.
PubMedDirect Link - Repka, T. and R.P. Hebbel, 1991. Hydroxyl radical formation by sickle erythrocyte membranes: Role of pathologic iron deposits and cytoplasmic reducing agents. Blood, 78: 2753-2758.
PubMedDirect Link - Sati, S.C., N. Sati, U. Rawat and O.P. Sati, 2010. Medicinal plants as a source of antioxidants. Res. J. Phytochem., 4: 213-224.
CrossRefDirect Link - Sautin, Y.Y. and R.J. Johnson, 2008. Uric acid: The oxidant antioxidant paradox. Nucleosides Nucleotides Nucleic Acids, 27: 608-619.
CrossRefDirect Link - Schmuck, G., E. Roehrdanz, R.K. Haynes and R. Kahl, 2002. Neurotoxic mode of action of artemisinin. Antimicrob. Agents Chemother., 46: 821-827.
CrossRefDirect Link - Shanmugarajan, T.S. and T. Devaki, 2008. Ficus hispida Linn. leaf extract possesses antioxidant potential and abrogates azathioprine induced prooxidant and antioxidant imbalance in rat liver. Int. J. Pharmacol., 4: 376-381.
CrossRefDirect Link - Shyur, L.F., J.H. Tsung, J.H. Chen, C.Y. Chiu and C.P. Lo, 2005. Antioxidant properties of extracts from medicinal plants popularly used in Taiwan. Int. J. Applied Sci. Eng., 3: 195-202.
Direct Link - Tiwari, A., 2001. Imbalance in antioxidant defence and human diseases: Multiple approach of natural antioxidants therapy. Curr. Sci., 81: 1179-1187.
Direct Link - Tsakiris, S., A. Giannoulia-Karantana, I. Simintzi and K.H. Schulpis, 2005. The effect of aspartame metabolites on human erythrocyte membrane acetylcholinesterase activity. Pharmacol. Res., 53: 1-5.
CrossRefPubMedDirect Link - Van Kuijk, F.J.G.M., A. Sevanian, G.J. Handelman and E.A. Dratz, 1987. A new role of phospholipase A2: Protection of membranes from lipid peroxidation damage. Trends Biochem. Sci., 12: 31-34.
CrossRef - Veeru, P., M.P. Kishor and M. Meenakshi, 2009. Screening of medicinal plant extracts for antioxidant activity. J. Med. Plants Res., 3: 608-612.
CrossRefDirect Link - Zabri, H., C. Kodjo, B. Anoubile, J.M. Bekro and Y.A. Bekro, 2008. Phytochemical screening and determination of flavonoids in Secamone afzelii (Asclepiadaceae) extracts. Afr. J. Pure Applied Chem., 2: 80-82.
Direct Link - Aslan, M., D. Thornley-Brown and B.A. Freeman, 2000. Reactive species in sickle cell disease. Ann. N.Y. Acad. Sci., 899: 375-391.
CrossRefDirect Link